
饲粮添加亮氨酸和谷氨酸对肥育猪肌肉脂肪酸组成和脂质代谢相关基因表达的影响
2018-05-24 06:57:55胡诚军印遇龙孔祥峰江青艳
动物营养学报 2018年5期
胡诚军 张 婷 张 涛 印遇龙 孔祥峰* 江青艳
(1.中国科学院亚热带农业生态研究所,亚热带农业生态过程重点实验室,动物营养生理与代谢过程实验室,畜禽养殖污染控制与资源化技术国家工程实验室,长沙 410125;2.华南农业大学,动物科学学院,广州 510642;3.赢创德固赛(中国)投资有限公司,北京 100600)
随着我国居民生活水平的提高,消费者对猪肉品质的要求越来越高,优质猪肉更易受到消费者的青睐。脂肪是影响胴体品质和肉品风味的关键因素,腹部和皮下脂肪沉积过多会降低胴体品质,肌内脂肪含量增加则可提高肉的嫩度和风味;脂肪酸组成也是影响肉品质的关键因素,其不仅决定肉的嫩度和多汁性,还与肉的营养价值密切相关[1]。育种手段提高瘦肉率的同时又会引起肌内脂肪含量降低等问题[2],而饲粮营养调控机体脂肪代谢效果显著,这使其成为优质猪肉生产的一个重要调控手段。近年来的研究表明,亮氨酸不仅可以调节哺乳动物骨骼肌蛋白质的代谢[3],还可加速体内脂肪氧化分解,减少脂肪在体内的沉积以及脂肪细胞中甘油三酯的含量[4-5]。另外,亮氨酸还可显著增加肥育猪肌内脂肪的含量[6]。饲粮添加谷氨酸可降低肥育猪背膘厚[7],提高肌内脂肪含量[8],改善肌肉中脂肪酸的组成[9]。不过也有研究表明,饲粮添加谷氨酸可增加脂肪在体内的沉积,引起肥胖[10]。笔者前期研究表明,饲粮添加1.00%亮氨酸可提高背最长肌和股二头肌中肌内脂肪含量,且不影响肥育猪的生长性能;添加1.00%谷氨酸后,肥育猪背膘厚降低了34.3%;而添加1.00%亮氨酸+1.00%谷氨酸提高了股二头肌肌内脂肪含量,且不影响肥育猪的生长性能[11]。由上可见,亮氨酸和谷氨酸在调控动物机体脂质代谢方面均具有一定作用,且其作用靶点不尽相同,但其具体作用机制尚不明确。脂肪的沉积取决于脂肪合成和分解2个动态过程,该过程受到多个基因的调控。因此,本试验进一步探讨饲粮添加亮氨酸和谷氨酸对肌肉脂肪酸组成和脂质代谢相关基因表达的影响,旨在揭示其调控脂质代谢的分子机制,为功能性氨基酸用于优质猪肉的生产提供依据。
1 材料与方法
1.1 试验动物、分组与饲养管理
试验选取平均体重为77 kg左右的杜×长×大肥育猪60头,随机分为5组,每组12头,公母各占1/2,饲喂于HHIS-02A型全自动饲喂系统(河南河顺自动化设备有限公司产品)中。对照组饲喂基础饲粮,在基础饲粮中添加2.05%L-丙氨酸(L-丙氨酸可被机体多个组织分解代谢,因而可调节饲粮氮平衡[12])作为等氮对照组,试验组在基础饲粮中分别添加1.00%亮氨酸+1.37%L-丙氨酸(亮氨酸组)、1.00%谷氨酸+1.44%L-丙氨酸(谷氨酸组)、1.00%亮氨酸+1.00%谷氨酸(亮氨酸+谷氨酸组)。参照NRC(2012)猪营养需要推荐量配制玉米-豆粕型基础饲粮,其组成及营养水平见表1。饲养试验在位于湖南新五丰股份有限公司永安分公司的中国科学院亚热带农业生态研究所动物试验基地开展,试验期为60 d。饲养试验期间自由采食和饮水。
1.2 样品采集
饲养试验结束后,颈动脉放血处死试验猪,采集背最长肌和股二头肌,液氮速冻后-80 ℃保存,用于分子生物学分析;另外各取100 g肌肉样品于-20 ℃保存,用于脂肪酸含量测定。

表1 基础饲粮组成及营养水平(风干基础)
1)预混料为每千克饲粮提供Premix provided the following per kg of the diet:Cu 15.10 mg,Fe 150 mg,Se 0.30 mg,Zn 90 mg,Mn 61 mg,VD 386 IU,VA 9 100 IU,VE 135 IU,VK 2.24 mg,VB61.40 mg,泛酸钙 calcium pantothenate 19.70 mg,烟酸 niacin 32.20 mg,VB120.028 mg,NaCl 4.10 g,CaHPO46.50 g,CaCO310.80 g。
2)营养水平为计算值。Nutrient levels were calculated values.
1.3 脂肪酸含量测定
取肌肉冻干样品,用石油醚-乙醚(1∶1,体积比)提取脂肪后,用1 mL氢氧化钾-甲醇溶液(0.4 mol/L)酯化30 min,然后热水浴浓缩,加水分层,取500 μL上层液用气相色谱-质谱联用仪(Agilent Technologies,美国)测定,由微机按面积归一化法计算脂肪酸各组分的含量,各脂肪酸含量以占总脂肪酸的百分比表示。
1.4 脂质代谢相关基因表达测定


表2 PCR引物序列
1.5 数据处理
采用SPSS 17.0统计软件对试验数据进行Duncan氏法多重比较,结果以“平均值±标准误”表示,P<0.05表示差异显著。
2 结果与分析
2.1 饲粮添加亮氨酸和谷氨酸对肥育猪背最长肌中脂肪酸组成的影响
由表3可知,与对照组相比,亮氨酸组背最长肌中C18∶0含量显著降低(P<0.05),C18∶2n-6和C20∶1含量显著增加(P<0.05);谷氨酸组背最长肌中C14∶0和C16∶0含量显著降低(P<0.05),C17∶0、C18∶2n-6和C20∶1含量显著增加(P<0.05)。
2.2 饲粮添加亮氨酸和谷氨酸对肥育猪股二头肌中脂肪酸组成的影响
由表4可知,与对照组相比,亮氨酸组股二头肌中C16∶0含量显著降低(P<0.05),谷氨酸组股二头肌中C18∶2n-6含量显著增加(P<0.05)。
2.3 饲粮添加亮氨酸和谷氨酸对肥育猪背最长肌中脂质代谢相关基因表达的影响
由表5可知,与对照组相比,亮氨酸组背最长肌中脂肪酸转运蛋白1(FATP-1)mRNA的相对表达量显著上调(P<0.05),脂蛋白脂酶(LPL)mRNA的相对表达量显著下调(P<0.05);亮氨酸+谷氨酸组背最长肌中脂肪酸转运蛋白4(FATP-4)mRNA的相对表达量显著下调(P<0.05)。与等氮对照组相比,亮氨酸组背最长肌中FATP-1和FATP-4 mRNA的相对表达量显著上调(P<0.05),谷氨酸组和亮氨酸+谷氨酸组LPLmRNA的相对表达量显著上调(P<0.05)。
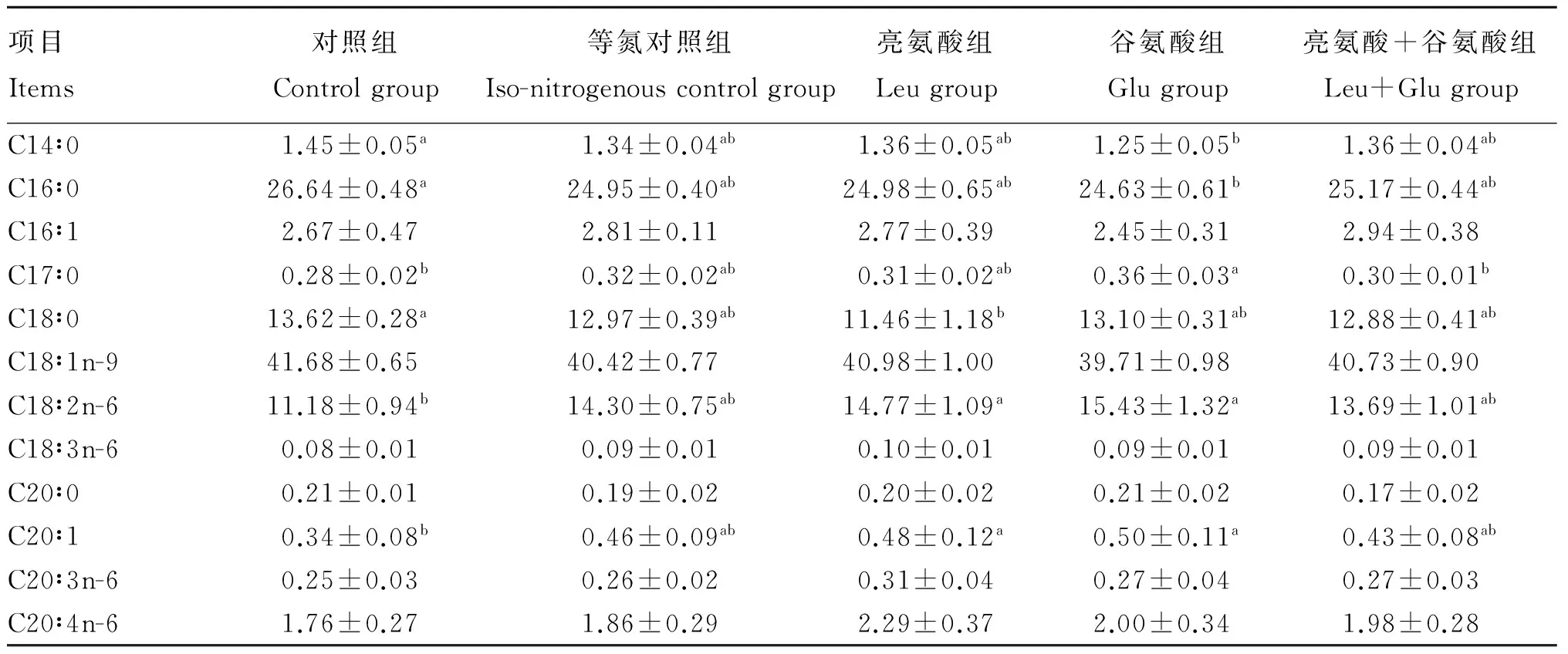
表3 饲粮添加亮氨酸和谷氨酸对肥育猪背最长肌中脂肪酸组成的影响(占总脂肪酸的百分比)

表4 饲粮添加亮氨酸和谷氨酸对肥育猪股二头肌中脂肪酸组成的影响(占总脂肪酸的百分比)Table 4 Effects of dietary Leu and Glu on fatty acid composition in biceps femoris muscle of finishing pigs (percentage of total fatty acids) %

表5 饲粮添加亮氨酸和谷氨酸对肥育猪背最长肌中脂质代谢相关基因表达的影响
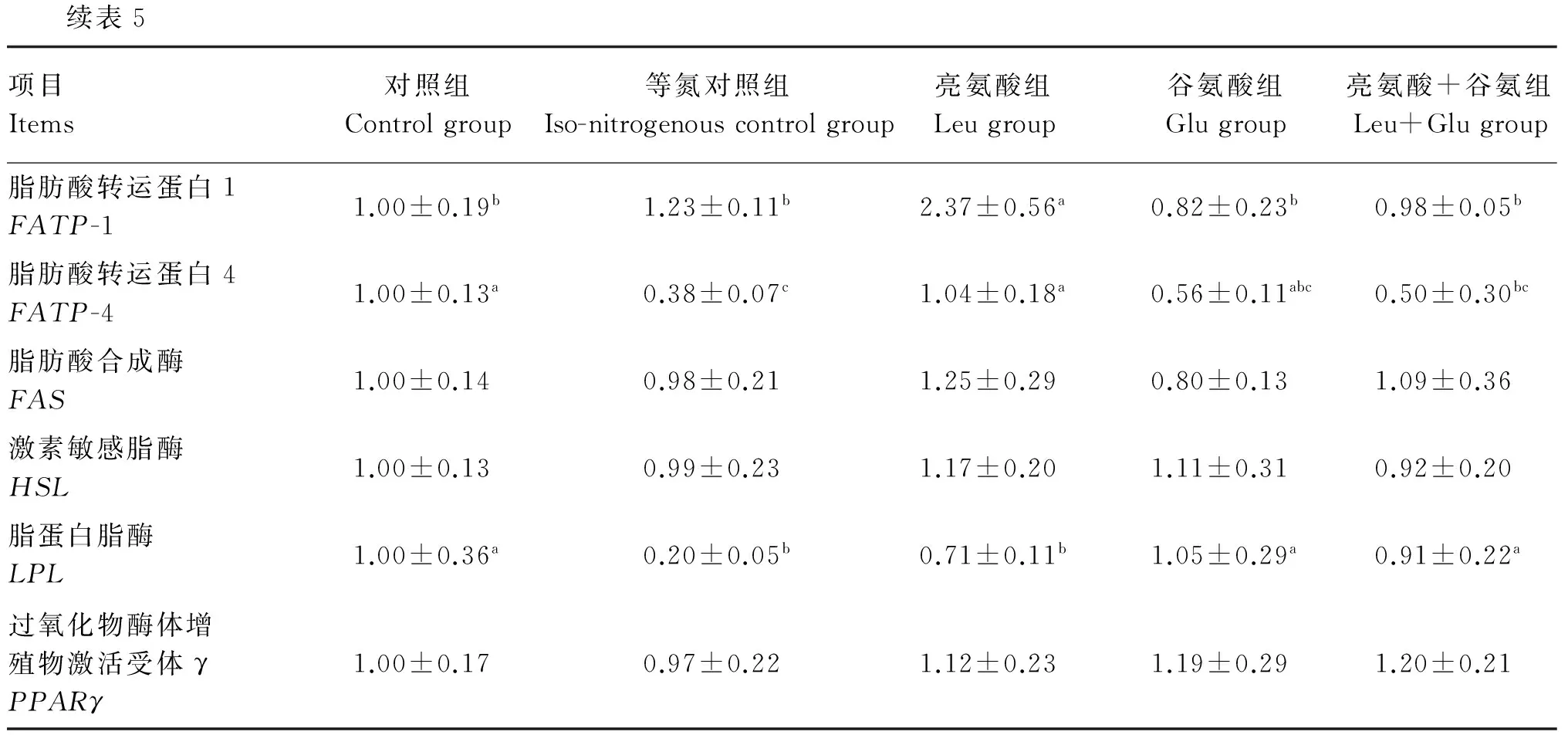
续表5项目Items对照组Controlgroup等氮对照组Iso-nitrogenouscontrolgroup亮氨酸组Leugroup谷氨酸组Glugroup亮氨酸+谷氨组Leu+Glugroup脂肪酸转运蛋白1FATP-11.00±0.19b1.23±0.11b2.37±0.56a0.82±0.23b0.98±0.05b脂肪酸转运蛋白4FATP-41.00±0.13a0.38±0.07c1.04±0.18a0.56±0.11abc0.50±0.30bc脂肪酸合成酶FAS1.00±0.140.98±0.211.25±0.290.80±0.131.09±0.36激素敏感脂酶HSL1.00±0.130.99±0.231.17±0.201.11±0.310.92±0.20脂蛋白脂酶LPL1.00±0.36a0.20±0.05b0.71±0.11b1.05±0.29a0.91±0.22a过氧化物酶体增殖物激活受体γPPARγ1.00±0.170.97±0.221.12±0.231.19±0.291.20±0.21
2.4 饲粮添加亮氨酸和谷氨酸对肥育猪股二头肌中脂质代谢相关基因表达的影响
由表6可知,与对照组相比,谷氨酸组和亮氨酸+谷氨酸组股二头肌中FATP-4 mRNA的相对表达量显著下调(P<0.05)。

表6 饲粮添加亮氨酸和谷氨酸对肥育猪股二头肌中脂质代谢相关基因表达的影响
3 讨 论
肌肉中脂肪的含量及脂肪酸的组成对猪肉品质具有重要影响,深入了解营养素调控肌肉中脂质代谢的分子机制,将有助于提高动物的胴体性状和肉品质,为消费者提供优质的肉类产品。脂肪酸组成是影猪肉风味、营养价值及氧化稳定性的一个重要因素。本试验结果发现,饲粮添加亮氨酸可显著降低肥育猪背最长肌中C18∶0[属于饱和脂肪酸(SFA)],显著增加C18∶2n-6[属于多不饱和脂肪酸(PUFA)]的含量;添加谷氨酸可显著降低肥育猪背最长肌中C14∶0和C16∶0(属于SFA),显著增加背最长肌和股二头肌中C18∶2n-6(属于PUFA)的含量;而同时添加亮氨酸和谷氨酸对肥育猪肌肉中脂肪酸组成未产生显著影响。SFA摄入过多不但会增加消费者患心血管疾病的风险[13],还易引起肌肉组织胰岛素抵抗[14],而PUFA则可预防心血管疾病[13]。C18∶2n-6,即亚麻油酸,是一种必需脂肪酸,具有抗炎和免疫调节等作用[15-16]。上述结果提示,饲粮添加亮氨酸或谷氨酸可通过降低肌肉中SFA含量来改善肌肉中脂肪酸的组成,进而提高猪肉的营养价值。然而Kong等[17]研究表明,饲粮添加谷氨酸提高了生长猪背最长肌中C16∶0的含量,上述差异可能是由于基础饲粮和肥育猪的生长阶段不同造成的。
笔者前期研究发现,饲粮添加1.00%亮氨酸和1.00%谷氨酸显著提高了肥育猪背最长肌和股二头肌中肌内脂肪的含量[11],因此本研究进一步检测了肌肉中与脂质代谢相关基因的表达情况,进一步探讨亮氨酸和谷氨酸影响肌内脂肪代谢的机制。本研究发现,饲粮添加亮氨酸可显著上调肥育猪背最长肌中FATP-1和FATP-4基因的表达。FATP-1和FATP-4是脂肪酸转运蛋白家族的2个成员,在脂肪酸转运过程中发挥着重要作用。敲除FATP-1基因可减少肌肉细胞对长链脂肪酸(LCFA)的摄取、降低肌肉中甘油三酯的沉积[18],过表达FATP-1基因可提高肌肉细胞对葡萄糖的氧化[19];而FATP-4基因的表达量则与肥胖有关[20]。上述结果提示,饲粮添加亮氨酸可能通过促进肌肉细胞对LCFA的摄取,进而促进脂肪在肌肉中的沉积。笔者前期研究发现饲粮同时添加亮氨酸和谷氨酸组合可显著提高背最长肌肌内脂肪含量[11],然而本试验中饲粮同时添加亮氨酸和谷氨酸则显著下调了背最长肌中FATP-1和FATP-4基因表达,其具体机制还有待进一步研究。
乙酰辅酶A羧化酶(ACC)和脂肪酸合成酶(FAS)是合成脂肪酸的2个关键酶[21],FAS基因表达量上升可显著促进甘油三酯的沉积,进而引起肥胖[22]。过氧化物酶体增殖物激活受体γ(PPARγ)是调节脂肪分化的核心调控因子,具有脂肪组织特异性,除了直接参与脂肪分化外,还可调节FAS、ACC、LPL和激素敏感脂酶(HSL)等基因的表达[23],间接影响脂质代谢。可见,ACC、FAS和PPARγ基因均与脂肪合成代谢有关。在本试验中,饲粮添加亮氨酸、谷氨酸和亮氨酸+谷氨酸均未对育肥猪肌肉中ACC、FAS和PPARγ的表达产生显著影响,提示亮氨酸和谷氨酸增加肌内脂肪含量可能不是通过上调脂肪合成基因表达实现的。本研究结果与Madeira等[24]的结果一致,即饲粮添加亮氨酸不显著影响背最长肌中ACC和FAS基因的表达;而Kong等[17]研究发现,饲粮添加谷氨酸可提高背最长肌中肌内脂肪含量,上调ACC基因的表达。结果不一致的原因可能是因为基础饲粮不同,Kong等[17]是在高脂饲粮中添加谷氨酸。
FAS可催化乙酰辅酶A生成C16∶0[25]。在本研究中,饲粮添加谷氨酸显著降低了肥育猪背最长肌中C16∶0的含量,饲粮添加亮氨酸显著降低了股二头肌中C16∶0的含量,但未对肌肉中FASmRNA的相对表达量产生显著影响,提示亮氨酸和谷氨酸可能不影响C16∶0的合成代谢,但可上调与C16∶0分解相关基因的表达。LPL的主要功能是催化水解乳糜微粒和极低密度脂蛋白中的甘油三酯,动物体内LPL活性是决定外周组织从血浆中摄取甘油三酯的重要因素,其表达量增加能够显著增加脂肪在皮下和肌肉中的沉积[25-26]。本试验结果表明,饲粮添加谷氨酸和亮氨酸+谷氨酸可显著上调肥育猪背最长肌中LPL基因的表达,提示其可通过调控LPL基因的表达影响肌肉中脂肪的沉积。
4 结 论
饲粮添加亮氨酸可显著降低肥育猪背最长肌中C18∶0含量,显著增加C18∶2n-6含量;饲粮添加谷氨酸可显著降低肥育猪背最长肌中C14∶0和C16∶0含量,显著增加C18∶2n-6含量;饲粮同时添加亮氨酸和谷氨酸对肥育猪肌肉中脂肪酸的组成未产生显著影响,但可显著上调背最长肌中LPL基因的表达。
参考文献:
[1] WOOD J D,RICHARDSONI R I,NUTE G R,et al.Effects of fatty acids on meat quality:a review[J].Meat Science,2004,66(1):21-32.
[2] ROS-FREIXEDES R,REIXACH J,BOSCH L,et al.Response to selection for decreased backfat thickness at restrained intramuscular fat content in Duroc pigs[J].Journal of Animal Science,2013,91(8):3514-3521.
[3] JEWELL J L,KIM Y C,RUSSELL R C,et al.Differential regulation of mTORC1 by leucine and glutamine[J].Science,2015,347(6218):194-198.
[4] ZHANG Y Y,GUO K Y,ROBERT E L,et al.Increasing dietary leucine intake reduces diet-induced obesity and improves glucose and cholesterol metabolism in mice via multi mechanisms[J].Diabetes,2007,56(6):1647-1654.
[5] FREUDENBERG A,PETZKE K J,KLAUS S.DietaryL-leucine andL-alanine supplementation have similar acute effects in the prevention of high-fat diet-induce obesity[J].Amino Acids,2013,44(2):519-528.
[6] HYUN Y,ELLIS M,MCKEITH F K,et al.Effect of dietary leucine and lysine levels on intramuscular fat content in finishing pigs[J].Canadian Journal of Animal Science,2003,83(2):315-318.
[7] HU C J,JIANG Q Y,ZHANG T,et al.Dietary supplementation with arginine and glutamic acid modifies growth performance,carcass traits,and meat quality in growing-finishing pigs[J].Journal of Animal Science,2017,95(6):2680-2689.
[8] 周笑犁,孔祥峰,范觉鑫,等.味精与高脂日粮对生长猪胴体性状与组成的影响[J].食品工业科技,2014,35(5):330-333,337.
[9] HU C J,JIANG Q Y,ZHANG T,et al.Dietary supplementation with arginine and glutamic acid enhances key lipogenic gene expression in growing pigs[J].Journal of Animal Science,2017,95(12):5507-5515.
[10] NARDELLI T R,RIBEIRO R A,BALBO S L,et al.Taurine prevents fat deposition and ameliorates plasma lipid profile in monosodium glutamate-obese rats[J].Amino Acids,2011,41(4):901-908.
[11] 胡诚军,张婷,李华伟,等.饲粮添加亮氨酸和谷氨酸对肥育猪生长性能、胴体性状和肉品质的影响[J].动物营养学报,2017,29(2):590-596.
[12] TAN B,YIN Y L,LIU Z Q,et al.DietaryL-arginine supplementation increases muscle gain and reduces body fat mass in growing-finishing pigs[J].Amino Acids,2009,37(1):169-175.
[13] KIM M J,PARVIN R,MUSHTAQ M M H,et al.Influence of monochromatic light on quality traits,nutritional,fatty acid,and amino acid profiles of broiler chicken meat[J].Poultry Science,2013,92(11):2844-2852.
[14] KENNEDY A,MARTINEZ K,CHUANG C C,et al.Saturated fatty acid-mediated inflammation and insulin resistance in adipose tissue:mechanisms of action and implications[J].The Journal of Nutrition,2009,139(1):1-4.
[15] PISCHON T,HANKINSO S E,HOTAMISLIGIL G S,et al.Habitual dietary intake of n-3 and n-6 fatty acids in relation to inflammatory markers among US men and women[J].Circulation,2003,108(2):155-160.
[16] KALOGEROPOULOS N,PANAGIOTAKOS D B,PITSAVOS C,et al.Unsaturated fatty acids are inversely associated and n-6/n-3 ratios are positively related to inflammation and coagulation markers in plasma of apparently healthy adults[J].Clinica Chimica Acta,2010,411(7/8):584-591.
[17] KONG X F,ZHOU X L,FENG Z M,et al.Dietary supplementation with monosodiumL-glutamate modifies lipid composition and gene expression related to lipid metabolism in growing pigs fed a normal-or high-fat diet[J].Livestock Science,2015,180:247-252
[18] RICHARDS M R,LISTENBERGER L L,KELLY A A,et al.Oligomerization of the murine fatty acid transport protein 1[J].Journal of Biological Chemistry,2003,278(12):10477-10483.
[19] GUITART M,ANDREU A L,GARCA-ARUMI E,et al.FATP1 localizes to mitochondria and enhances pyruvate dehydrogenase activity in skeletal myotubes[J].Mitochondrion,2009,9(4):266-272.
[20] GERTOW K,PIETILINEN K H,YKI-JRVINEN H,et al.Expression of fatty-acid-handling proteins in human adipose tissue in relation to obesity and insulin resistance[J].Diabetologia,2004,47(6):1118-1125.
[21] LIU Y Y,LI F N,HE L Y,et al.Dietary protein intake affects expression of genes for lipid metabolism in porcine skeletal muscle in a genotype-dependent manner[J].British Journal of Nutrition,2015,113(7):1069-1077.
[22] 尹靖东.动物肌肉生物学与肉品科学[M].北京:中国农业大学出版社,2011.
[23] LAPLANTE M,SELL H,MACNAUL K L,et al.PPAR-γ activation mediates adipose depot-specific effects on gene expression and lipoprotein lipase activity-mechanisms for modulation of postprandial lipemia and differential adipose accretion[J].Diabetes,2003,52(2):291-299.
[24] MADEIRA M S,PIRES V M,ALFAIA C M,et al.Combined effects of dietary arginine,leucine and protein levels on fatty acid composition and gene expression in the muscle and subcutaneous adipose tissue of crossbred pigs[J].British Journal of Nutrition,2014,11(9):1521-1535.
[25] 祝仁铸,尹逊河,王元虎,等.猪肌肉组织MDH和LPL基因表达与肌内脂肪含量和脂肪酸组成关系的研究[J].畜牧兽医学报,2013,44(8):1182-1188.
[26] VINCENT G,ISABELLE L,JACQUES M,et al.Lipogenic enzyme activities in subcutaneous adipose tissue and skeletal muscle from neonatal pigs consuming maternal or formula milk[J].Reproduction,Nutrition,Development,2000,40(2):103-112.
猜你喜欢
快乐作文(1.2年级)(2023年4期)2023-05-08 13:27:42
临床骨科杂志(2021年6期)2022-01-08 09:11:50
解剖学杂志(2021年6期)2021-12-31 03:25:40
兽医导刊(2021年19期)2021-11-12 03:07:40
猪业科学(2018年5期)2018-07-17 05:56:14
中华骨与关节外科杂志(2017年1期)2017-05-17 06:11:23
天然产物研究与开发(2016年11期)2016-06-15 20:29:17
动物营养学报(2015年10期)2015-12-01 02:26:21
大连工业大学学报(2014年2期)2014-09-19 08:52:52
食品科学(2013年19期)2013-03-11 18:27:35
