
黄土高原植物标本采集完整性评估 及其环境解释
2018-05-20 06:56:58田慧敏简琴范雪
数据与计算发展前沿 2018年5期
田慧敏,简琴,范雪
1.安徽农业大学资源与环境学院,安徽 合肥 230036
2.中国科学院植物研究所,北京 100093
引言
黄土高原地区从南至北纵跨暖温带,中温带两个热量带,从东到西横跨半湿润、半干旱两个干湿区,地理成分复杂[1];黄土高原地理位置相对特殊,处于从平原过渡到山地高原、从沿海过渡到内陆、从农业过渡到牧业的地区,多种成分相互交错,是中国气候变化的敏感地带[2];黄土高原在中国植物区系上属于泛北极植物区,计有种子植物 147 科 864 属 3224 种,具有中国特有属 32 个,黄土高原地区特有属 4个,特有种 1 64 个[3]。黄土高原地区植物种类比较丰富,在全国植物区系上占有重要的地位;植物区系起源古老,既有原始类群和孤立类群,又有进化类群;一些大科大属的种类非常丰富,代表性的如禾本科、菊科、豆科、蔷薇科等在整个区系中都具有重要作用。小型科的一些种分布也很广泛,如臭椿、香椿、柿树等都对黄土高原地区的生态恢复和经济建设具有重要的意义。黄土高原地区分布的国家级珍稀濒危保护植物有 84 种,包括国家一级重点保护植物水杉、国家二级重点保护植物银杏等,且包含一些规模小、密度低的生理生态类型,但黄土高原地区自然环境条件不够稳定,生态环境脆弱,水土流失等自然灾害频繁,可能导致这些类群灭绝,因此黄土高原的研究对生物地理学以及保护生物学都有极其重要的意义。
黄土高原地区由于其特殊的地理位置和丰富的生物多样性,得到了国内外学者的诸多研究,其中物种丰富度的大尺度地理格局是研究热点问题。冯建孟利用大尺度上的物种分布数据和地理信息,讨论物种空间分布与地理因子之间的关系[4];信忠保等利用归一化植被指数,研究了黄土高原植被覆盖的时空变化,并解释了气候和人为因素方面的原因[5];程杰研究了黄土高原地区不同草地植被类型退化与恢复的水分、养分时空变化过程,揭示不同植被地带指示种群与气候变化的响应趋势及空间变异特征,并提出植被恢复的调控措施[6];王世雄研究了黄土高原群落演替中物种多样性的变化规律,用CCA分析了物种多样性和环境的联系,并采用加性分配方法,提出物种多样性与空间尺度之间的关系[7-8];李强使用 GIS 分析了黄土高原南部和典型区土地利用的时空差异特征,并参照CA-Markov 模型和土地适宜性选择耕地的优化配置方案[9]。
上述研究都是利用黄土高原地区已有的物种分布信息进行研究,但目前的物种分布数据存在着不足,黄土高原地区标本采集强度存在明显的空间和类群上的偏差,以及标本的地理坐标信息不精确,导致县级物种名录的不完整,这对大尺度物种丰富度格局的分析产生了不利的影响。因此数据质量的评估对数据的正确使用和研究结果的准确性有重要作用。但目前对黄土高原标本采集数据质量的研究较少,本研究将从标本采集偏差入手,对黄土高原采集完整性进行评估,为黄土高原地区的植物保护提供参考意见。
本研究拟以国家标本资源共享平台收录的黄土高原植物的标本采集记录为研究对象,地理单元选取县级行政单位,进行如下分析:(1) 黄土高原植物标本记录质量评估,包括:标本记录数量,物种丰富度,预估物种丰富度,物种采集完整性评估。(2) 利用与采集地点相对应的环境因子对物种丰富度格局和标本采集完整性进行解释。探讨温度、降水量对物种丰富度格局影响的相对重要性,解释其多样性维持的重要影响因素。相关结果对指导黄土高原标本的采集、生物多样性空间格局研究以及濒危植物保护策略制定等领域有着重要的借鉴意义。
国家标本资源共享平台 (简称NSII,http://www.nsii.org.cn/) 是国家科技部认定并资助的国家科技基础条件平台之一,包括植物、动物、教学、自然保护区、岩矿化石、极地等六个标本子平台。其汇集了植物、动物、岩矿化石和极地资源等标本记录、标本照片、名录、文献等资源。标本数字化是 NSII 的核心和重点,经过十几年的建设和积累,NSII 成为拥有将近 1400 万份数字化标本的大数据平台,其中植物标本已超过 1013 万份。植物标本数字化的同时,标本信息包括分类信息 (科属种的学名、标本状态:花果)、采集信息 (采集人、采集时间、采集地、采集号、经纬度坐标)、馆藏信息 (条形码、存放的标本馆) 等整合在数据库中。特别是学名和分布地字段的规范和完善,为分类学和生态学的研究提供了坚实的数据基础。经过地标化的标本数据,为解决生态学的一些关键问题具有很好的助推作用。
1 材料与方法
1.1 数字化标本记录的获取与筛选
本研究所用的数字化标本记录全部来源于国家标本资源共享平台 (National Specimen Information Infrastructure,NSII)。经过初步筛选,共获取到黄土高原植物标本采集记录 84 万份。由于植物标本采集信息经过很长的时间,年代久远,部分数据的标签信息存在缺失或字迹模糊等情况,在数字化过程中会产生错误信息。为了进一步提高数据质量,本文首先对这些数据进行筛选和校对,步骤如下:
(1) 剔除采集地点缺失以及在中国行政区以外的采集记录。(2) 剔除重复的标本采集记录。(3) 校对植物拉丁名。利用 (http://tnrs.iplantcollaborative.org/TNRSapp.html) 网站将标本拉丁名进行剔除错误数据与标准化,将查询标本拉丁名准确率得分在0.9以下的标本数据剔除,准确率得分在0.9以上的确定为正确拉丁名。(4) 对于采集地点在自然保护区内的标本记录,根据百度地图查询自然保护区所在地,若完整的自然保护区在某个县级地区内,将保护区名改为该县名。(5) 剔除采集地点中无详细地名,属于路边或街道的采集记录。(6) 县级行政单位名称校对。根据中国县级行政区划地图确定最新县名。经过以上整理后共获取了约 187140 条地标化至县级的黄土高原植物标本数据,覆盖了黄土高原内 166 个县级行政单位。
1.2 环境因子的获取
根据 (E.M.O'Brien) 提出的水分-能量动态假说,物种多样性的大尺度格局是由水分和能量共同决定的[10]。水分和能量是物种丰富度的重要影响因素,但在不同区域有着相对的重要性[11]。依据这两大类因子,本文从世界气候数据库 (http://www.worldclim.org/) 中获取了具体的 19 个生物气候因子,包括 11 个能量因子 (年平均气温,平均温度日较差,等温性,温度季节性,最热月的最高温度,最冷月的最低温度,气温年较差,最湿季节的平均气温,最干季节的平均气温,最暖季节的平均气温,最冷季节的平均气温) 和 8 个水分因子 (年降水量,最湿月的降水量,最干月的降水量,季节性降水量,最湿季度降水量,最干季度降水量,最暖季度降水量,最冷季度降水量)。为了避免因子之间出现多重共线性,本文采用 SPSS 中的 spearman 相关分析法来计算各因子之间的相关系数,将所有因子两两相互作用添加进模型,若两者相关性大于 0.75,表示相关性大,则只保留一个因子。再利用 SAM 进行模型筛选,当 Delta AICc<3 时,说明模型重要,可以得出符合要求的几种重要模型,模型中环境因子出现的次数越多,说明重要性越大。
1.3 物种丰富度与 CHAO1 估计
通常情况下,采集记录的物种丰富度小于实际物种丰富度。Chao (1984) 提出了一种基于稀有种数量的群落丰富度估计方法来估计每个县真实的物种数,通常被称为 Chao1 估计量。Chao1 的估计以已有取样中物种个体数量的分布频率为基础[12],当取样中每个物种都是两个或两个以上的个体时,Chao1 认为该地区的取样比较完全;而当取样中每个物种只有一个个体时,Chao1 认为该地区仍有新物种有待发现。其计算方程为:
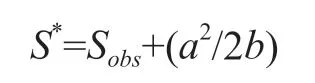
其中,Sobs是采集到的物种数,a 是取样中只有一个个体的物种数,b 是取样中只有两个个体的物种数, S*是 Chao1 估计的物种数。根据该公式可得知每个县中稀有种数量越多,该地区新物种未被发现的概率越大,估计丰富度越高;而每个县中稀有种数量越少,该地区新物种未被发现的概率越小,估计丰富度越小。在计算过程中本研究将稀有种个数定为 10。
1.4 标本采集记录完整性评估
由于某些偏远地区难以采集以及采集人的主观因素导致有些热点地区采集强度很高,另一些偏远地区采集明显不足,标本采集数量在地理上分布极不均匀。采集地理偏差会导致植物分布信息不能完整记录以及某些稀有种未被发现,因此对每个县的采集记录进行完整性评估对于生物地理学和保护生物学的研究十分重要。
本文采用两种方法来评估黄土高原每个县的植物物种在该地的采集数据的不完整程度。第一种方法用估计丰富度和采集丰富度的差值来表示采集的不完整程度,差值越大,表明不完整程度越高。第二种方法是利用物种累积曲线 (species accumulation curve, SAC) 的弯曲程度来反映不完整程度。物种积累曲线的弯曲程度越小即越接近于直线,说明采样越差;弯曲程度越大,采样越好。物种积累曲线用R程序包“vegan”中的“specaccum”函数构建,方法是“exact”。该方法计算的是当对县的样本进行重抽样时,抽取 n 个个体时,这 n 个个体包含的平均 (或期望) 物种数量。物种积累曲线的原理是对采集记录随机抽样,随着采集强度的增加,新物种增加的概率也会增加。我们选取物种积累曲线末端 10% 的斜率表示物种积累曲线的弯曲程度来代表每个县的采集名录不完整程度,斜率接近 0,采集越完整;斜率接近 1,采集越不完整。由于统计学上 95% 的置信区间,因此可以设定一个阈值 0.05,当末端斜率小于或者等于 0.05 (SAC slope≤0.05) 时,可以认为县的物种名录较为完整,采集继续时,新物种增加的概率较小;当曲线末端斜率大于 0.05 (SAC slope>0.05) 时,认为该县的物种名录不完整,采集继续时,新物种增加的概率较大。
1.5 植物标本采集完整性的环境因子解释
本文在 SPSS 中进行回归分析,以标本采集数量、原始丰富度、丰富度估计值和丰富度残差四个参数分别作为因变量,环境因子作为自变量,采用逐步回归的方法对其进行拟合。本文在回归分析结果中选取了三个参数,分别是调整后的 R2 值、标准系数 β 和 t 值 (表 1)。
2 结果
2.1 黄土高原地区标本采集数量及丰富度情况
从采集数量来看,黄土高原地区植物标本的采集集中在山西省,青海省和陕西省。其中陕西省眉县和山西省交城县的采集数量最为丰富,植物标本数量分别为 18924 份和 10341 份。采集强度主要在陕西省和山西省的西北部较低,有标本采集记录的县级行政单位呈零星分布 (图 1,A)。
黄土高原植物物种丰富度记录数和 Chao1 估计量最高的地区为眉县、长安县和周至县,物种丰富度较高的地点主要是陕西省西南部,与陕西省植物物种丰富度格局相似。
2.2 黄土高原标本采集完整性评估
Chao1 估计物种丰富度和记录物种丰富度残差最高的地区为周至县,眉县和户县 (图 1,D),残差值>100 种,代表区域内的稀有种数量较多。其中有 84 个县的残差值都大于 100,说明黄土高原大部分地区的稀有种数量都很多。物种积累曲线结果显示尾端 10% 斜率均大于 0.05,表明有标本采集记录的全部县级行政单位的标本采集都是不完整的。
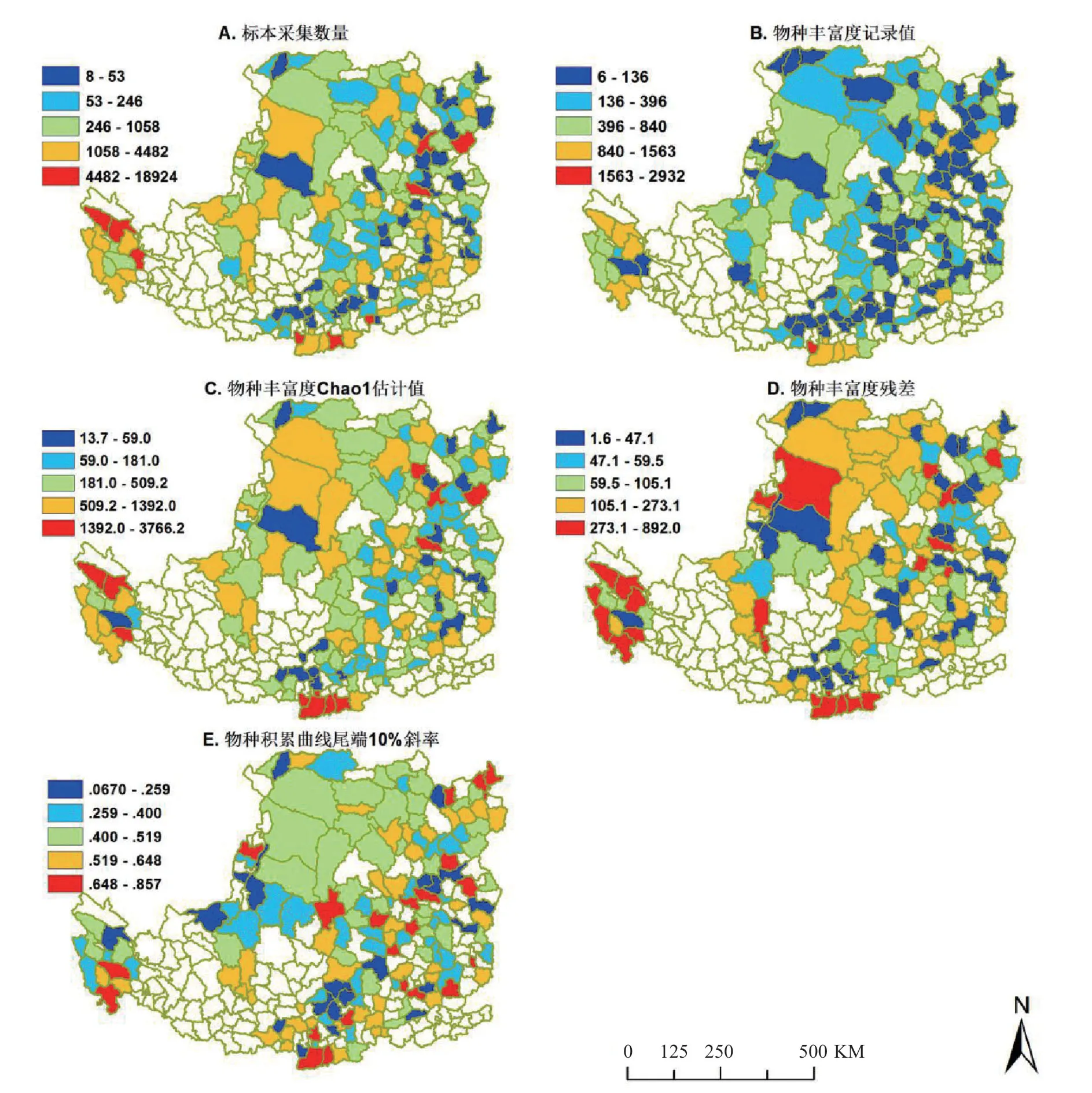
图1 A.标本采集数量;B.原始丰富度(物种数)C.丰富度估计值;D.残差(丰富度估计值-实际值);E.物种积累曲线尾端10%斜率。Fig.1 A: Number of collected specimens; B: Original richness; C: Richness estimates; D: Richness residue (estimated value minus actual value); E: Gradient of the last 10% section of the accumulated species curve
2.3 环境因子对黄土高原丰富度格局及采集的影响
经过 SPSS 和 SAM 中的模型筛选后,共保留了 1 个能量因子 (AMT年平均温度) 和 3 个水分因子 (AP年降水量、WMP 最湿月降水量、WQR 最暖季度降水量)。
环境因子 (能量和水分因子) 对标本采集数量、原始丰富度、丰富度估计值、丰富度残差均有较强的解释能力 (表 1)。
其中环境因子对丰富度估计值的解释能力最强。对标本采集数量、原始丰富度、丰富度估计值、丰富度残差进行逐步回归,根据最优线性模型分析结果中的 R2-adj,WQR 最暖季度降水量在所有回归模型中的解释率均是最强的。通过回归方程的标准化系数可知,年降水量和最湿月降水量与标本采集数量、原始丰富度、丰富度估计值、丰富度残差之间均呈显著关系,且均为正相关;年平均温度和最暖季度降水量与标本采集数量、原始丰富度、丰富度估计值、丰富度残差之间均呈显著关系,且均为负相关。各丰富度之间与环境因子均有一定的相关性。
3 讨论
3.1 黄土高原地区植物分布格局分析
黄土高原植物种类丰富,山西省、陕西省都存在着丰富的黄土高原植物记载,但是由于其气候和地形的多变导致采集地点呈零星分布 (图 1,A)。黄土高原植物标本从采集数量来看多集中在我国山西省、青海省和陕西省。山西省的种子植物区系以温带分布区为主,具有典型的温带性质[13]。陕西省几乎包含了我国北温带分布的全部典型木本属,陕西省植物区系不仅是表面上的温带性质,还很可能是温带植物区系分化和发育的重要场所[14]。因此黄土高原地区种子植物主要分布于温带地区,主要是北温带地区。从采集地点来看,陕西省和山西省的东南部植物标本采集记录较多,西北部采集记录较少,原因可能是黄土高原地区从东南到西北温度逐渐降低,降水量逐渐减少,水热条件变差,物种丰富度较低。另外在山西省中部的清徐县与交城县地理位置很相近,但标本采集记录差距非常明显,可能是由于采集学家的个人主观意识差异。
黄土高原植物丰富度和丰富度估计值最高的区域出现在陕西省的眉县、长安县和周至县,均位于陕西省的西南部。与陕西省植物丰富度分布格局相似。吴征镒提出黄土高原地区中我国特有属分布以西南为中心,呈扇形向东南,东北,东部辐射[15]。陕北气候干旱降雨少,黄土覆盖很厚,且切割严重;陕南温度较高且湿润,山地多,森林茂密,植物数量较多,稀有种数量多,导致南北植物丰富度差异大。因此黄土高原地区物种丰富度与纬度有一定的相关性,物种丰富度随纬度的升高而逐渐降低。
3.2 黄土高原植物标本采集完整性评估
根据物种积累曲线尾端 10% 斜率最大得出的采集最不完整的县是交口县和寿阳县,与物种丰富度残差值最高的县级行政单位有差别,分析其可能的原因是:交口县的采集物种数量极少,利用 Chao1 计算出的物种丰富度与记录丰富度的残差值也较小,但是按照物种积累曲线的斜率来看稀有种数量较多,因此两种完整性评估方式对于采集强度极度分化的地区而言可能有所差异。但总体而言,物种丰富度残差值越大的县,其物种积累曲线尾端 10% 的斜率值也越大,表明 Chao1 估计量和物种积累曲线都能够对标本采集完整性进行评估,两种方法的评估结果具有相关性。分析结果表明黄土高原所有的县级行政单位均为采集不完整,很多地区的大量稀有种还有待发现,大部分学者针对黄土高原地区植物物种多样性的调查可能并不十分准确。因此对黄土高原进行全面的调查采集非常有必要,未来野外调查的任务仍然十分艰巨。
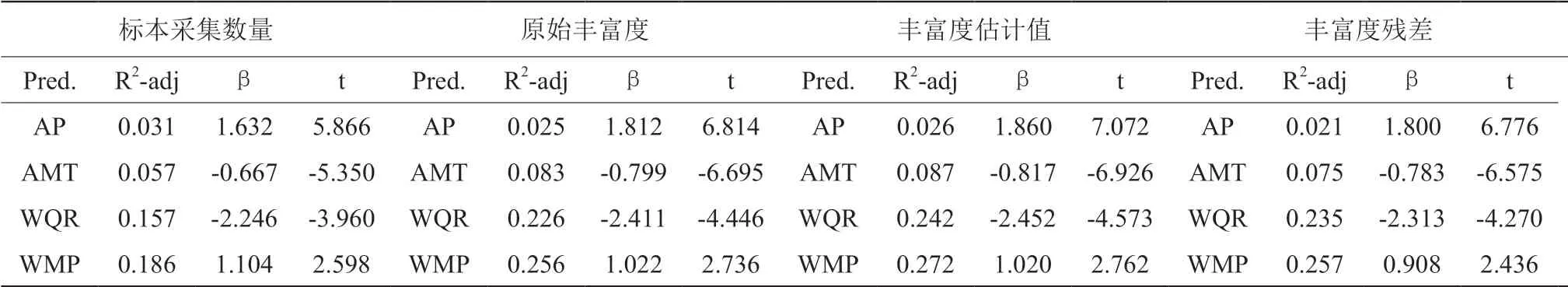
表1 环境因子与标本采集数量、原始丰富度、丰富度估计值、丰富度残差之间逐步回归结果Table 1 Results of gradual regression among environmental factors, collected specimen quantity, original richness, richness estimates, richness residue of the species accumulated curve
3.3 物种丰富度及采集完整性与环境因子关系
年降水量和最湿月降水量与黄土高原植物原始丰富度,估计丰富度以及丰富度差值有显著的正相关关系,即年降水量的增加更有利于植物的生长,黄土高原物种丰富度高的区域出现在降水量丰富的地区,且降水量的增加有利于标本采集完整。黄土高原处于干旱的内陆地区,属于温带大陆性季风气候,降水量季节差异明显,全年总雨量少,降雨主要集中在 6-9月[16]。对于干旱半干旱的黄土高原地区,降水量是其丰富度的主要限制因子。
年平均温度和最暖季度降水量与黄土高原植物丰富度有显著的负相关关系,表明温度和最暖季度降水量的增加不利于标本采集的完整。黄土高原日照充足,年平均温度≥10°,气温除了受到季风气候的影响,纬度和下垫面也会影响温度变化,温度与植物丰富度呈负相关主要是由于自然环境变化引起作物熟制不同导致。最暖季度降水量对黄土高原植物丰富度有最强的影响力,原因是黄土高原降雨集中在暖季,且降水强度大,一次暴雨的量可达到全年降水的 30%,因此最暖季度降水量对黄土高原地区植物丰富度有着显著的影响,其增多尤其不利于植物丰富度的增加,也不利于标本采集完整。
最优线性模型对各物种丰富度的总体解释率偏低,物种丰富度的地理变异大部分未被解释,说明除了水分和能量因子,仍然有一些影响物种丰富度格局的因素未被提及,如局域尺度的环境要素和区域历史对物种丰富度格局形成和演化的影响[17]。
致谢
感谢国家标本资源平台提供的 84 万份黄土高原地区植物标本记录数据,以及感谢张明罡老师,肖翠老师的指导。
猜你喜欢
《学习方法报》地理商务星球七年级(2023年13期)2023-10-11 02:44:46
启蒙(3-7岁)(2019年8期)2019-09-10 03:09:08
中国房地产业·上旬(2019年12期)2019-03-24 09:44:41
烟台果树(2019年1期)2019-01-28 09:34:46
江西农业(2018年23期)2018-02-11 07:26:59
现代园艺(2018年3期)2018-02-10 05:18:21
公民与法治(2016年3期)2016-05-17 04:09:00
公民与法治(2016年1期)2016-05-17 04:07:56
水利科技与经济(2016年6期)2016-04-22 05:08:06
中国火炬(2015年7期)2015-07-31 17:39:57
