
The nutritional composition and digestion of plants foraged by red deer(Cervus elaphus xanthopygus)in northeast China
2018-05-19 03:36:36YuanFengYanzeYuLinqiangZhongWeiqiZhangMinghaiZhang
Journal of Forestry Research 2018年3期
Yuan Feng•Yanze Yu•Linqiang Zhong•Weiqi Zhang•Minghai Zhang
Introduction
For free-living ungulates,the quality of their food is a key factor impacting their behavior and performance.Suttie and Hamilton(1983)revealed that the plane of nutrition is the single most important factor determining body weight in red deer.Body weight of deer is an indictor of body growth and fecundity(Albou et al.1986;Galliard et al.1992).Body weight is one constraint on entry into puberty for young female red deer(Hamilton and Blaxter 1980;Asher and Cox 2013).Pre-mating nutritional status of female red deer may affect the sex ratio of the offspring(Luna-Estrada et al.2006).
During the long,harsh Arctic winter,caribou eat lichens that are low in crude protein(CP)in advance of the early spring birthing and lactation seasons,and this results in a negative protein balance that lasts for 7 months(Gerhart et al.1996).Lichens contain relatively high content of digestible energy and the ratio of digestible energy to CP in lichens is higher than that of other species forged by caribou during winter(Parker et al.2005).Caribou show strong seasonal variations in nutritional requirements and acquisition.High demand for digestible energy during winter and for lactation in summer are the most important nutritional factors limiting Alaska black tailed deer(Parker et al.1999).Similarly,white-tailed deer(Odocoileus virginianus)in Quebec,Canada,selected winter forage species that contained the highest content of digestible energy,re flecting the physiological demands of winter survival(Berteaux et al.1998).Energy intake of red deer hinds during lactation is a major determinant of calf growth performance(Asher et al.2011).The annual energy balance is usually more sensitive to changes in energy intake than energy expenditure;therefore,wild animals may not limit their maximum energy intake(Parker et al.2009).
Another important factor for evaluating the nutritional value of plants is Dry Matter Digestibility (DMD).McDonald etal.(1995)showed thatplants with DMD>50%foraged by ruminants were considered to be high-quality food.DMD can be determined both by experiments and by theoretical calculations.Ouyang et al.(2000)compared the mask method with the ash insoluble acid(AIA)method for the determination of daily intake and drymatter digestion by Tibetan goats and reported similar results from the two methods.Lei(1994)used statistical analysis to investigate the relationship between DMD of alfalfa(Sativa medicago)and its chemical composition.They concluded that quantifying crude fiber content was the most reliable method for estimating the DMD of alfalfa.
The nutritional quality of plants,especially at northern latitudes,is generally much lower in winter than in other seasons(Gonzalez-Hernandez and Silva-Pando 1999;Lo´pez-Pe´rez et al.2012;Parlak et al.2011;Tixier et al.1997).The aim of this study was to evaluate nutritional components of winter diets of red deer(Cervus elaphus xanthopygus)occupying the Muling forest region of northeastern China.We investigated the energy content and macronutrients(crude protein,crude lipid,non- fiber carbohydrate(NFC),andneutraldetergent fiber(NDF))of15plant species foraged by red deer in January of two consecutive years.We quanti fied relationships between energy content and macronutrients and fiber content of the 15 plant species.
Materials and methods
The study area was the Muling Dongbei Hongdoushan Nature Reserve(MDHNR).This reserve is located in southeastern Heilongjiang Province.It has a typical temperate continental monsoon climate,four seasons,and an annual average temperature of-2°C.Winter in the reserve is cold and long,with an extremely low temperature in January(minimum-43.8°C).The snowfall period is from October to April with an average depth of 500 mm.Red deer forage plant samples were collected in January 2014 and 2015.Fresh trails of red deer in the reserve were followed and feces and plants along the trails were collected for laboratory analysis.CP content was determined by measuring total nitrogen concentration according to the Kjeldahl method.We used Soxhlet equipment with an ether-ethyl solvent to quantify crude lipid(CL)content.NDF,ADF(acid detergent fiber),and ADL(acid detergent lignin)were quanti fied using the Van Soest technique(van Soest and Lewis 1991).The relation between the content of various cellulose-base substances was calculated by the relative amount of NDF compounds,including the proportions of cellulose,hemicellulose and lignin in NDF.DMD was estimated by theoretical calculations based on ADF and nitrogen content as outlined by Oddy et al.(1978).The formula is given as follows:DMD=83.58-0.824ADF+2.626N%
To investigate the relationship between macronutrients and energy content,we used the Pearson product-moment correlation coefficient(PPMCC or PCC)calculated as follows:

PPMCC is a measure of the linear correlation between two variablesxandy.It has a value between+1 and-1,where 1 is total positive linear correlation,0 is no linear correlation,and-1 is total negative linear correlation.r is the coefficient of association and n is sample size(Gao et al.2011).
For representing the plant species foraged by red deer,we listed the abbreviations and icons which used in the figures and tables of this study(Table 1).
Results
Nutritional composition of red deer winter diets
The nutritional composition of 15 species foraged by red deer varied widely(Table 2).Energy content was statistically similar for all foraged plant species and averaged 18.4±0.7 kJ/g across all species.Among these,Taxus cuspidatehad the highest energy content(19.7 kJ/g),followed byEuonymus verrucosus(18.9 kJ/g)and the lowest energy content was recorded forPopulus cathayana(16.7 kJ/g).Carbohydrate,the most important component,accounted for 84.8±1.9%of total weight of dry matter,ranging from 82.2 to 88%of dry weight.CP accounted for 7.5±1.7%of dry matter weight,ranging from 4.5%to 10.3%and varying most widely among the 15 species.Higher CP content was recorded forAralia elata(10.3%),Euonymus alatus(9.5%),T.cuspidate(9.3%),andUlmus pumila(9.3%).In contrast,Acer ukurunduensecontained the lowest CP content(4.5%),followed byP.cathayana(4.8%).Crude lipid(CL)content was statistically similar for all plant species.Ten of the fifteen species had CL content ranging from 5 to 7%(x=7±1.5%).The highest CL content was recorded forSaussurea manshurica(9.8%)andP.cathayana(9.7%),and the lowest content forU.pumila(5.4%),Acer tschonoskii(5.6%)andAcer tegmentosum(5.6%).NFC content averaged 20.1±4.8%and varied widely among the 15 species.S.manshuricaexhibited the highest NFC content(up to 28.8%)andFraxinus mandshurica,a tall tree,had the lowest(13.5%).The 15 species had relatively stable but low ash content,averaging 0.8±0.3%.Tilia amurensishad the lowest ash content(0.3%).The following species had ash content>1%:Equisetum hyemale(1.2%),P.cathayana(1.1%),A.ukurunduense(1%),A.tschonoskii(1%),T.cuspidate(1%)andEuonymus macropterus(1%).DMD varied little by species(Table 2),averaging 46.1±4.8%.Thirteen of the fifteen species had DMD>40%.E.verrucosus(38.7%)andA.ukurunduense(39.4%)had the lowest DMD levels,whileS.manshurica(54.6%),A.elata(53.2%),andE.hyemale(52.1%)had the highest DMD levels.

Table 1 Plants species foraged by red deer in the reserve and their respective abbreviations and icons
The relationship between macronutrients and energy content
Macronutrients of plants are carbohydrate,CP and CL.These three types of nutrients can be converted into energy to fuel life activities.There was no correlation between CP content and energy content(r=0.004)in red deer forage species(Fig.1a).There was a statistically insigni ficant negative correlation between CL content and energy content(r=-0.190;Fig.1b).Carbohydrate was composed of NFC and NDF.NDF,included cellulose,hemicellulose,and lignin.These are main components of the plant cell wall and can not be used by animals other than ruminants.NDF content and energy content were significantly and negatively correlated(r=-0.343;Fig.1c).NFC mainly includes sugar(monosaccharide and starch),pectin,and organic acids.These are critical as the main energy sources and they release energy directly.NFC content and energy content were strongly and positively correlated(r=0.451;Fig.1d).
The relationship between fibers in winter diets of red deer
Fiber content varied significantly among the 15 species of forage plants(Table 2).Average NDF was 64.7±5.9%.F.mandschuricahad the highest NDF content at 71.8%,followed byE.verrucosus(71.7%)andA.ukurunduense(70.4%).Species with NDF content<60%were:S.manshurica(53.5%),E.hyemale(56.2%),T.cuspidate(57.5%),andA.elata(58%).Cellulose content averaged 35±4.9%.P.cathayanahad the highest cellulose content at 40.8%,followedbyA.elata(39.5%),andT.cuspidatehadthelowest at 20.5%.Mean hemicellulose content was 16.2±2.4%.F.mandshuricahad the highest hemicellulose content at 23.3%whileT.cuspidatehad the lowest at 11.9%.Twelve of fifteen species showed hemicellulose content ranging from 15 to 18%.Lignin contentranged from 1.6 to 25.1%(x=13.4±5.5%)and was lower than the content of celluloseorhemicelluloseacrossall15species.Thehighestvalues wererecordedforT.cuspidate,andthelowestforE.hyemale.Cellulose,hemicellulose,and lignin percentages accounted fortheNDFofthe15species(Fig.2).ThecellulosecontentofE.hyemaleaccounted for the highest proportion of NDF at>70%.Cellulose content ofTaxus cuspidateaccounted for the lowest proportion of NDF at 35.6%.The cellulose contents of the remaining 13 species ranged from 45 to 60%of NDF.Hemicellulose content was similar for all 15 species,rangingfrom20to30%exceptforF.mandshurica,whichhad over 35%.Lignin content was highly variable,ranging from 2.9(E.hyemale)to 43.6%(T.cuspidata),with the remaining 13 species ranging from 10 to 30%.
Discussion
Nutritional composition of winter diets of red deer
?
CP content is commonly used as an index of dietary quality in ungulates(Chen et al.1997).Gao and Yang(2004)studied farmed red deer and concluded that 27.3%of the newborns were weak when CP content of the fodder for pregnant red deer was 8.5%.However,offspring were healthy when CP was raised to 9.8%(ibid.).Similar results were reported for white-tailed deer by Murphy and Coates(1966).Elahi and Rouzbehan(2008)concluded that a CP content of 7–8%was required to maintain rumen microbial activity.For female red deer,a daily intake of 7%CP content is required to meet physiological nitrogen demands(Mould and Robbins 1981).In our study,average CP content was 7.5%,lower than that estimated for winter foraged of wapiti(Cervus elaphus roosevelti)in primary forest of western Washington,USA(9.2%)(Leslie et al.1984),and half of the CP content of forage consumed by red deer in mixed broadleaf-conifer forest of western France(15%)(Verheyden-Tixier et al.2008).The main reasons for these differences in CP content was variation in climate and vegetation.In western France,grasses,whose CP content was higher than that of other plants,accounted for most of the red deer winter diet.The local warm climate was conducive to the growth of gramineous plants that provided an abundant and high-protein resource for wintering red deer(Verheyden-Tixier et al.2008).Due to the deep snow accumulation typicalofsoutheastern Heilongjiang,highprotein grasses were generally unavailable to wintering red deer on our study area and consequently the CP content of their winter forage was low.
Soluble sugar is an important component of NFC.Although some reports have concluded that herbivores prefer plants containing soluble sugars,CP is not a crucial nutritional component determining forage species selection of herbivores(Bugalho and Milne 2003;Verheyden-Tixier et al.2008;Laska 2001;Tixier et al.1997;Parsons et al.2006).Verheyden-Tixier et al.(2008)reported on the nutritional selection of red deer in the mixed broadleaf-conifer forest of western France.Deer here had adequate food resources and mainly ate gramineous plants,usually avoiding conifers and ferns due to their higher lignin content and risk of toxicity despite the fact that their soluble sugar content is high(ibid.).T.cuspidateis a rare and endangered Taxus species and the only conifer in the winter diets of the red deer in the Muling forest region.Taxinol,which is obtained fromTaxus cuspidate,can inhibit cancer cell division and is considered a promising anti-cancer drug.However,the taxines also derived fromTaxusplants can harm the respiratory system of animals,and in severe cases,can cause respiratory failure and death(Shanker et al.2002;Guo et al.2010).Weaver and Broun(2004)reported that the rumen fluid of white-tailed deer had a strong detoxi fication effect on the leaves ofTaxus baccata,reducing the toxicity by 59%.In the Muling forest region,there were only few high-quality ruminant food species in winter.Therefore,red deer ateTaxus baccataduring winter at the cost of greater lignin intake and risk of poisoning.
McDonald et al.(1995)de fined nutritional assessment criteria for diets of ungulates and set a DMD threshold at 50%.Of the 15 species eaten by red deer at Muling,only three had DMD>50%.These wereE.hyemale,S.manshuricaandA.elata.This suggests that high-quality forage species for wintering red deer in the Muling forest region were relatively few.Alvarado et al.(2012)reported that plant species with higher DMD usually had higher content of other nutrients(e.g.,CP,NFC).In our study,the three plant species that had DMD>50%also had higher NFC and CP contents,consistent with the results of Alvarado et al.(2012).
The relationship between macronutrients and energy content
Simpson et al.(2003)reported that carbohydrate was an important source of energy for vegetarian insects,while CL content was insigni ficant.Simpson and Raubenheimer(2005)showed that protein can be converted into energy in some cases but at ef ficiency levels of around 15%.Here,NFC was determined as the main energy source of red deer in winter.However,the contribution of CL and CP to energy was negligible.This result supports the conclusions of Simpson et al.(2003)and Simpson and Raubenheimer(2005).
The relationship between fibers and winter diets of red deer
An increase in NDF content is usually accompanied by a reduction in the consumption of nutrients(Xu 2007).Gomez-Castro etal.(2006)considered plantswith NDF>50%as low-quality food for ungulates(Gomez-Castro et al.2006).Sosa-Rubio et al.(2004)concluded that plants with NDF content between 20 and 35%had higher digestibility(Sosa-Rubio et al.2004).Based on these results,the 15 plants species at Muling could not be considered high-quality food.We recorded low-NDF species such asE.hyemale,S.salicifolia,A.elata,andT.cuspidatethat had high NFC and DMD,consistent with Minson(1990).Minson(1990)concluded that plants with low cell wall content had higher digestible non-structural carbohydrate.Moya-Rodriguez et al.(2002)reported negative correlation between lignin and DMD,which reduced the consumption of food by ungulates.E.hyemale,S.manshurica,andA.elatahad low lignin content and high DMD.This supports the conclusion of Moya-Rodriguez et al.(2002).Therefore,we claim that these three plants species were good-quality food for red deer wintering in the reserve.

Fig.1 The relationship between vegetation energy contents and three macronutrients contents of 15 forage species in the Muling forest region
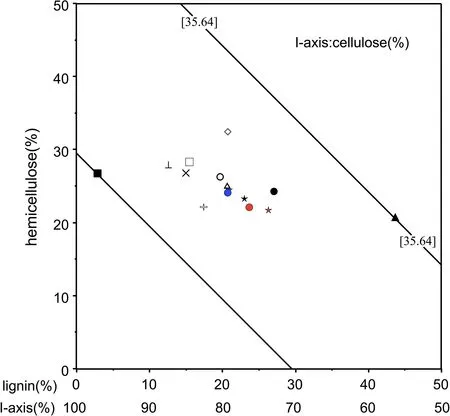
Fig.2 The composition of fibers(cellulose,hemicellulose and lignin as%of NDF)in 15 forage species by red deer taken from the Muling forest region in winter
Conclusions
There were few high-quality forage species for wintering red deer in the Muling forest region.A.elata,U.pumila,E.alatus,andT.cuspidatehad higher CP content among the 15 forage species whileE.hyemale,S.manshurica,andT.cuspidatehad higher NFC content.Based on their highest CP,NFC,DMD and lowest lignin content,E.hyemale,S.manshurica,andA.elataprovided the best nutritional value for wintering red deer.T.cuspidatewith higher CP and NFC content and higher lignin content,could also be categorized as good-quality forage for red deer.
AcknowledgementsWe are particularly grateful to Qiu Yanming,Dai Fu,Wang Daohui and other staffs at the reserve during the samples collection period.The nutritional composition was carried out by He Huan,Sun Yue,Xing Bo,whose time and competence are gratefully acknowledged.
References
Albou SD,Mitchell B,Huby BJ,Brown D(1986)Fertility in female red deer(Cervus elaphus):the effects of body composition,age and reproductive status.J Zool 209:447–460
Alvarado MS,Gonzalez-Rodriguez H,Ramirez-Lozano RG,Silva IC,Gomez-Meza MV,Correa MC,Jurado E,Dominguez-Gomez TG(2012)Chemical composition and digestion of shrubs browsed by white-tailed deer(Odocoileus virginianusTexanus).J Anim Vet Adv 11:4428–4434
Asher GW,Cox N(2013)The relationship between body-mass and puberty in young red deer(Cervus elaphus)hinds:evidence of early-life effects on permissive live-weight thresholds.Anim Reprod Sci 143:79–84
Asher GW,Stevens DR,Archer JA,Barrell GK,Scott IC,Ward JF,Littlejohn RP(2011)Energy and protein as nutritional drivers of lactation and calf growth of farmed red deer.Livest Sci 140:8–16
Berteaux D,Creˆte M,Huot J,Maltais J,Ouellet JP(1998)Food choice by white-tailed deer in relation to protein and energy content of the diet:a field experiment.Oecologia 115(1):84–92
Bugalho MN,Milne JA(2003)The composition of the diet of red deer(Cervus elaphus)in a Mediterranean environment:a case of summer nutritional constraint?For Ecol Manag 181:23–29
Chen HP,Luo LY,Xie XC,Han YZ,Wang YQ(1997)Protein nutrition in red deer(Cervus elaphus).J For Res 3:174–181
Elahi MY,Rouzbehan Y(2008)Characteriztion ofQuercus persica,Quercus infectoriaandQuercus libanias ruminant feeds.Anim Feed Sci Technol 140:78–89
Galliard JM,Sempere AJ,Bouttin JM,Laere GV,Boisaubert B(1992)Effects of age and body weight on the proportion of females breeding in a population of roe deer(Capreolus capreolus).Can J Zool 70:1541–1545
Gao XH,Yang HF(2004)Feed and nutrition of deer.China Agriculture Press,Beijing,p 93(in Chinese)
Gao YH,Zeng XY,Zhou GY,Wang GX(2011)Interspeci fic relationships of dominant populations in the Alpine Wetlands in the source region of Yangtze River.Wetl Sci 9:1–7
Gerhart KL,White RG,Cameron RD,Russell DE(1996)Body composition and nutrient reserves of Arctic caribou.Can J Zool 74:136–146
Gomez-Castro H,Nahed-Toral J,Tewolde A,Pinto-Ruiz R,Lopez-Martinez J(2006)A´reas con potencial para el establecimiento de a´ rboles forrajeros en el centro de Chiapas.Te´cnica pecuaria en Me´xico 44:219–230
Gonzalez-Hernandez MP,Silva-Pando FJ(1999)Nutritional attributes of understory plants known as components of deer diets.J Range Manag 52:132–138
Guo YB,Ren Y,Cheng F(2010)A review on anti-cancer action and clinical research of taxol.Clin J Chin Med 2:1–3(in Chinese)
Hamilton WJ,Blaxter KL(1980)Reproduction in farmed red deer.I.Hind and stag fertility.J Agric Sci 95:261–273
Laska M(2001)A comparison of food preferences and nutrient composition in captive squirrel monkeys,Saimiri sciureus,and pigtail macaques,Macacanemestrina. Physiol Behav 73:111–120
Lei X(1994)Statistical analysis on the relationship between drymatter digestibility of alfalfa and its chemical composition.J Anim Sci Vet Med 1:2–4(in Chinese)
Leslie DM,Starkey EE,Vavra M(1984)Elk and deer diets in oldgrowthforestsinWesternWashington.JWildlManag 48:762–775
Lo´pez-Pe´rez E,Serrano-Aspeitia N,Aguilar-Valde´s BC,Herrera-Corredor A(2012)Composicio´n nutricional de la dieta del venado cola blanca(Odocoileous virginianusssp.mexicanus)en Pitzotla´n,Morelos.Rev Chapingo Ser Cienc For Ambient 18:219–229
Luna-Estrada AA,Vera-Avila HR,Mora O,Anguiano-Serrano B,Vasquez Pela´ez CG,Shimada A(2006)Effect of pre-mating nutritional status in red deer(Cervus elaphusscoticus)hinds on thesex ratio oftheiroffspring.SmallRumin Res 65:154–160
McDonald P,Edwards RA,Greenhalgh JFD,Morgan CA(1995)Animal nutrition,5th edn.Longman Press,Harlow,p 607
Minson DJ(1990)Forage in ruminants nutrition.Academic Press,San Diego,pp 310–331
Mould ED,Robbins CT(1981)Nitrogen metabolism in elk.J Wildl Manag 45:323–334
Moya-Rodriguez JG,Ramirez RG,Foroughbakhch R(2002)Seasonal changes in cell wall digestion of eight browse species from northeastern Mexico.Livest Res Rural Dev 14:1–7
Murphy DA,Coates JA(1966)Effects of dietary protein on deer.Trans North Am Wildl Nat Resour Conf 31:129–139
Oddy VH,Robards GE,Low SG(1978)Prediction of in vivo dry matter digestibility from the fibre nitrogen content of a feed.Analyst 103:406–409
Ouyang X,Wang QF,Wang J,Wang Y(2000)Comparison of testing daily intake and dry-matter digestion coefficient of Tibetan goat.J Southwest Natl Coll Nat Sci Ed 26:188–191(in Chinese)
Parker KL,Gillingham MP,Hanley TA,Robbins CT(1999)Energy and protein balance of free-ranging black-tailed deer in a natural forest environment.Wildl Monogr 143:1–48
Parker KL,Barboza PS,Stephenson TR(2005)Protein conservation of female caribou(Rangifer tarandus):effects of decreasing diet quality during winter.J Mammal 86:610–622
Parker KL,Barboza PS,Gillingham MP(2009)Nutrition integrates environmental responses of ungulates.Funct Ecol 23:57–69
Parlak OA,Gokkus A,Hakyemez BH,Baytekin H(2011)Shrub yield and forage quality in Mediterranean shrublands of west Turkey for a period of one year.Afr J Agric Res 6:1726–1734
Parsons MH,Lamont BB,Davies SJJF,Kovacs BR(2006)How energy and coavailable foods affect forage selection by the western grey kangaroo.Anim Behav 71:765–772
Shanker K,Pathak NKR,Trivedi VP,Chansuria JPN,Pandey VB(2002)An evaluation of toxicity ofTaxus baccataLinn.(Talispatra) in experimental animals.J Ethnopharmacol 79:69–73
Simpson SJ,Raubenheimer D(2005)Obesity:the protein leverage hypothesis.Obesity 6:133–142
Simpson SJ,Bately R,Raubenheimer D(2003)Geometric analysis of macronutrient intake in humans:the power of protein?Appetite 41:123–140
Sosa-Rubio EE,Perez-Rodriguez D,Ortega-Reyes L,Zapata-Buen fil G (2004)Evaluacion del potencial forrajero de arboles y arbustos tropicales para la alimentacion de ovinos.Te´cnica pecuaria en Me´xico 42:129–144
Suttie JM,Hamilton WJ(1983)The effect of winter nutrition on growth of young Scottish Red deer(Cervus elaphus).J Zool 201:153–159
Tixier H,Duncan P,Scehovic J,Yani A,Gleizes M,Lila M(1997)Food selection by European roe deer(Capreolus capreolus):effects of plant chemistry,and consequences for the nutritional value of their diets.J Zool 242:229–245
van Soest Robertson JB,Lewis BA(1991)Methods for dietary fiber,neutral detergent fiber,and non-starch polysaccharides in relation to animal nutrition.J Dairy Sci 74:3583–3597
Verheyden-Tixier H,Renaud PC,Morellet N,Je Jamot,Besle JM,Dumont B(2008)Selection for nutrients by red deer hinds feeding on a mixed forest edge.Oecologia 156:715–726
Weaver JD,Broun DL(2004)Incubation of European yew(Taxus baccata)with white-tailed deer(Odocoileus virginianus)rumen fluid reducestaxineaconcentrations.VetHum Toxicol 46:300–302
Xu M(2007)Nutritional regulation of rumen health and carbohydrate energy utilization of ruminants.Ph.D.thesis,Northwest Agriculture and Forestry University Press,Yangling,p 3(in Chinese)
 Journal of Forestry Research2018年3期
Journal of Forestry Research2018年3期
- Journal of Forestry Research的其它文章
- Vacuum press drying studies on two fast-growing Indian wood species
- Water-repellent ef ficiency of thermally modi fied wood as affected by its permeability
- Predicting intensity of white-tailed deer herbivory in the Central Appalachian Mountains
- A taxonomic review of the genus Miaenia Pascoe,1864(Coleoptera:Cerambycidae)from South Korea with a new record of M.tonsa(Bates)
- A study on JA-and BTH-induced resistance of Rosa rugosa‘Plena’to powdery mildew(Sphaerotheca pannosa)
- Using UAVs for detection of trees from digital surface models