
茶树CsGPX基因克隆及干旱胁迫下的表达分析
2018-05-18 01:25乔新荣张继英
江苏农业科学 2018年8期
乔新荣,张继英
(信阳农林学院,河南信阳464000)
谷胱甘肽过氧化物酶(glutathione peroxidase,简称GPX)是生物机体内重要的抗氧化酶,它可以清除机体内的H2O2及脂质过氧化物,阻断活性氧自由基对机体的进一步损伤,保护细胞免受氧化胁迫,提高植物的抗逆能力[1-2]。谷胱甘肽过氧化物酶也可以作为信号分子,与激素交联,调节植物的生长发育[3-4]。因此,对谷胱甘肽过氧化物酶进行开发和利用具有重要的意义,现已从烟草、柑橘、拟南芥、大白菜、番茄、豌豆、水稻、龙眼等许多植物中分离和鉴定[5]。
茶树[Camellia sinensis(L.)O.Kuntze]属于山茶科双子叶植物,茶叶具有高效的抗癌、抗衰老、抗辐射、抗氧化、清除人体自由基、降血糖和血脂等一系列药理功能。我国茶区地形分布复杂,包括平原、丘陵、山地和高原。这些地区经常出现季节性干旱、高温、低温等逆境,导致茶芽萌发迟、生长瘦弱,造成茶产量和品质下降。本研究拟克隆茶树CsGPX基因,并对其进行生物信息学方面及干旱胁迫下的转录表达进行初步分析,旨在为今后通过基因工程研究CsGPX基因的表达,探究茶树依赖CsGPX途径清除活性氧的分子基础,以期为鉴定和选育茶树抗逆品种提供分子水平上的理论依据。
1 材料与方法
1.1 试验材料
本研究所使用的茶树材料为河南省信阳茶叶示范园区信阳群体种。选取健壮茶树的新梢幼叶,提取总RNA并反转录得到cDNA模板,利用RT-PCR技术克隆CsGPX基因片段。
选取长势一致的茶树健壮植株枝条,于人工气候箱内进行Hoagland完全营养液水培,每3 d更换1次营养液,1周后进行干旱处理。用渗透势为-1.0 MPa的聚乙二醇6000模拟干旱胁迫进行培养,分别收集胁迫 0、3、6、9、12、18、24 h的茶树幼叶,液氮冷冻后保存,分别提取总RNA并反转录得到不同的cDNA模板,利用real-time PCR技术检测CsGPX基因转录表达量。
1.2 试验试剂
DNA分子量标准、Taq DNA聚合酶、dNTPs、克隆载体pMD18-T、DNA回收试剂盒、质粒提取试剂盒、反转录试剂盒,均购自天根生化科技(北京)有限公司;Trizol试剂购自Invitrogen公司;荧光定量试剂盒SYBR Premix Ex Taq,购自TaKaRa公司;在Primer Premier 5.0中进行引物设计;引物合成及测序由南京金斯瑞生物科技有限公司完成。
1.3 引物序列
CsGPX基因克隆引物为F1:5′-GTCAATGTTGCCTCACA ATGTGG-3′,R1:5′-CAGCTTCTTCACGTCCTTCTCAAT-3′。茶树β-Actin基因real-time PCR引物为 F2:5′-AGGGTAT GCTCCTCCTCAT-3′,R2:5′-TCAGCAGTGGTGGTGAACAT-3′。CsGPX基因 real-time PCR引物为 F3:5′-GAGCCAGGG AATAATGAGC-3′,R3:5′-ATTGGAGCAGCATTCTCAC-3′。
1.4 CsGPX基因片段的信息学分析
分别利用 DNAMAN 6.0、Clustal X 1.81及 MEGA 5.2对CsGPX基因片段进行氨基酸序列推断、保守序列分析、进化树构建。
2 结果与分析
2.1 茶树CsGPX基因片段的克隆
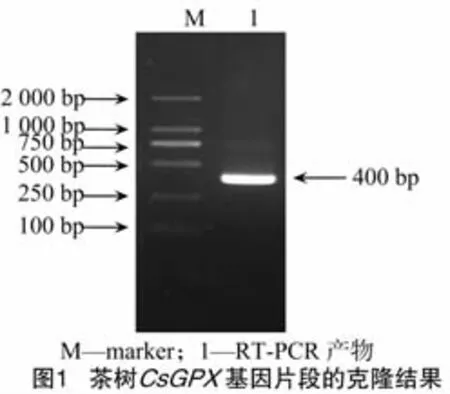
根据其他物种GPX基因的保守序列设计特异扩增引物F1和R1,利用Trizol试剂提取茶树幼叶总RNA,检测RNA的质量后,进行反转录获得cDNA第1链,以此为模板,经PCR扩增获得1条大小约为400 bp的目的片段(图1)。反应程序为94℃预变性5min;94℃变性30 s,55℃退火30 s,72℃延伸50 s,32个循环;72℃延伸10min。扩增产物经1%琼脂糖凝胶电泳后,用DNA凝胶回收试剂盒将目的片段回收纯化,连接于克隆载体pMD18-T上,热激转化大肠杆菌DH5α。菌落PCR扩增鉴定后,挑取阳性克隆进行测序,经测序克隆到的基因片段长387 bp,利用DNAMAN 6.0软件推断其编码129个氨基酸(图2)。
2.2 茶树CsGPX基因片段的生物信息学分析
从GenBank和PeroxiBase数据库中下载拟南芥(AtGPX 1~AtGPX 8)、杨树(PtGPX 1、2、5、8,PtGPX 6-1、PtGPX6-2)、水稻 (OsGPX1)、玉米 (ZmGPX1)、番茄 (SlGPX)、小 麦(TaGPX)等蛋白质的氨基酸序列,利用 DNAMAN 6.0和Clustal X 1.81软件,与推断的CsGPX基因片段编码的氨基酸序列进行同源性比对。由图3可知,CsGPX编码的氨基酸序列具有GPX蛋白保守特征序列,即具有3个保守结构域(Ⅰ、Ⅱ、Ⅲ)以及酶活性中心的3个保守的半胱氨酸(C)残基。初步推断其克隆的基因序列是茶树CsGPX基因片段。
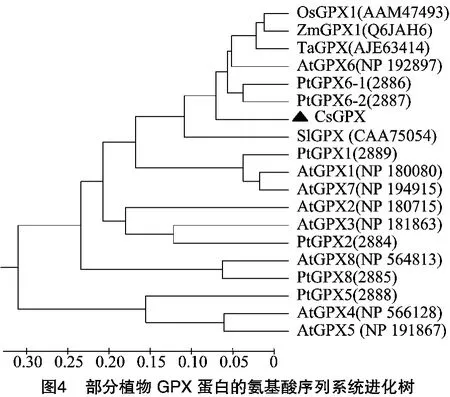
利用 MEGA 5.2软件,按 Neighbor-Joining方案对CsGPX及部分植物GPX蛋白的氨基酸序列进行分子系统进化分析,由图4可知,CsGPX保守序列区域与拟南芥AtGPX6、杨树PtGPX6-1、杨树PtGPX6-2等亲缘关系较近。
2.3 CsGPX基因在干旱胁迫下的转录表达分析
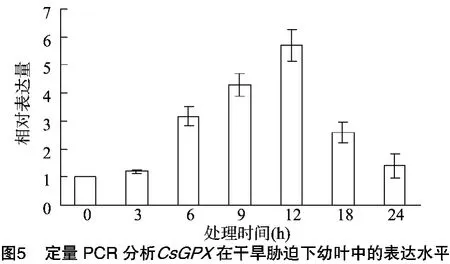
使用ABI7300荧光定量PCR仪进行表达量分析,采用20μL反应体系:10μL SYBR Premix Ex Taq,正、反向引物(10μmol/L)各0.5μL,0.4μL ROX Reference Dye(50×),2.0μL cDNA模板,6.6μL ddH2O。反应程序为95℃预变性90 s;PCR反应:95℃ 15 s,60℃ 40 s,共40个循环,每个样品进行3次重复检测,使用2-ΔΔCT法求得待测样品的相对表达量。分别提取干旱胁迫处理 0、3、6、9、12、18、24 h茶树幼叶的RNA,反转录为 cDNA,以此为模板,利用 Primer Premier 5.0设计茶树β-Actin定量PCR引物为F2、R2,CsGPX基因定量PCR引物为F3、R3,以茶树β-Actin片段为内参,检测CsGPX在干旱胁迫下的转录表达量变化。由图5可知,随着干旱胁迫处理时间的延长,相对表达量呈现先升后降的变化趋势,在处理12 h时,CsGPX转录相对表达量达到最大值,约是对照的6倍。随后又降低,24 h后逐渐恢复至处理前的水平。
3 结论与讨论
本研究中克隆茶树CsGPX基因片段所编码的氨基酸序列具有植物GPX的3个特征保守结构域,且含有GPX活性中心的3个保守的半胱氨酸残基,因此,所克隆的CsGPX基因属于植物GPX家族成员。且与已报道的AtGPX6、PtGPX6、TaGPX等亲缘关系较近。初步研究CsGPX的转录表达受干旱胁迫诱导,表明CsGPX参与茶树抗旱信号转导途径。已报道AtGPX6定位于线粒体和胞质,在多种组织的整个发育过程中都高度表达[3],盐、铁、铝胁迫增强了AtGPX6的mRNA表达水平[6]。利用水杨酸(SA)、生长素(IAA)、脱落酸(ABA)、茉莉酸(JA)等处理后,也引起AtGPX6转录表达量的增加[3]。在AtGPX6基因的启动子区含有ABA、SA、IAA响应元件。另外,定位于小麦叶绿体中的TaGPX基因转入拟南芥超表达后,表现出对盐、H2O2、ABA胁迫的强耐性,且盐、H2O2、ABA信号转导通路中的关键调节子基因(SOS1、RbohD、ABI1/ABI2)的表达水平也在转基因株系中改变[7]。干旱胁迫下水稻的GPX酶活性也增强[8]。这些植物GPX的功能研究为CsGPX的深入研究提供了参考。将来对CsGPX基因的全长进行克隆,将其原核表达,检测其亚细胞定位。也将进一步研究CsGPX酶活性在干旱胁迫中的变化以及其作用底物的特异性,为阐明CsGPX在干旱胁迫信号转导中的作用机制奠定基础。

参考文献:
[1]Mittler R.Oxidative stress,antioxidants and stress tolerance[J].Trends in Plant Science,2002,7(9):405-410.
[2]Yang X D,LiW J,Liu JY.Isolation and characterization of a novel PHGPx gene in Raphanus sativus[J].Biochimica et Biophysica Acta,2005,1728(3):199-205.
[3]Rodriguez Milla M A,Maurer A,Rodriguez Huete A,et al.Glutathione peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through diverse signaling pathways[J].Plant Journal,2003,36(5):602-615.
[4]Gaber A,Ogata T,Maruta T,et al.The involvement of Arabidopsis glutathione peroxidase8 in the suppression of oxidative damage in the nucleus and cytosol[J].Plant&Cell Physiology,2012,53(9):1596-1606.
[5]肖 蓉,罗慧珍,张小娟,等.干旱和盐胁迫条件下枣树谷胱甘肽过氧化物酶基因(ZjGPX)的差异表达及功能分析[J].中国农业科学,2015,48(14):2806-2817.
[6]Sugimoto M,Sakamoto W.Putative phospholipid hydroperoxide glutathione peroxidase gene from Arabidopsis thaliana induced by oxidative stress[J].Genes&Genetic Systems,1997,72(5):311-316.
[7]Zhai C Z,Zhao L,Yin L J,et al.Two wheat glutathione peroxidase genes whose products are located in chloroplasts improve salt and H2O2tolerances in Arabidopsis[J]. PLoS One,2013,8(10):e73989.
[8]Goswami A,Banerjee R,Raha S.Drought resistance in rice seedlings conferred by seed priming:role of the anti-oxidant defense mechanisms[J].Protoplasma,2013,250(5):1115-1129.
猜你喜欢
黑龙江大学自然科学学报(2022年4期)2022-11-17
河北科技师范学院学报(2022年2期)2022-08-26
乡村地理(2018年2期)2018-09-19
中国塑料(2016年3期)2016-06-15
天然产物研究与开发(2016年6期)2016-06-05
湖南农业(2016年3期)2016-06-05
音乐天地(音乐创作版)(2016年11期)2016-02-05
分析测试学报(2015年6期)2016-01-13
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
集美大学学报(自然科学版)(2015年4期)2015-02-28
