
茶树叶绿素荧光参数叶位差异及其稳定性分析
2018-05-14 11:32唐敏翟秀明罗红玉邬秀宏侯渝嘉
南方农业·上旬 2018年11期
唐敏 翟秀明 罗红玉 邬秀宏 侯渝嘉

摘 要 2017年,以福鼎大白茶为试验对象,比较了不同叶位茶树叶片叶绿素荧光参数的差异及稳定性。结果表明:茶树新梢上部叶片光启动速率快,对强光相对敏感,荧光参数稳定性较差。随叶位下移,叶片逐渐成熟,到第5叶时,其PSⅡ反应中心最大光能转换效率、潜在活性、实际光化学效率和电子传递速率均处于较高水平,且所测数据变异系数相对较小,表现出较强的稳定性。建议在进行茶树叶绿素荧光参数测定时,选择芽下第5叶或邻近的功能叶为试验对象以提高数据稳定性,从而更准确地反应茶树光合生理特征。
关键词 茶树;叶片;叶绿素;荧光参数;叶位差异;稳定性
中圖分类号:Q945.11 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2018.31.003
叶绿素荧光分析技术作为植物胁迫生理研究中的一种无损探针,能快速准确地揭示植物对光能的吸收、传递、耗散、分配等特征。基于荧光数据的分析,能有效反应不同植物或不同品种在抗性、光合潜能、区域适应性等方面的差异[1-4]。然而,叶绿素荧光参数的差异不仅存在于同一植物的不同品种之间,也存在于同一品种的不同叶位之间[5-7];即使用同一品种的同一叶位叶片进行荧光参数测定,也会因叶片成熟度差异,生理活动的活跃程度差异、对环境的适应能力差异等原因,导致重复测定时数据波动性较大,从而影响试验结果的准确性[8]。笔者以福鼎大白茶为研究对象,拟通过对不同叶位主要荧光参数的差异及数据稳定性进行分析,以增强试验对象选择时的科学性,从而提高试验数据的稳定性和试验结果的可重复性。
1 材料与方法
1.1 试验地点
试验地为重庆市农业科学院茶叶研究所种植资源圃,位于重庆市永川区茶山竹海街道。
1.2 试验材料
供试品种为三年生福鼎大白茶,按双行双株侧窝种植,大行距150 cm,小行距40 cm,穴距33 cm,常规管理。选取生长健壮,长势一致,无病虫害的当年生茶树枝条芽下1~6叶作为测定对象。
1.3 试验方法
试验于2017年8月下旬进行,为尽量减少叶绿素荧光参数日变化产生的影响,测定时间均安排在9:00—12:00。主要荧光参数和光响应曲线采用PAM-2500便携式调制叶绿素荧光仪测定,测定前叶片进行暗适应20~30 min。所有指标测定均重复4次,取平均值。
1.4 数据处理
所测数据用Excel进行整理。差异显著性分析与多重比较(P=0.05)用SPSS Statistics 22 (SPSS Statistics IBM)计算。变异系数按如下公式计算:变异系数(C.V.)=标准差/平均值×100%。
2 结果与分析
2.1 不同叶位PSⅡ活性差异
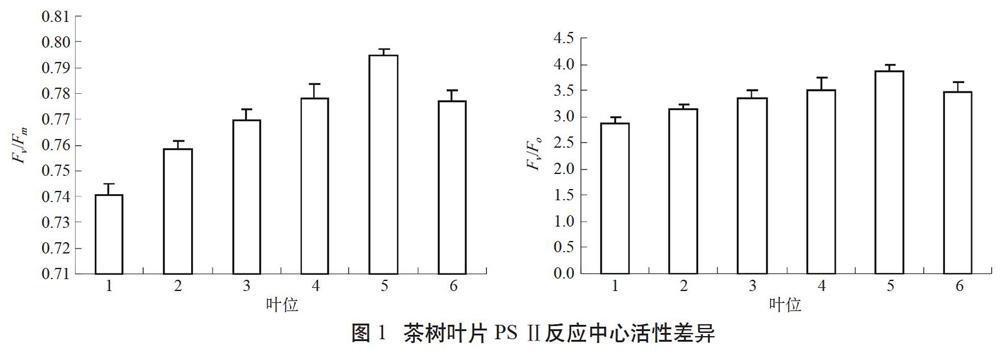
Fv/Fm与Fv/Fo是常用于度量植物潜在光合能力和PSⅡ活性的两个重要指标,前者反映植物叶片PSⅡ反应中心最大光能转换效率,后者反映PSⅡ的潜在活性[9]。由图1可知,茶树新梢不同叶位叶片充分暗适应后,Fv/Fm和Fv/Fo随叶位变化呈先升后降趋势,在第5叶时达到最大值,第6叶开始降低,但仍显著高于1~3叶。说明茶树叶片成熟度对Fv/Fm和Fv/Fo具有显著影响,且4~6叶PSII反应中心的光能转化效率和潜在活性相对较高。
2.2 不同叶位光化学效率与电子传递能力差异
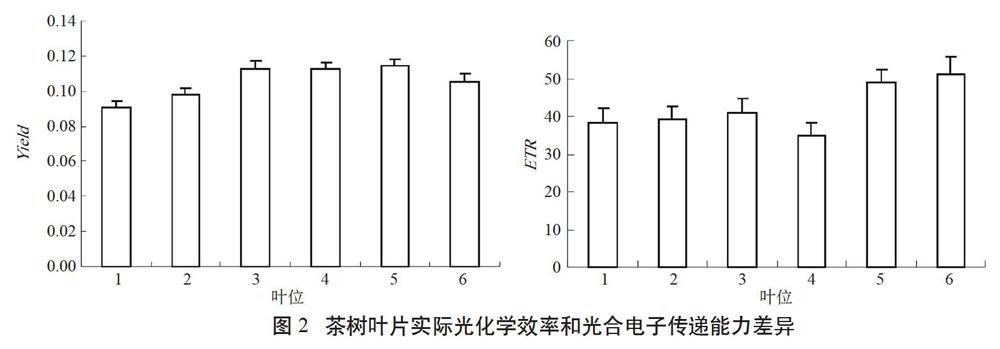
Yield是PSⅡ实际光化学效率,它是由光适应下的初始荧光(Fo′)、 稳态荧光产量(Ft)和光适应下的最大荧光(Fm′)按Yield=(Fm′-Ft)/Fm′计算所得,用于反映植物叶片在照光条件下反应中心部分关闭时的光化学效率。PSⅡ的电子传递速率(ETR),主要用于反映光合机构电子传递能力,与其生理状况和生长的环境因素有关[10]。由图2可知,叶片Yield随叶位变化呈先升后降趋势,所测叶片中,3~5叶Yield最高,但与其他叶位叶片的差异并未达到统计学意义的显著水平;而ETR以第5叶和第6叶最高,且与其它叶片差异显著,这说明成熟叶片的实际光化学效率和电子传递能力均强于未成熟叶片,4~6叶为茶树光合作用的主要功能叶。
2.3 不同叶位光响应曲线及光合启动速率差异
众多研究表明,在未达到光饱和之前ETR与PAR(光合有效辐射)的强度呈线性关系,当光能过量时,ETR和入射的PAR不再呈线性关系,要低于按线性关系估计的值,甚至由于发生光抑制而开始降低[11-13]。通过比较不同叶位叶片的光响应曲线,能直观地反映不同叶位电子传递能力的差异。本试验采用Eilers and Peeters的公式[1]分别对不同叶位叶片的光响应曲线进行拟合,结果见图3。图3显示,不同叶位叶片通过PSII的电子传递速率随着光强的增加而不断增加,当光强增大到900~1 200 μmol/(m2·s)时,ETR增速减缓直至不再增加或降低;相同PAR强度下,ETR随叶片成熟程度逐渐增加,其中第5叶和第6叶光合电子传递能力显著强于1~4叶。
光响应曲线初始斜率(alpha)及最小半饱和光强(lk)可用来反映植物的光化学启动速率和对强光的耐受能力[14]。不同叶位叶片alpha和lk计算结果见图4。图4显示,第1叶光响应曲线的初始斜率显著高于4~6叶,而最小饱和光强以第5叶和第6叶最高,且与第1叶差异显著。这说明茶树新梢上部叶片对光照敏感,光化学启动速率较快,而下部叶片逐渐成熟,光化学反应趋于稳定,对强光具有较强的耐受能力。
2.4 不同叶位主要荧光参数稳定性分析
由于茶树生长是一个动态变化的过程,不同成熟度叶片生理活动的活跃程度存在明显差异,是影响所测数据稳定性的重要因素之一。本试验比较了不同叶位叶片叶绿素荧光数据的波动情况,结果列于表1。
由于枝条顶端叶片生长势力最强,生理活动旺盛,变化较快,因此第1、2叶位的变异系数普遍较大,且明显高于中下部,随叶片逐渐成熟,其生理活动趋于稳定,所测数据重现性较高。
3 讨论
同一茶树品种不同叶位的叶绿素荧光参数具有明显差异,表明茶树叶绿素荧光参数的差异不仅受到基因型的影响,还与被测叶片的成熟度有关。试验结果显示,除PSⅡ实际光化学效率外,叶位对主要荧光参数均具有显著影响,表现为:随叶位下移,叶片逐渐成熟,其PSⅡ反应中心最大光能转换效率、潜在活性、实际光化学效率和电子传递速率逐渐升高,在第5叶时达到最高水平,之后部分参数略有下降,这说明茶树新梢第5叶为茶树光合生理的主要功能叶,之后叶片光合功能开始下降,但仍强于未成熟叶片。
由于不同叶位成熟度不同,因此其光化学反应启动速率和对环境的适应能力也不相同,试验结果显示,枝条顶端的叶片(第1叶)光响应曲线的初始斜率显著高于成熟叶片,且最小半饱和光强最小,这说明未成熟叶片对光照敏感,光化学启动速率较快,但对强光的耐受能力较弱,而成熟叶片则与之相反。
由于茶树生长具有明显的顶端优势,位于枝条顶端的叶片生长势最强,生理活动旺盛,变化幅度大,加之未成熟叶片对环境适应能力较弱,因此所测荧光数据在同一品种同一叶位的单株之间仍有较大波动性,其变异系数明显高于成熟叶片。随叶片生长,其光合机构逐渐成熟,光化学过程趋于稳定,相同条件下所测数据稳定性更强。因此,建议在进行茶树叶绿素荧光参数测定时,选择芽下第5叶或邻近的功能叶为试验对象,以便更准确地反应茶树光合生理特征。
参考文献:
[1] Eilers P, Peeters J. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton[J]. Ecological Modelling, 1988, 42(3): 199-215.
[2] 李晓,冯伟,曾晓春.叶绿素荧光分析技术及应用进展[J].西北植物学报,2006,26(10):2186-2196.
[3] 王少先,李再军,王雪云,等.不同烟草品种光合特性比较研究初报[J].中国农学通报,2005,21(5):
245-257.
[4] 郭小强,赵明,李少昆.不同玉米自交系光合特性的研究[J].玉米科学,1997,5(3):46?49.
[5] 李刚华,薛利红,尤娟,等.水稻氮素和叶绿素SPAD叶位分布特点及氮素诊断的叶位选择[J].中国农业科学,2007(6):131-139.
[6] 冯伟,李晓,王永华,等.小麦叶绿素荧光参数叶位差异及其与植株氮含量的关系[J].作物学报,2012,38(4):657-664.
[7] 李艳大,汤亮,张玉屏,等.水稻冠层光截获与叶面积和产量的关系[J].中国农业科学,2010,43(16):
3296-3305.
[8] 童淑媛,宋凤斌,徐洪文.玉米不同叶位叶片SPAD值的变化及其与生物量的相关性[J].核农学报,2008,22(6):869-874.
[9] 張守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[10] 张阿宏,齐孟文,张晔晖.调制叶绿素荧光动力学参数及其计量关系的意义和公理化讨论[J].核农学报,2008,22(6):909-912.
[11] 彭予咸,胡留洁,邓敏,等.植物生长调节剂对茶树新梢叶绿素荧光特性的影响[J].西南农业学报,2013,26(2):514-519.
[12] 唐敏,翟秀明,姚永红,等.不同茶树品种(系)绿素荧光参数季节性差异研究[J].西南农业学报,2015,28(1):79-83.
[13] 桂仁义,刘亚迪,郭小勤,等.不同剂量137Cs-γ辐射对毛竹幼苗叶片叶绿素荧光参数的影响[J].植物学报,2010,45(1):66-72.
[14] 叶子飘.光合作用对光和CO2响应模型的研究进展植物[J].生态学报,2010,34(6):727-740.
猜你喜欢
茶道(2022年3期)2022-04-27
少儿科学周刊·少年版(2021年17期)2021-01-17
少儿科学周刊·儿童版(2021年17期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
华东师范大学学报(自然科学版)(2019年5期)2019-11-11
新农业(2018年3期)2018-07-08
电机与控制学报(2018年9期)2018-05-14
求知导刊(2017年22期)2017-10-13
艺术评鉴(2017年6期)2017-06-01
女性天地(2016年10期)2017-04-25
