
生长激素调控PCOS大鼠血清与卵巢组织中血管因子表达的研究
2018-05-09 05:51王施元李肖肖于彩虹王珊尹晓静刘羽王芳芳王瑞静刘梅梅
生殖医学杂志 2018年5期
王施元,李肖肖,于彩虹,王珊,尹晓静,刘羽,王芳芳,王瑞静,刘梅梅*
(1.哈尔滨医科大学,哈尔滨 150000;2.济宁医学院附属医院妇产科,济宁 272029;3.石家庄市第四医院妇产科,石家庄 050000;4.新乡市中心医院妇产科,新乡 453000;5.朝阳市中心医院妇产科,朝阳 122000;6.哈尔滨医科大学附属第二医院,哈尔滨 150000)
多囊卵巢综合征(PCOS)是由神经、内分泌系统及代谢紊乱而引起的一种以排卵功能紊乱和高雄激素血症为特征的疾病[1]。临床上,PCOS主要表现为月经不调、不孕、多毛和肥胖,在育龄期妇女中发病率高达4%~12%。有研究显示,未及时治疗的PCOS患者将增加Ⅱ型糖尿病、心血管疾病、高血脂、冠心病及子宫内膜癌等疾病的发生风险[2]。目前PCOS发病机制尚不清楚,多数学者认为PCOS患者存在下丘脑-垂体-卵巢性腺轴的功能调节障碍,也有研究认为生长激素(GH)对下丘脑-垂体-卵巢轴具有一定的调节作用,影响卵巢的成熟及卵泡的周期性发育。同时,PCOS患者也存在GH分泌异常的现象。目前PCOS的研究除在妇科内分泌领域方面外,也涉及免疫学、基因学及卵巢血管异常生成等领域。有研究证实[3],PCOS患者血清中血管相关因子——血管内皮生长因子(VEGF)水平升高,提示PCOS可能与血管生成异常改变相关。在我们之前关于PCOS大鼠的研究结果也提示[4],PCOS的发生可能与血管生成密切相关。已知,血管形成相关因子内皮抑素(ES)、内皮素-1(ET-1)及血小板反应素-1(TSP-1)的相互作用参与了血管生成。在本研究中我们拟在原实验基础上,通过来曲唑灌胃法诱导PCOS大鼠模型,检测血清与卵巢组织中ES、ET-1及TSP-1的表达情况,及应用GH后PCOS大鼠血清及卵巢组织中ES、ET-1及TSP-1表达水平的变化,试图从血管生成方面探讨GH在PCOS发生发展的调节作用。
材料和方法
一、材料
1.实验动物:选取6周龄SD 雌性大鼠70只,体质量(160±10)g,由哈尔滨医科大学附属第二医院动物实验中心提供[合格证号:SCXK(黑)2015-001)]。
2. 主要试剂和药物:来曲唑(Letrozole,江苏瑞恒医药);羧甲基纤维素(胶州浆糊粉厂);生长激素(长春金赛药业);内皮抑素抗体(Endostatin Antibody)、血小板反应素-1抗体(THBS1 Antibody、多聚体抗兔IgG-HRP(武汉博士德);内皮素-1抗体(Rabbit Anti-ET1 antibody)(北京博奥森);DAB显色试剂盒(北京中杉金桥)。
二、实验方法
1. 动物模型制备与分组:饲养条件:室温20℃~25℃,相对湿度40%~70%,颗粒饲料和自来水喂养,自由饮食,12 h/12 h昼夜明暗交替(日光灯照明)。将70只大鼠适应性喂养2 d后,随机分为两组,对照组20只,模型组50组。模型组给予来曲唑1 mg/(kg·d),溶于1% 羧甲基纤维素(CMC)中,灌胃21 d;同期对照组给予等量的1% 的CMC灌胃;所有入组动物均每日行阴道涂片监测动情周期的变化。雌性动物在正常生理周期内或者非妊娠的情况下,每间隔一段时间都会有一次发情现象,这种情况被称为雌性动物的发情周期。将雌鼠固定,暴露阴道口,将蘸有生理盐水的棉签伸入阴道口,轻轻来回转动,涂抹在载玻片上,显微镜下进行发情周期观察并记录。灌胃21 d后,模型组随机抽取10只处死,采取颈总动脉血并摘取双侧卵巢组织。通过检测大鼠动情周期鉴定及对比、显微镜下观察卵巢的形态学变化评估造模效果。当造模成功率>80%时,将模型组余下大鼠随机分为生长激素组(GH组 20只)和PCOS组(PCOS组 20只),GH组皮下注射rhGH,剂量为1 U/(kg·d)(给药剂量按照人与大鼠公斤体重折算系数计算),溶于生理盐水中;PCOS组与对照组注射等量的生理盐水,每日1次,连续21 d。
2. 标本采集与处理:不同组大鼠喂养相应时间后,用止血钳固定大鼠四肢,剔除一侧颈部皮毛,逐层分离暴露颈总动脉,切断动脉取血3 ml,置于离心管中,室温静置30 min 后置于离心机中,以3 000 r/min,离心15 min,收集血清,分装后置于-20℃冰箱待检。取大鼠双侧卵巢,剔除周围脂肪组织并称重,迅速置于10%甲醛溶液中固定,以卵巢横切面作为待检平面行石蜡包埋。以上课题实验均经过我院伦理委员会论证。将石蜡包埋标本制成常规切片,切片脱蜡至水后,95℃水浴中抗原修复,加入1∶100稀释的ES一抗、1∶200稀释的ET-1一抗、1∶100稀释的TSP-1一抗。依次加入即用型二抗作用,DAB显色。以抑制阳性片为阳性对照,用PBS代替一抗作为阴性对照。
3. 检测指标:(1)大鼠动情周期鉴定及对比;(2)模型组动物卵巢形态学鉴定;(3)ELISA法检测大鼠血清ES、ET-1、TSP-1水平;(4)免疫组化法检测各组大鼠卵巢组织中的ES、ET-1及TSP-1的表达,细胞浆中出现棕黄色或褐色颗粒为阳性,根据阳性细胞染色深浅和阳性细胞百分率作为结果进行判定。显色深浅:不显色为0分,浅黄色为1分,棕黄色为2分,深褐色为3分。阳性细胞百分率:无阳性细胞为0分,阳性细胞数<25 %为1分,阳性细胞数在25%~50%为2分,阳性细胞数>50%为3分。二者得分相加,0分表示阴性(-),1~2分表示弱阳性(+),3~4分表示阳性(++),5~6分表示强阳性(+++)。结果判断在双盲下进行,切片结果均由两位以上病理专家审阅评定。
三、统计学分析
大鼠血清ES、ET-1、TSP-1表达的统计学分析采用SPSS 17.0软件。因大鼠随机分成3组,每组均有不同的处理,故需首先采用单因素方差分析,其后运用LSD-t检验进行两两间的多重比较,P<0.05为差异有统计学意义;免疫组织化学结果采用非参数秩和检验分析,P<0.0001为差异有统计学意义。
结 果
一、大鼠动情周期鉴定及对比
模型组大鼠的动情周期在应用来曲唑12 d后均失去规律,处于动情间期时相:镜下少量白细胞和中间层细胞,未见角化细胞和上皮细胞。提示模型组无排卵。对照组大鼠仍存在正常的动情周期变化,出现发情前期、发情期、发情后期等。
二、卵巢形态学鉴定
对10只随机处死的模型组大鼠首先通过HE染色来观察卵巢的形态学变化。可见大鼠卵巢中的二级卵泡结构性萎缩,没有黄体出现。以此判断PCOS造模成功。
三、3组大鼠血清ES、ET-1、TSP-1的表达
与对照组相比,PCOS组大鼠血清ES、ET-1表达升高,而TSP-1表达下降(P<0.05);与PCOS组相比,GH组大鼠血清ES、ET-1表达下降,但未达到正常对照组水平(P<0.05),GH组大鼠血清TSP-1表达升高,与对照组水平相接近(P>0.05)(表1)。
四、ES在3组大鼠卵巢组织中的表达比较
3组大鼠卵巢组织中,ES阳性表达的对照组有7例,PCOS组有19例,GH组有15例(图1)。与对照组相比,PCOS组大鼠卵巢颗粒细胞及卵泡膜细胞中ES表达水平增高(P<0.0001);与PCOS组相比,GH组大鼠卵巢颗粒细胞及卵泡膜细胞ES表达降低(P=0.0066),但未达到正常对照组水平(P=0.0098)(表2)。
五、ET-1在3组大鼠卵巢组织中的表达
3组大鼠卵巢组织中,ET-1阳性表达者在对照组有8例,PCOS组有18例,GH组有15例(图2)。与对照组相比,PCOS组大鼠卵巢颗粒细胞及卵泡膜细胞中ET-1表达水平增高(P<0.0001);与PCOS组相比,GH组大鼠卵巢颗粒细胞及卵泡膜细胞ET-1表达降低(P=0.0134),但未达到正常对照组水平(P=0.0390)(表2)。
六、TSP-1在3组大鼠卵巢组织中的表达
3组大鼠卵巢组织中,TSP-1阳性表达者在对照组有18例,PCOS组有7例,GH组有20例(图3)。与对照组相比,PCOS组大鼠卵巢颗粒细胞及卵泡膜细胞中TSP-1表达水平降低(P=0.0001);与PCOS组相比,GH组大鼠卵巢颗粒细胞及卵泡膜细胞TSP-1表达增加(P<0.0001),与正常对照组水平相接近(P=0.6559)(表2)。

A:对照组(++); B:PCOS组(+++); C:GH组(+)图1 大鼠卵巢组织中ES的表达(免疫组织化学 ×400)
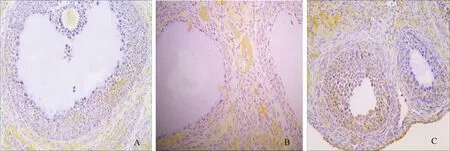
A:对照组(++); B:PCOS组(+++); C:GH组(+)图2 大鼠卵巢组织中ET-1的表达(免疫组织化学 ×400)

A:对照组(++); B:PCOS组(-); C:GH组(+)图3 大鼠卵巢组织中TSP-1的表达(免疫组织化学 ×400)
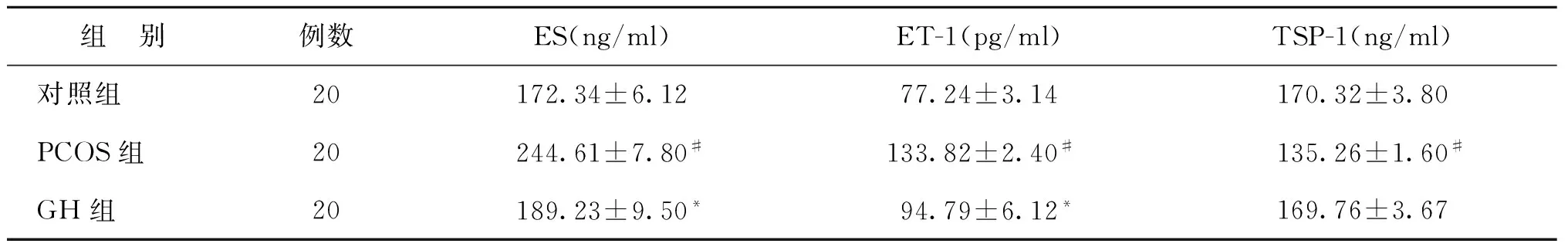
组 别例数ES(ng/ml)ET-1(pg/ml)TSP-1(ng/ml)对照组20172.34±6.1277.24±3.14170.32±3.80PCOS组20244.61±7.80#133.82±2.40#135.26±1.60#GH组20189.23±9.50*94.79±6.12*169.76±3.67
注:与对照组比较,#P<0.05;与PCOS组比较,*P<0.05

表2 大鼠卵巢组织中ES、ET-1及TSP-1的表达
讨 论
一、GH与PCOS
GH主要由腺垂体前叶嗜酸性细胞分泌,调节糖、蛋白质、脂肪的代谢,其主要作用是促进生长和调节代谢。尽管PCOS的病因尚不明确,但多数学者普遍认为PCOS患者存在下丘脑-垂体-卵巢性腺轴功能调节障碍,且涉及了多种内分泌生物分子的表达、代谢及信号通路异常[5]。研究发现GH可能通过对性腺轴的调节,影响卵巢的成熟及其卵泡的周期性发育。目前,GH在PCOS中促排卵作用成为现在研究的热点,GH可作用于卵巢上GH受体,与卵泡生成和黄素化、卵母细胞成熟、排卵等密切相关,其主要通过依赖和非依赖促性腺激素的机制发挥调节功能。
我们的实验结果显示,在给予GH后PCOS大鼠血清ES、ET-1的表达下降,TSP-1表达上升;在卵巢组织血管相关因子表达方面,表现为PCOS组大鼠卵巢组织中ES、ET-1高表达,TSP-1低表达;在应用GH后,ES、ET-1的表达下降,但并没有达到正常状态,推测在PCOS中,GH可能通过调节大鼠血清与卵巢组织中ES及ET-1的表达而发挥作用,但抗血管生成因子与GH的关系目前尚不清楚,且GH在PCOS中具体的调节机制仍需进一步的研究。
目前,关于GH的研究大多数集中于PCOS患者的促排卵中效果,而在PCOS中的具体作用机制尚未明确。本实验研究发现,给予GH后,ES、ET-1在PCOS大鼠血清与卵巢组织中表达下降,而TSP-1的表达上升。血管生成相关因素在PCOS中的作用以及GH在PCOS中调节血管因子表达平衡的作用机制,均需要深入的研究探讨,将为PCOS的治疗开辟新的思路。
二、ES与PCOS
ES是一种特异性的抑血管生成因子,选择性的作用于参与形成新生血管过程中的小血管内皮细胞,其主要通过促进内皮细胞凋亡、抑制血管内皮增殖及抑制细胞因子诱导的血管内皮细胞的迁移等途径有效抑制新生血管的形成[6]。ES多与VEGF同时被探讨,研究发现,ES能选择性抑制内皮细胞上一氧化氮合酶(eNOS) 的磷酸化,诱使eNOS失活,从而抑制VEGF介导的血管新生和血管通透性增加作用[7]。已有研究表明,血管生成需要多种细胞因子的调节。我们之前的研究发现[8],PCOS患者血清ES水平与VEGF呈正相关,表明二者在调节血管生成方面有着重要相关性。异常升高的VEGF促进机体血管生成的同时,为维持内环境的稳定,机体同时分泌大量的血管生成抑制剂如ES以抗衡高水平的血管生成趋势,从而导致ES在PCOS患者体内增多,但这种代偿作用仍不能使卵巢微环境达到正常的生理状态,从而可能导致卵巢囊性增生,常表现为包膜增厚,及卵泡囊的血管增生引起的卵巢增大、充血为主[9]。
我们的实验结果显示,与对照组比较,PCOS组大鼠血清与卵巢组织中ES的表达升高,与我们之前的研究结果相符[10]。我们的研究结果还提示[8]PCOS患者由于ES与VEGF的表达失衡而导致卵巢间质血管化、卵泡周围血管新生,卵巢血流丰富呈现多囊状态,从而导致PCOS的病理生理损害进一步发展。可见,ES在PCOS 的发生发展中可能发挥着重要的血管抑制作用。本次实验中还发现,GH组大鼠血清与卵巢组织中ES的表达下降,但未达到正常对照组水平,据此推测,GH可能通过调节大鼠血清与卵巢组织中ES与VEGF的表达,进而在PCOS的发生发展中发挥作用,其具体调节机制仍需进一步研究。
三、ET-1与PCOS
ET-1是多肽类家族中一员,可涉及多个生物学过程[11],研究证实ET-1通过与卵巢组织内的相应受体结合从而调节血管的舒缩活动,以实现对卵巢血流的调控[12],还能通过刺激NADPH(还原型辅酶Ⅱ)氧化酶的活性来增加体内活性氧的产生,使NO所介导的内皮舒张作用受抑制,导致血管舒缩障碍[13]。有研究表明,PCOS患者血清中ET-1表达升高,NO水平降低[14],对内皮有损害作用的ET-1和对血管有保护作用的活性分子NO表达水平的失衡而增加了血管内皮损伤。Imbar等[15]研究发现ET-1 mRNA在PCOS患者颗粒黄体细胞中的表达是排卵正常女性的两倍,由此认为其可能与PCOS患者的排卵障碍有关。
我们的实验结果显示,PCOS大鼠血清与卵巢组织中ET-1的表达高于对照组,GH作用后ET-1的表达下降,但未达到正常对照组水平。也有学者认为PCOS患者ET-1血清表达水平与正常女性并无明显差别[16],这与本实验的结果并不相同,可能与研究对象的不同(本实验是动物实验)、我们研究例数较少等因素有关。据我们的实验结果推测,在PCOS的发生发展中,GH可能通过调节大鼠血清与卵巢组织中ET-1的表达而发挥一定的作用。
四、TSP-1与PCOS
TSP-1抑制血管生成包括抑制血管内皮细胞的迁移及诱导内皮细胞凋亡及通过抑制VEGF发挥作用。TSP-1缺乏的小鼠和对照组相比,产仔数减少,故TSP-1影响生殖系统。我们之前的另一项临床研究表明,TSP-1的表达在PCOS患者血清中较正常人群低,且PCOS患者中合并有胰岛素抵抗者的TSP-1表达水平较非胰岛素抵抗者高[17],这与本次实验研究结果相一致。另有资料[18]表明,正常卵巢组织中卵巢颗粒细胞可以产生TSP-1,其主要作用卵巢颗粒细胞及黄体细胞,从而调节血管生成以及啮齿类动物卵巢卵泡和黄体发育,且与卵泡的生长发育具有一定的相关性。
我们的实验结果显示,与对照组相比,PCOS组大鼠血清与卵巢组织中TSP-1表达下降;与PCOS组相比,GH作用后大鼠血清与卵巢组织中TSP-1的表达升高,但GH作用后的TSP-1和对照组相比,却不具有统计学意义。TSP-1可通过影响内皮细胞迁移、增殖与凋亡、以及改变血管生成因子VEGF活性来抑制血管生成[19-20]。根据以上实验结果推测,TSP-1其发挥抑制血管的作用与VEGF相互依赖,二者可能共同参与了PCOS的发生发展,平衡制约着卵巢组织中血管生成的调节;GH可能通过调节TSP-1和VEGF的相互表达而在PCOS的发生发展中起到一定作用,但其具体作用机制仍需进一步的研究证实。
在目前关于GH调控PCOS大鼠血清与卵巢组织中相关血管因子表达的研究中,样本量较小。关键在于GH调控PCOS的基础研究不足,这是目前解决动物实验结果瓶颈所在。从目前的结果来看GH和PCOS是具有一定的相关性的。
本研究评估了PCOS大鼠经过GH治疗后,血清与卵巢组织中相关血管因子表达,结果显示:与对照组相比,PCOS组大鼠血清及卵巢组织中ES和ET-1均为高表达,TSP-1低表达;与PCOS组大鼠相比,GH组大鼠血清及卵巢组织中ES和ET-1表达降低,而TSP-1表达升高。但仍需要更大样本量的研究,以及GH和相关血管因子与PCOS关系的更为深层的研究。这可能将为PCOS的发病机制以及诊疗思路带来新进展。
【参考文献】
[1] 檀丽,彭巧捷,陈丽春. 维生素D辅助治疗对多囊卵巢综合征内分泌代谢相关指标及胰岛素抵抗的影响[J]. 数理医药学杂志,2016,29:1497-1499.
[2] Gao H,Meng J,Xu M,et al. Serum heat shock protein 70 concentration in relation to polycystic ovary syndrome in a non-obese Chinese population[J/OL]. Plos One,2013,8:e67727.
[3] Ng EHY,Chan CCW,Yeung WSB,et al. Comparison of ovarian stromal blood flow between fertile women with normal ovaries and infertile women with polycystic ovary syndrome[J].Hum Reprod,2005,20:1881-1886.
[4] 原振宇,刘梅梅,张艳华,等. 两种多囊卵巢综合征动物模型比较及来曲唑诱导的动物模型卵巢组织中血管内皮生长因子和内皮素1的表达[J]. 国际免疫学杂志,2015,38:124-128.
[5] 付竹霓,曲洪美,于晓艳,等. 多囊卵巢综合征患者血清生长激素释放多肽及脂联素的检测及意义[J]. 中华临床医师杂志:电子版,2012,6:4051-4053.
[6] 王韧,侯立军. VEGF和内皮抑素对血管生成作用的研究进展[J]. 重庆医学,2011,40:2582-2585.
[7] Hajitou A,Grignet C,Devy L,et al. The antitumoral effect of endostatin and angiostatin is associated with a down-regulation of vascular endothelial growth factor expression in tumor cells[J]. FASEB J,2002,16:1802-1804.
[8] 刘梅梅,丁慧,尹晓静,等. 多囊卵巢综合征患者血清血管内皮生长因子、内皮抑素水平及对卵巢间质血流的影响研究[J]. 中国全科医学,2017,20:448-453.
[9] 赵鸿娟,王煜,王丽岩,等. 多囊卵巢综合征患者卵泡液中分子异常的研究进展[J]. 生殖医学杂志,2015,24:166-169.
[10] 刘梅梅,李佩玲,尹晓静,等. 血管生成相关因子内皮抑素和血小板反应素-1在多囊卵巢综合征大鼠卵巢组织中的表达及意义[J]. 疑难病杂志,2016,15:1178-1181.
[11] Khimji AK,Rockey DC. Endothelin--biology and disease[J]. Cell Signal,2010,22:1615-1625.
[12] Mancina R,Barni T,Calogero AE,et al. Identification,characterization,and biological activity of endothelin receptors in human ovary[J]. J Clin Endocrinol Metab,1997,82:4122-4129.
[13] Mancini F,Cianciosi A,Reggiani GM,et al. Endothelial function and its relationship to leptin,homocysteine,and insulin resistance in lean and overweight eumenorrheic women and PCOS patients: a pilot study[J]. Fertil Steril,2009,91:2537-2544.
[14] Schiffrin EL. Vascular endothelin in hypertension[J]. Vascul Pharmacol,2005,43:19-29.
[15] Imbar T,Klipper E,Greenfield C,et al. Altered endothelin expression in granulosa-lutein cells of women with polycystic ovary syndrome[J]. Life Sci,2012,91:703-709.
[16] Foltyn W,Strzelczyk J,Marek B,et al. Selected markers of endothelial dysfunction in women with polycystic ovary syndrome[J]. Endokrynol Pol,2011,62:243-248.
[17] Liu M,Gao J,Zhang Y,et al. Serum levels of TSP-1,NF-κB and TGF-β1 in polycystic ovarian syndrome(PCOS) patients in northern China suggest PCOS is associated with chronic inflammation[J]. Clin Endocrinol(Oxf),2015,83:913-922.
[18] Rhee KS,Park JH,Kim HS,et al. Antiangiogenic treatment with thrombospondin-1 enhances primary tumor radiation response and prevents growth of dormant pulmonary micrometastases after curative radiation therapy in human melanoma xenografts[J]. Cancer Res,2003,63:4055-4061.
[19] Lawler PR,Lawler J. MolecularbBasis for the regulation of angiogenesis by thrombospondin-1 and-2[J]. Cold Spring Harb Perspect Med,2012,2:a006627.
[20] Claus C,Kazuichi M,Felix B,et al. Thrombospondin 1 inhibits inflammatory lymphangiogenesis by CD36 ligation on monocytes[J]. J Exp Med,2011,208:1083-1092.
猜你喜欢
中老年保健(2021年11期)2021-11-30
中国生殖健康(2020年8期)2021-01-18
保健与生活(2020年18期)2020-09-26
解放军医学杂志(2020年7期)2020-09-02
中国生殖健康(2018年1期)2018-11-06
中国卫生标准管理(2015年25期)2016-01-14
安徽医科大学学报(2015年9期)2015-12-16
医学研究杂志(2015年11期)2015-06-10
科技视界(2014年29期)2014-08-15
中国民间疗法(2013年8期)2013-01-25
