
外源钙和硫化氢对镉胁迫下平邑甜茶幼苗根系活性氧代谢和线粒体特性的影响
2018-05-09 05:59曹慧潘利姜倩倩邹岩梅束怀瑞
华北农学报 2018年2期
曹慧,潘利,姜倩倩,邹岩梅,束怀瑞
(1.潍坊学院 山东省高校生物化学与分子生物学重点实验室,山东 潍坊 261061;2.潍坊一中,山东 潍坊 261041;3.国家苹果工程技术研究中心,山东 泰安 271018)
由于工业“三废”和生活垃圾的不得当处置,污水灌溉,以及农药化肥的不合理使用,果园土壤已不同程度受到镉等重金属的污染。镉是毒性很强的重金属,可对果品安全生产及人类健康构成严重威胁[1]。高浓度镉能够诱导植物产生大量活性氧(Reactive oxygen species,ROS),对细胞膜、蛋白和核酸等生物大分子具有破坏作用,严重时导致细胞死亡。植物根系中的ROS主要来源于线粒体,反过来线粒体又是ROS作用的靶子,它可提高线粒体膜通透性,引起细胞色素c(Cyt c)释放;Cyt c是线粒体呼吸链上传递电子的载体,线粒体Cyt c的流失可直接引起电子传递受阻或中断,刺激线粒体膜通透性进一步开放,从而产生更多的ROS,继而引发一系列下游反应,最终导致细胞受损坏死[2]。镉胁迫使平邑甜茶根系线粒体MTP增大,膜电位(Δψm)和 Cyt c含量下降,H2O2含量升高,最终导致细胞程序性死亡(Programmed cell death,PCD)的发生[3]。
近几年,越来越多的研究表明新型内源性气体信号分子硫化氢(Hydrogen sulfide,H2S) 具有复杂的生物学活性,参与多种生理和病理过程,其在植物中的作用逐渐被发现。在调节植物生长发育方面,H2S可促进种子萌发[4],调控保卫细胞[5],增强叶片光合作用[6],延迟衰老等[7]。在植物抗逆方面,H2S可以通过减小叶片气孔孔径,激活抗氧化系统,调控离子通道活性及基因表达等参与植物对盐碱[8]、重金属[9]、温度[10]等逆境胁迫的响应。并且,H2S与ROS、NO、Ca2+和植物激素等信号路径存在相互作用,共同调控植物生长发育和逆境胁迫响应过程[11-12]。Ca2+作为植物细胞的第二信使,胞内Ca2+水平改变,转导多种生理过程。Ca2+/CaM参与H2S介导的烟草悬浮细胞耐热性的提高[13],在谷子响应Cr6+胁迫的过程中,Ca2+调控H2S的产生,外源H2S促进Ca2+响应基因的表达,Ca2+作为初始信号与H2S相互作用[12]。
关于H2S对植物Cd胁迫的缓解作用已有一些报道,但多以草本植物或悬浮细胞为研究对象。果树生长在受镉污染的土壤中,根系首当其害,但国内外对根系的研究远不如对地上部的研究广泛和深入。平邑甜茶(MalushupehensisRehd.)是优良的苹果砧木资源,抗性较强,在生产上具有重要的应用价值[14]。本试验以平邑甜茶幼苗为材料,探究H2S能否缓解Cd胁迫对平邑甜茶幼苗根系造成的损伤,利用外源Ca2+以及Ca2+螯合剂EGTA、质膜Ca2+通道阻断剂La3+和钙调素拮抗剂CPZ,研究了平邑甜茶幼苗活性氧代谢和线粒体特性的变化,以研究证明Ca2+在H2S介导重金属胁迫信号传导中的作用,从而为提高果树抗逆应用研究提供理论依据。
1 材料和方法
1.1 材料与处理
精选饱满、均匀的平邑甜茶种子,用饱和漂白粉溶液消毒15 min,蒸馏水洗净后浸泡24 h,4 ℃冰箱层积35 d,待种子露白后,播种于12 cm×18 cm营养钵,基质为草炭、珍珠岩和蛭石按照3∶1∶1的比例配制。每钵播种2粒种子,待幼苗长至5~6片真叶,选取整齐一致的幼苗移入Hoagland营养液(pH值6.5)中进行水培,每2 d更换一次营养液。当幼苗生长至12~13片真叶时,用硫酸镉(CdSO4,200 μmol/L)处理,同时在营养液中加入H2S供体硫氢化钠(NaHS,购自Sigma公司)或氯化钙(CaCl2)/钙抑制剂处理,钙离子专一螯合剂为EGTA,钙离子通道阻断剂为LaCl3或钙调素拮抗剂为CPZ。
试验处理为对照CK:正常营养液;T1:200 μmol/L Cd胁迫处理;T2:200 μmol/L Cd+200 μmol/L NaHS处理;T3:200 μmol/L Cd+200 μmol/L NaHS+10 mmol/L EGTA处理;T4:200 μmol/L Cd+200 μmol/L NaHS+50 μmol/L LaCl3处理;T5:200 μmol/L Cd+ 200 μmol/L NaHS + 50 μmol/L CPZ处理;T6:200 μmol/L CdSO4+10 mmol/L CaCl2处理。每处理3次重复,处理后第2天取根系进行活性氧代谢和线粒体特性等生理指标测定;处理后第8天取幼苗植株测定株高、茎粗和干鲜质量等生长指标。采用软件 SPSS和Excel 2010进行数据统计分析和图表绘制。
1.2 测定方法
植株生物量测定:株高(茎基部到生长点)用直尺测量;茎粗(茎基部)用游标卡尺测量;地上部和地下部鲜质量、干质量用称量法和烘干法测量。
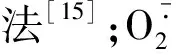
线粒体的提取和线粒体特性检测参照苏宏等[17]的方法。线粒体膜通透性的检测:配制缓冲液(220 μmol/L甘露醇,70 μmol/L蔗糖,4.2 μmol/L琥珀酸钠,pH值7.2)制备线粒体悬浮液,并用考马斯亮蓝法测定蛋白质含量后调整悬浮液蛋白浓度为0.3 mg/mL,将悬浮液置于20 ℃恒温箱中保温2 min后,紫外分光光度计检测540 nm处的吸光度变化。线粒体膜电位测定:配制缓冲液(250 μmol/L蔗糖,2 μmol/L Hepes,0.5 μmol/LKH2PO4,4.2 μmol/L琥珀酸钠,pH值7.4)制备线粒体悬浮液,并调整悬浮液蛋白浓度为0.3 mg/mL。加入罗丹明123(1 mg/mL)在25 ℃恒温箱中保温30 min孵育后,荧光分光光度计(激发波长为505 nm,发射波长为534 nm)上检测荧光强度,每个样品测定3次,每次间隔5 min,样品荧光强度取3次的平均值。线粒体细胞色素c/a测定:用牛血清白蛋白(0.2 %)制备线粒体悬浮液,调整悬浮液蛋白含量为0.5 mg/mL。紫外分光光度计检测550 nm和630 nm处的吸收值,2种波长的吸收值之比即为Cyt c/a。
2 结果与分析
2.1 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗生长的影响
如表1所示Cd胁迫导致平邑甜茶幼苗的株高、茎粗、地上部干、鲜质量,地下部干、鲜质量显著下降,明显抑制幼苗生长。Cd胁迫下分别添加外源H2S供体NaHS和CaCl2均缓解了Cd对幼苗生长的抑制,株高、茎粗和植株干鲜质量均显著增加,二者缓解水平相当;Cd胁迫下添加NaHS的同时添加钙离子专一螯合剂EGTA,钙离子通道阻断剂LaCl3或钙调素拮抗剂CPZ抑制了外源H2S对平邑甜茶幼苗生长的缓解作用,株高、茎粗、地上部干质量和地下部干、鲜质量都比Cd胁迫下仅添加NaHS的显著下降(表1)。
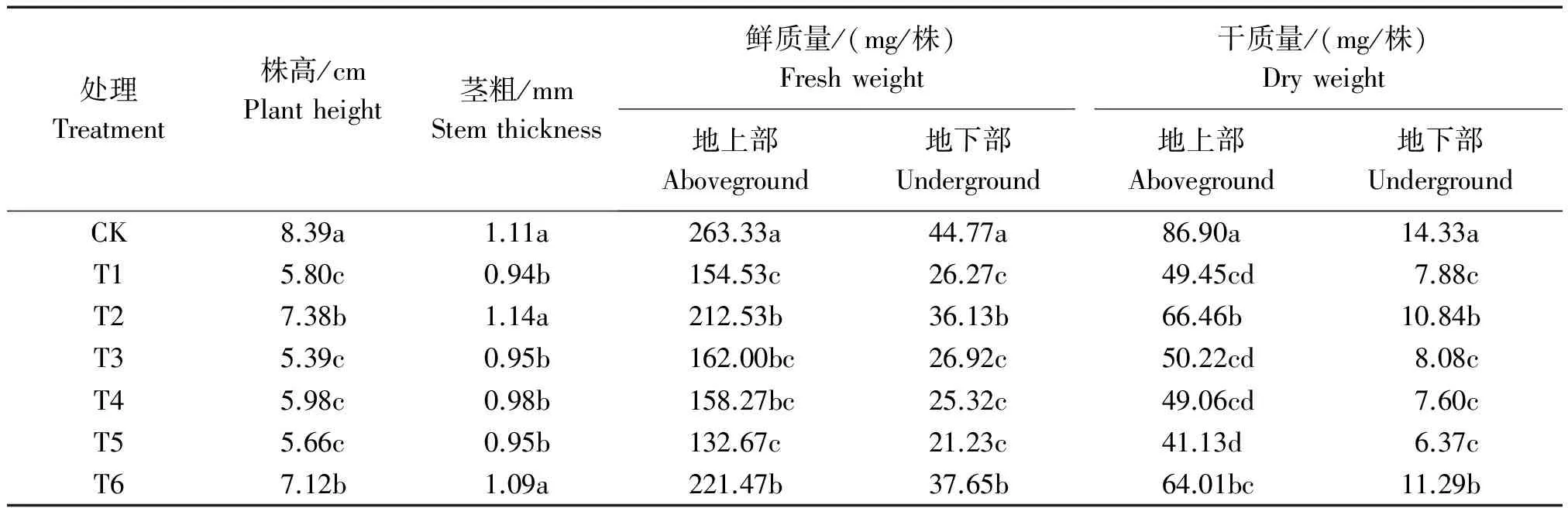
表1 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗生长的影响Tab.1 Effects of Ca2+ and H2S on growth of Malus hupehensis Rehd. seedlings under Cd stress
注:同列数值后不同字母表示差异性达5%显著水平。图1-4同。
Note:Different letters within the same column indicate significant difference at 5% level. The same as Fig.1-4.
2.2 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗根系细胞死亡的影响
伊文思蓝(Evans blue)是可透过受损的细胞膜将细胞染成蓝色的非侵透性染料,被细胞截留的伊文思蓝量可用来反映细胞的受损程度,染色越深根系活力越低,死亡细胞数量越多[3]。如图1所示,200 μmol/L CdSO4处理平邑甜茶根系48 h,可导致根系细胞截留伊文思蓝量显著增加,说明大量根系细胞死亡。Cd胁迫下添加NaHS,在外源H2S的作用下,根系细胞死亡数量比单独Cd处理降低了40.4%。Cd胁迫下添加外源Ca2+也降低了根系细胞死亡数量,与Cd+NaHS处理的水平相当,而比单独Cd处理的降低了39.3%,差异显著。而Cd胁迫下添加NaHS的同时添加EGTA、LaCl3或CPZ的处理根系细胞死亡数量又高于Cd+NaHS的处理。
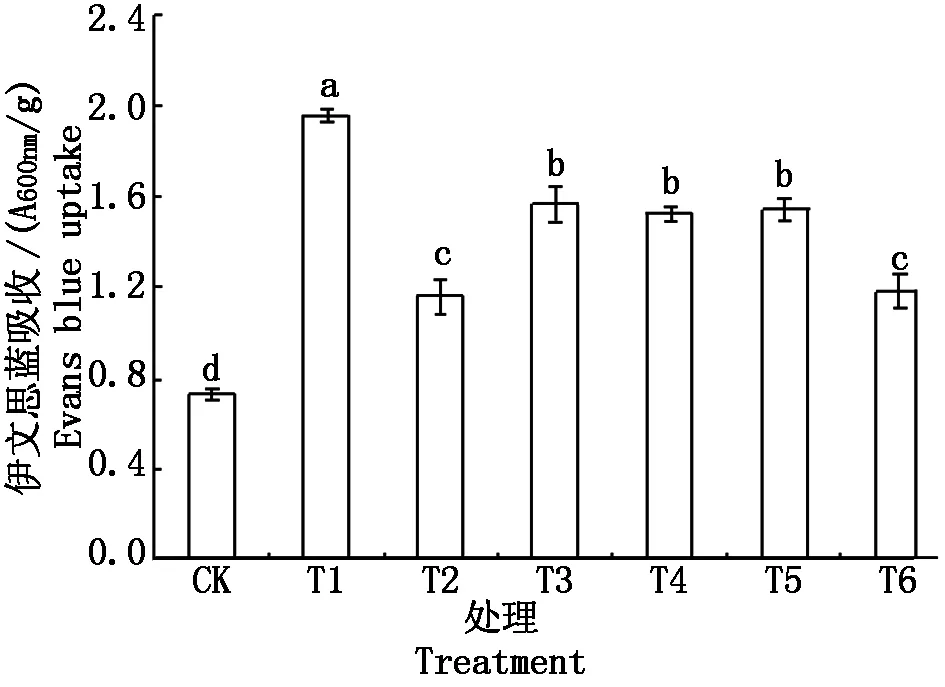
图1 Ca2+和H2S对 Cd胁迫下平邑甜茶幼苗根系细胞死亡数量的影响Fig.1 Effect of Ca2+ and hydrogen sulfide on the cell death quantity in roots of Malus hupehensisRehd. seedlings under Cd stress
2.3 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗根系产生速率、H2O2和MDA含量的影响
由图2-B可知,相同处理下,平邑甜茶根系H2O2含量(以鲜质量计)变化与超氧阴离子产生速率的变化基本一致:Cd胁迫下,根系活性氧爆发,H2O2含量保持在较高水平;Cd+NaHS和Cd+CaCl2处理的H2O2含量分别比单独Cd处理的显著降低了30.7%和29.6%,二者水平相当;Cd+ NaHS+EGTA、 Cd+NaHS+LaCl3或Cd+NaHS+CPZ处理的H2O2含量比Cd+NaHS处理的又分别提高了30.3%,24.0%和30.0%,差异显著。
Cd胁迫下,苹果幼苗根系内MDA含量(以鲜质量计)显著提高,比对照增加了45.0%;添加外源H2S显著缓解了Cd胁迫下MDA在根系中的积累,MDA含量比单独Cd胁迫处理降低了25.9%;Cd+ NaHS +EGTA、 Cd+ NaHS +LaCl3或Cd+NaHS+CPZ处理抑制了单独H2S的作用,与单独Cd胁迫处理水平相当。Cd胁迫下添加外源Ca2+也抑制了MDA产生,MDA含量比单独Cd胁迫处理降低了16.8%(图2-C)。
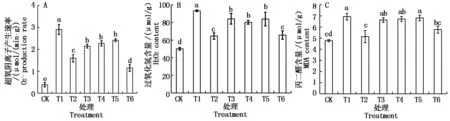
图2 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗根系产生速率、H2O2和MDA含量的影响Fig.2 Effect of Ca2+ and hydrogen sulfide on the production rate, H2O2 content and MDA content in roots of Malus hupehensis Rehd. seedlings under Cd stress
2.4 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗根系抗氧化酶活性的影响
Cd胁迫条件下,平邑甜茶幼苗根系SOD活性(以鲜质量计)明显升高,比对照高74.7%。Cd胁迫下添加NaHS或CaCl2处理的均进一步提高了根系SOD活性,分别比Cd单独处理的增加了31.8%和41.5%。Cd+NaHS+EGTA、Cd+NaHS+LaCl3或Cd+NaHS+CPZ处理的抑制了H2S的作用,SOD活性较Cd+NaHS处理的显著下降,与单独Cd胁迫处理水平相当(图3-A)。
由图3-B、C可知,POD和CAT活性(以鲜质量计)的变化趋势与SOD相似,即Cd胁迫下显著高于对照,Cd+ NaHS和Cd+CaCl2处理的进一步提高,Cd+NaHS+EGTA、Cd+NaHS+LaCl3或Cd+NaHS+CPZ处理的显著抑制了H2S的作用,与单独Cd胁迫处理水平相当。
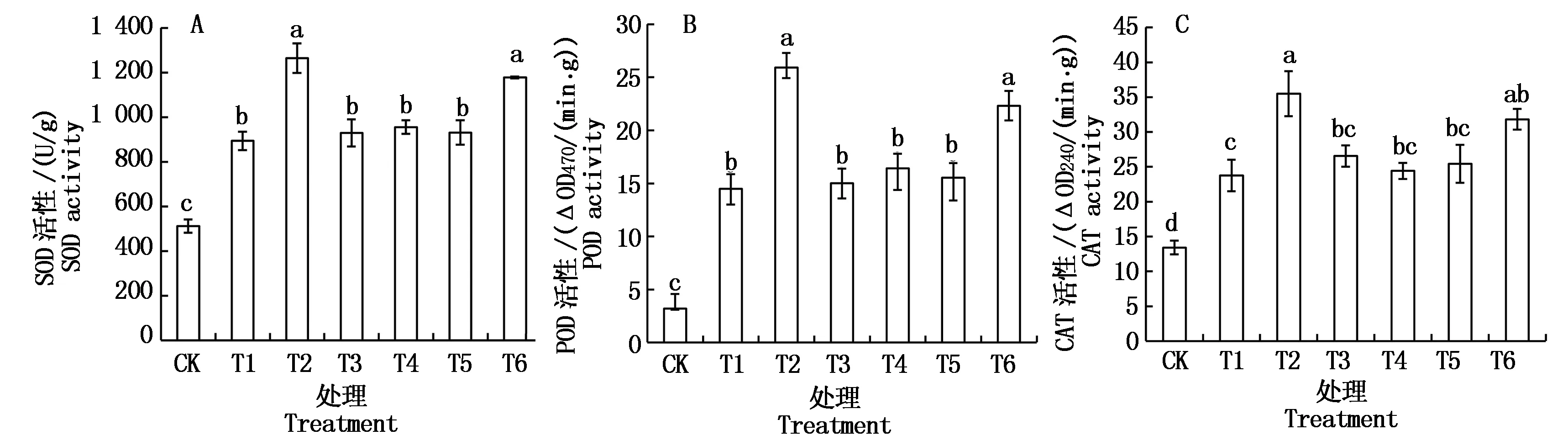
图3 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗根系抗氧化酶活性的影响Fig.3 Effect of Ca2+ and hydrogen sulfide on the antioxidant enzymes activities in roots of Malus hupehensis Rehd. seedlings under Cd stress
2.5 Ca2+和H2S对Cd胁迫下平邑甜茶幼苗根系线粒体特性的影响
线粒体膜通透性转变通道(Mitochondrial permeability transition pore,MPTP)的开放程度决定膜通透性(MPT)的转变,MPTP的过度开放使MPT不断增大,线粒体膜蛋白在540 nm处的吸光度减小。如图4-A,在单独Cd处理下根系线粒体膜蛋白吸光度显著低于对照。Cd胁迫下NaHS或CaCl2处理稳定了Cd胁迫下根系线粒体膜的通透性,Cd+ NaHS和Cd+CaCl2处理下MPTP开放程度分别比Cd处理降低了21.0%,18.8%。Cd+NaHS+EGTA、Cd+NaHS+LaCl3或Cd+NaHS+CPZ处理的不同程度上抑制了H2S的作用,其中Ca2+专一螯合剂EGTA,Ca2+通道阻断剂LaCl3的抑制效果显著,MPTP开放程度比Cd+NaHS处理的又分别提高了9.72%和9.14%。钙调素拮抗剂CPZ抑制效果稍差,MPTP开放程度比Cd+NaHS处理的提高了8.02%。
线粒体膜电位(Δψm)是评价线粒体功能的特征指标,MPTP的过度开放会导致Δψm不可逆地改变。Cd处理的根系线粒体Δψ与对照相比显著降低。Cd胁迫下添加NaHS提高了线粒体Δψ,与单独Cd处理差异显著;Cd胁迫下添加NaHS的同时添加Ca2+专一螯合剂EGTA,Ca2+通道阻断剂LaCl3和钙调素拮抗剂CPZ抑制了H2S的作用,根系线粒体Δψ与Cd+NaHS处理的均差异显著。Cd胁迫下添加外源Ca2+处理的根系线粒体Δψ与Cd+NaHS处理的水平相当(图4-B)。
与对照相比,在Cd处理下,随着MPTP 开放程度的增大,Δψm的降低,Cyt c/a 也显著降低。与单独Cd胁迫相比,Cd+ NaHS和Cd+CaCl2处理显著提高了根系线粒体Cyt c/a。而Cd胁迫下添加NaHS的同时,添加钙离子相关抑制剂均显著抑制了H2S的缓解作用,其中Cd+ NaHS +CPZ处理的抑制效果最明显,Cyt c/a比Cd+NaHS处理降低了29.9%(图4-C)。
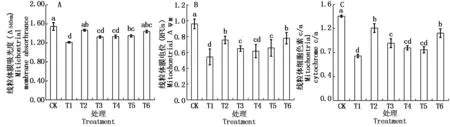
图4 Ca2+和H2S诱导Cd胁迫下平邑甜茶幼苗根系线粒体膜透性、膜电位和细胞色素c/a的影响Fig.4 Effect of Ca2+ and hydrogen sulfide on the membrane absorbance, Δψ,Cyt c/a of mitochondrial in roots of Malus hupehensis Rehd. seedlings under Cd stress
3 讨论
已有研究发现,在较严重的胁迫条件下,线粒体是响应逆境信号的初始位点,通过启动一系列代谢过程来增强植物的环境适应[17]。镉胁迫下连接线粒体内外膜的MPTP过度开放,使线粒体膜MPT不断增大,导致呼吸链解偶联,Δψm下降甚至消失,松散得结合在线粒体内膜上的Cyt c穿过线粒体膜进入细胞质。Cyt c是线粒体电子传递的组成成分,它的脱落破坏线粒体抗氧化防御体系,导致ROS和MDA等物质的大量产生,诱导细胞死亡,使细胞正常生理代谢受阻,幼苗生长受到明显抑制[3]。
近年来越来越多的证据表明,H2S可以通过调节抗氧化酶活性和非酶类物质含量,清除细胞内过量的ROS,缓解逆境对细胞的氧化损伤[6-7,11]。但H2S对植物遭遇镉胁迫后的线粒体功能是否具有调节作用未见报道。本试验中在镉胁迫下添加NaHS处理可显著减少平邑甜茶幼苗根系细胞的死亡数量,减轻Cd对幼苗生长的抑制。这可能是由于外源H2S可有效提高线粒体的完整性,抑制线粒体内ROS的产生;同时H2S激活了抗氧化酶系统,SOD、CAT、POD活性增强,使产生的ROS能很快被还原,减轻Cd胁迫引起的膜脂过氧化对线粒体膜的伤害和对电子传递链的阻抑,保护了线粒体功能,缓解伤害。
植物在受到非正常刺激时,可以通过细胞质膜上Ca2+通道的开放或细胞内钙库的Ca2+释放提高细胞质基质中的Ca2+水平,从而形成Ca2+信号,引发系列生理生化代谢[18-19]。已有研究表明,一定浓度的外源Ca2+处理可以有效提高大蒜对镉胁迫的抗性及葡萄对光氧化胁迫的适应[20-21]。本研究中,Cd胁迫下添加外源Ca2+处理减小了根系MPTP 的开放程度,显著提高了根系线粒体的Δψm和抗氧化酶活性,使ROS的水平显著低于单独Cd处理,膜脂过氧化保持在较低的水平,最终缓解了Cd胁迫对幼苗生长的抑制。但添加NaHS的同时添加钙离子专一螯合剂EGTA,钙离子通道阻断剂LaCl3或钙调素拮抗剂CPZ后,外源H2S的作用被解除。这是因为逆境下胞外Ca2+通过Ca2+通道的激活进入胞内,引起胞质Ca2+浓度瞬间增加,Ca信号与其靶蛋白CaM结合成Ca2+-CaM启动复杂的信号转导体系,导致植物做出相应的应答反应[22]。本试验中,当Ca2+被EGTA螯合、Ca2+通道被LaCl3抑制及Ca2+-CaM的生成被CPZ抑制时,Ca2+的信号传导被阻断,使Ca2+对植物体的生理功能被解除。这表明,Ca2+在H2S诱导的保护酶活性增强,线粒体功能稳定,活性氧清除能力提高,膜脂过氧化降低,从而缓解镉胁迫伤害的过程中起重要作用,而H2S的作用可能依赖于Ca2+。植物响应逆境的信号传导是复杂的网络系统,可能存在多种交叉对话,在某个地方汇集启动Ca2+依赖或非依赖性的信号传导途径,因此,明确H2S和Ca2+的作用需要进一步的试验支持。
参考文献:
[1] Azevedo R A,Gratao P L,Monteiro C C,et al. What is new in the research on cadmium-induced stress in plants?[J]. Food and Energy Security,2012,1(2):133-140.
[2] Bartoli C G,Gomez F,Martinez D E,et al. Mitochondria are the main target for oxidative damage in leaves of wheat (TriticumaestivumL.)[J]. Journal of Experimental Botany,2004,55(43):1663-1669.
[3] 姜倩倩. 镉胁迫下平邑甜茶根系细胞死亡及其中介因素研究[D]. 泰安:山东农业大学,2011.
[4] Li Z G,Gong M,Liu P. Hydrogen sulfide is a mediator in H2O2-induced seed germination inJatrophaCurcas[J]. Acta Physiologiae Plantarum,2012,34(6):2207-2213.
[5] Garc′a-Mata C,Lamattina L.Hydrogen sulphide,a novel gasotransmitter involved in guard cell signaling[J]. The New Phytol,2010,188(4):977-984.
[6] 周超凡,吴帼秀,李 婷,等. 外源 H2S 对低温下日光温室黄瓜光合作用及抗氧化系统的影响[J]. 园艺学报,2016,43(3):462-472.
[7] Zhang H,Hu S L,Zhang Z J,et al. Hydrogen sulfide acts as a regulator of flower senescence in plants[J]. Postharvest Biology and Technology,2011,60(3):251-257.
[8] Christou A,Manganaris G A,Papadopoulos I,et al. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defence pathways[J]. Journal of Experimental Botany,2013,64(7):1953-1966.
[9] Chen J,Wang W H,Wu F H,et al. Hydrogen sulfide alleviates aluminum toxicity in barley seedlings[J]. Plant and Soil,2013,362(1/2):301-318.
[10] Li Z G,Ding X J,Du P F. Hydrogen sulfide donor sodium hydrosulfide-improved heat tolerance in maize and involvement of proline[J]. Journal of Plant Physiology,2013,170(8):741-747.
[11] Cui W T,Chen H P,Zhu K K,et al. Cadmium-induced hydrogen sulfide synthesis is involved in cadmium tolerance inMedicagosativaby reestablishment of reduced (homo) glutathione and reactive oxygen species homeostases[J]. PLoS One,2014,9(10):e109669.
[12] Fang H H,Jing T,Liu Z Q,et al. Hydrogen sulfide interacts with calcium signaling to enhance the chromium tolerance inSetariaitalica[J]. Cell Calcium,2014,56(6):472-481.
[13] Li Z G,Gong M,Xie H,et al. Hydrogen sulfide donor Sodium hydrosulfide-induced heat tolerance in tobacco (NicotianatabacumL.) suspension cultured cells and involvement of Ca2+and calmodulin[J]. Plant Science,2012,185(9):185-189.
[14] 魏国芹,曹 辉,孙玉刚,等. 硫化氢对淹水平邑甜茶根系形态构型,叶片活性氧和光合特性的影响[J]. 应用生态学报,2017,28(10):3267-3273.
[15] Steffens B,Sauter M. Epidermal cell death in rice is regulated by ethylene,gibberellin,and abscisic acid[J]. Plant Physiology,2005,139(2):713-721.
[16] 赵世杰,史国安,董新纯. 植物生理学实验指导[M]. 北京:中国农业科技出版社,1998.
[17] 苏 宏,李丽杰,马怀宇,等. 变温条件下钙对山定子根系线粒体功能的影响[J]. 中国农业科学,2014,47(19):3866-3873.
[18] 张 召,梁元存,王 利,等. 钙对酸化处理平邑甜茶根系抗氧化酶活性及线粒体功能的影响[J]. 林业科学,2012,48(8):87-93.
[19] 应叶青,杜 旭,华姜琴,等. 干旱胁迫下毛竹根尖Ca2+分布及外源Ca2+作用机制[J]. 林业科学,2013,49(4):141-146.
[20] 李 贺,连海峰,刘世琦,等. 镉胁迫对大蒜苗生理特性的影响及施钙的缓解效应[J]. 应用生态学报,2015,26(4):1193-1198.
[21] 王晓芳,相 昆,孙 岩,等. 叶面肥对“巨峰”葡萄光氧化胁迫的缓解效应[J]. 果树学报,2017,34(3):312-320.
[22] Yang T B,Poovaiah B W. Calcium/calmodulin-mediated signal network in plants[J]. Trends in Plant Science,2003,8(10):505-512.
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
中华实用诊断与治疗杂志(2022年1期)2022-08-31
舰船科学技术(2022年11期)2022-07-15
海洋通报(2021年5期)2021-12-21
农业资源与环境学报(2021年5期)2021-10-06
家庭影院技术(2021年7期)2021-08-14
海洋通报(2021年1期)2021-07-23
西北农林科技大学学报(自然科学版)(2021年2期)2021-03-09
中国果业信息(2019年10期)2019-01-05
医药前沿(2019年35期)2019-01-05
