
基于DCQGA的PID参数自整定
2018-04-26 05:45:24江鸿潮白国振
石油化工自动化 2018年2期
江鸿潮,白国振
(上海理工大学 机械工程学院,上海 200093)
PID控制器参数优化的关键问题,是如何获得偏差的比例、积分、微分三者之间的最佳组合。PID参数整定的方法很多,比如常规的Ziegler-Nichol法[1]和Cohen-Coon法等,然而常规方法在实际工程应用中需要依赖很强的实践经验,且不易得到理想的结果。近年来,智能算法在PID控制中的成功应用和发展为PID参数整定提供了新的路径,如人工免疫整定[2]、蚁群算法整定[3]、鱼群算法整定[4]、佳点遗传算法集整定[5]等,这些智能PID整定方法,大幅提高了PID控制系统的控制性能。
在用于PID参数整定的各智能算法中,遗传算法[6]GA(genetic algorithm)便是其中最常用的算法之一,由于GA仅用目标函数在概率准则引导下进行全局自适应搜索,能够处理传统优化方法难以解决的复杂问题,具有极高鲁棒性和广泛适用性,因而在各个优化领域得到了广泛应用.但是GA在实际应用中存在迭代次数多、收敛速度慢、易陷入局部极值和过早收敛等缺陷和不足。
为解决这些问题,Narayanan[7]和Han Kuk-Hyun[8]等将量子计算中的量子比特(qubit)、量子态叠加和量子干涉等概念引入遗传算法,提出了一种新的遗传算法——量子遗传算法(QGA),该算法使用量子比特进行基因编码,用量子门进行染色体更新,有效地解决了GA中存在的染色体丢失和过早收敛的问题[9]。但QGA的旋转角大小和方向的确定过于复杂,对于多参数函数优化问题容易出现早熟现象[10]。标准QGA由于需要反复查表,计算量大,非常耗时,不便于工程实际应用。
针对以上问题,本文采用一种基于实数编码的双链量子遗传算法[11-12](DCQGA)来整定PID参数。该算法在基本量子遗传算法的基础上进行改进,对转角方向的确定,给出了一种全新的但简单实用的确定方法;对转角迭代步长的确定,构造出1个包含目标函数的梯度信息的转角函数;此外,把量子比特的2个概率幅值都看做基因位,使得每条染色体带有2条基因链,加快了搜索速度。通过与GA和QGA的比较,结果验证了DCQGA在PID参数优化中的优越性。
1 量子遗传算法简述[13]
量子遗传算法是将量子计算原理中的量子态、量子态叠加、幺正变换等引入普通遗传算法而得到的一种新的优化算法,其既具有普通遗传算法优胜劣汰不断进化的优势,又具有量子计算中强大的并行搜索优势。QGA仅用1条染色体就可以同时表示多个状态,而 GA的1条染色体只能表示一种状态,这使得QGA较传统GA具有更好的种群多样性和更高的计算并行性。相较于GA的选择、交叉和变异操作,QGA在染色体更新时采用了量子旋转门操作,大幅地提高了算法的收敛速度。
1.1 量子比特编码
量子比特编码不同于遗传算法中的二进制编码,它的染色体基因位不再用确定的值表示,而是用量子位表示,量子位的值通过随机的方式取得,是一种概率表示形式。遗传算法采用经典比特来表征不同的信息,每个基因位要么是“0”,要么是“1”,是确定的值,它的编码方式属于确定论的范畴;而QGA采用的是量子比特|φ>来表征信息,其中:
|φ>=α|0>+β|1>
(1)
|α|2+|β|2=1
(2)
它的基因位不再是确定的数值,而是量子位|φ>,它是1个不确定的值,它的编码方式属于概率论的范畴,只有人为地去观测时才能得到它的确定值(坍缩为基本态)。就像薛定谔的猫一样,在没有打开箱子时,它是处于既死又活的叠加态,只有打开箱子才能确定猫是处于死态还是活态。
同理,量子比特也有2个基本态|0>态和|1>态,未被观察时量子比特处于两种状态的线性叠加态,一旦有人为的观察时就会坍缩到基本态|0>态或|1>态。其中|α|2给出了量子比特被观察时坍缩到|0>态的概率,|β|2给出了量子比特被观察时坍缩到|1>态的概率。这样,用量子比特编码的染色体不再表示一种单一的状态,而是表示多个状态的叠加。如用n个量子位就可以表示2n个状态。采用多量子比特编码m个参数的基因如下[14]:
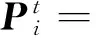
(3)

采用量子比特编码,1条染色体就可以同时表达多个态的叠加,使得QGA比普通遗传算法拥有更好的多样性特征。这样QGA相比传统的GA可以大量减少种群规模,大量地减少计算时间。同时QGA通用性好,且实现简单。
1.2 量子旋转门更新
QGA中染色体的更新是通过量子门来实现的,而量子旋转门是用得最多的一种量子门,它通过对量子比特实施幺正变换来控制量子态的演化和传递,进而实现种群的进化,其更新过程如下[15]:
(4)
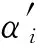
2 基于DCQGA的PID参数整定
PID参数整定是多目标优化问题。首先给出连续优化问题的一般描述[16]:
约束条件为n维空间,其有界闭集Ω中每个点都看成优化问题的近似解,为反映这些近似解的优劣程度,可定义适应度函数:
fit(x)=Cmax-f(x)
(6)
式中:Cmax——给定的1个适度值或目前为止的最大值。
2.1 实数双链编码方案
本文采用的DCQGA中,对编码方案进行了改进[16]:
(7)
式中:Pi——每一代中第i个个体的染色体;θij=2π×rand;rand为[0,1]之间随机数;i=1,2,…,m;j=1,2,…,n;m——种群规模;n——量子位数。
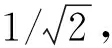
(8)
式中:Pic——余弦解;Pis——正弦解。
2.2 解空间变换
由2.1节的分析可知,无论是QGA还是DCQGA,通过染色体解码得到的值与实际优化问题的解空间都不一致,所以需要进行解空间变换。变换公式如下:
(9)
(10)
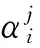
2.3 改进的种群进化策略
2.1节给出了对QGA编码方式的改进,下面对种群进化策略进行改进,DCQGA的种群进化策略同样运用量子旋转门,但在转角方向和大小的确定上都给出了改进措施。
2.3.1量子旋转门的转角方向的确定
DCQGA的量子旋转门为
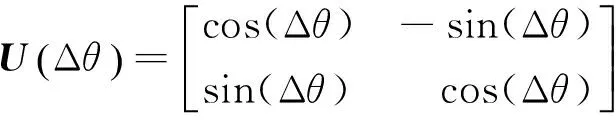
(11)
该量子旋转门不改变量子位的长度,只改变量子位的相位。其更新过程为
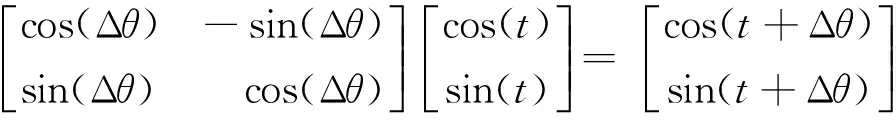
(12)
转角Δθ的大小和方向直接关系到算法的速度和效率。确定转角Δθ的方向,QGA的做法是构造1个查询表[14],虽然直观但异常繁琐,而且需要频繁地查表,使得QGA的效率偏低。为了简化,DCQGA给出了以下确定转角方向的策略: 令
(13)
当A≠0时,Δθ方向为-sgn(A);当A=0时,方向取正负均可。其中,α0和β0是当前搜索到的全局最优解中某量子位的概率幅,α1和β1是当前解中相应量子位的概率幅。
证明: 记量子位[α0,β0]T和[α1,β1]T在单位圆中的幅角分别为θ0和θ1,则
(14)
当A≠0时,若0<|θ1-θ0|<π,sgn(Δθ)=-sgn(θ1-θ0)=-sgn(sin(θ1-θ0))=-sgn(A);若π<|θ1-θ0|<2π,则sgn(Δθ)=sgn(θ1-θ0) =-sgn(sin(θ1-θ0))= -sgn(A)。当A=0时,由sin(θ1-θ0)=0,得θ1=θ0或|θ1-θ0|=π,此时,正、反向旋转没有区别,效果相同,故sgn(Δθ)取正负均可。(证毕)
2.3.2量子旋转门转角大小的确定
量子旋转门转角大小的确定一般是构造1个查询表,如文献[14],旋转幅度大多是根据经验给出,如文献[7]给出了开区间(0.005π,0.100π)的范围,但是并没有给出选择的具体依据。文献[17]给出了一种转角的迭代步长单调下降的调整策略,是一种以进化代数为自变量的负指数函数的自适应调整策略。现有文献大多没有考虑种群中各染色体之间的差异,也没有充分利用目标函数的变化趋势。本文的改进策略是: 把目标函数在搜索点(单个染色体)处的变化趋势(梯度)加入到转角步长函数中,让转角步长的大小随着目标函数变化率增大而减小,反之亦然。这样,既可以加快搜索速度,又不至于越过全局最优解。考虑可微目标函数的变化率,利用梯度定义转角步长函数为
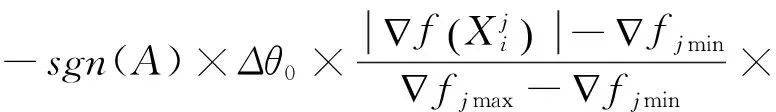
exp(-gen/maxgen)
(15)
式中:Δθ0——迭代初值;gen——当前代数,maxgen为终止代数;适应度函数f(X)在处的梯度;fjmax,fjmin分别定义为
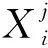
(18)
式中:Xp,Xc——父代和子代染色体。
2.4 变异处理
除了对编码方式以及转角方向和大小进行改进外,DCQGA在基本QGA基础上加入了变异处理。首先依据实际确定的变异概率随机选择需要变异的若干染色体,然后再用量子非门对随机选中的量子位施加变换,使该量子位的4个概率幅互换。这样可使2条基因链同时得到变异。
比如,假设某量子位幅角为θ,则变异后幅角变为π/2-θ,也就是说幅角正向旋转了π/2-2θ。实际上该种变异是对量子位幅角的一种旋转: 该旋转不与当前最佳染色体比较,一律正向旋转,有助于增加种群的多样性,降低早熟的概率。
2.5 DCQGA的算法流程
实现上述实数双链编码梯度QGA的步骤如图1所示。
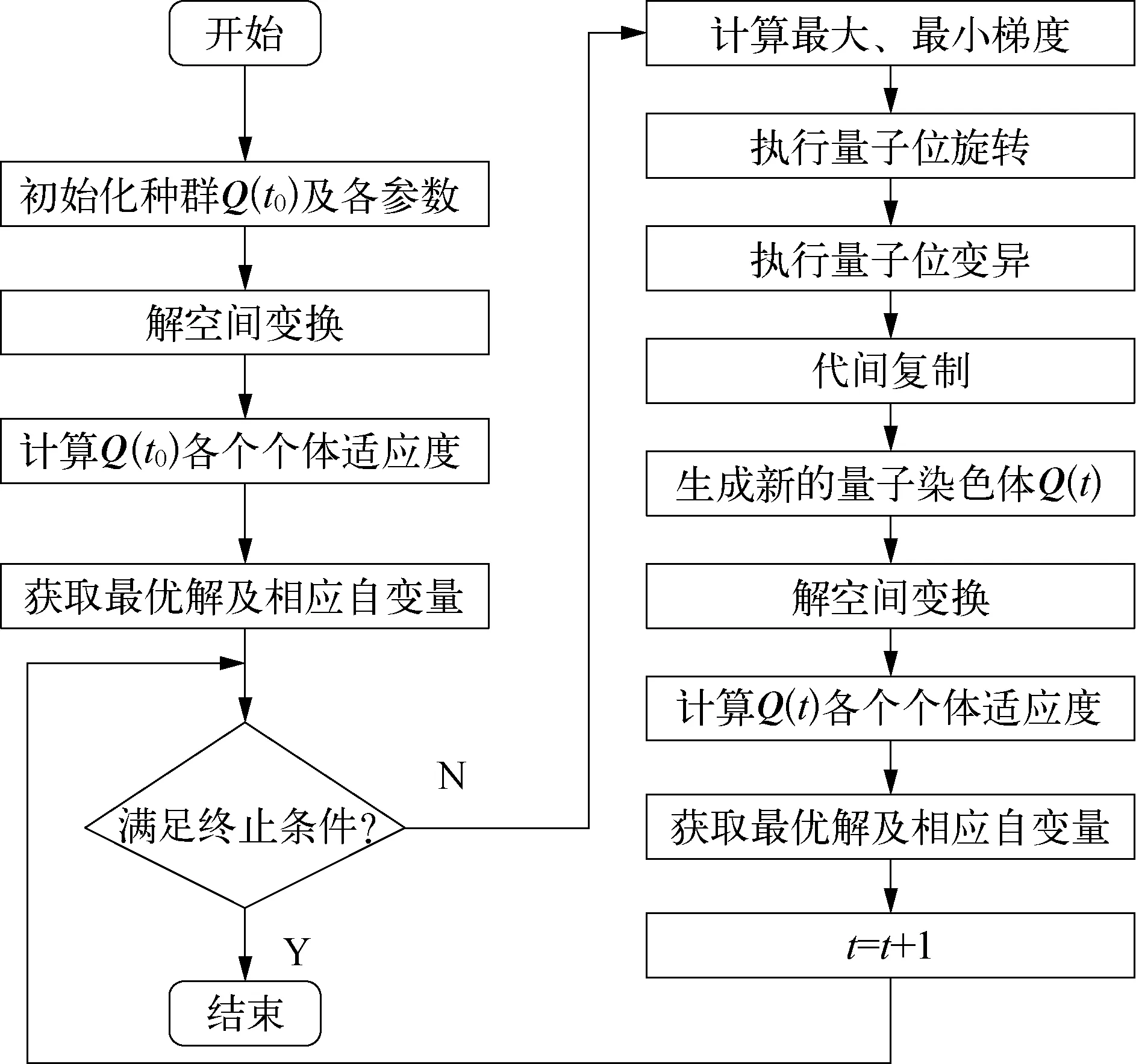
图1 DCQGA流程示意
1) 种群初始化。按式(7)产生m条染色体(2m条基因链)组成初始种群Q(t0);设定初始转角步长θ0、变异概率为pm。
2) 解空间变换。将每条染色体代表的近似解,由单位空间In=[-1,1]n映射到连续优化问题式(5)的解空间Ω。
3) 计算初始种群Q(t0)的适应度。按式(6)计算染色体的适应度fit。
4) 获取最优解及相应自变量。记当代最优解为bestX,对应染色体为bestChrome,到目前为止的最优解为BsX,对应染色体为BsChrome。若fit(bestX)>fit(BsX),则BsChrome=bestChrome。
5) 判断是否满足终止条件。终止条件的设定一般有两种: 适应度值达到要求时终止程序;达到指定代数时终止程序,本文采用后者。当满足终止条件时,6)之后的步骤不执行,而不满足终止条件时,进入下一步的循环体。
6) 计算最大最小梯度。根据式(16)和式(17)计算最大最小梯度fjmax,fjmin。
7) 执行量子位旋转。根据式(15)执行量子位旋转,旋转后得到1个关于转角步长Δθ的矩阵。
8) 执行量子位变异。依据变异概率对要变异的量子位实行变异操作: Δθij=0.5π-θij。
9) 代间复制。因为要用到相邻两代的一阶差分替代梯度,所以要先把本代染色体复制上一代,即令oldchrome=chrome。
10) 生成新的量子染色体。由步骤8)得到更新后的Δθij,chrome=cos(Δθij)或chrome=sin(Δθij)得到新的量子染色体。
11) 得到新的染色体后,反复执行步骤2)~5),直到t达到指定代数时结束。
4 仿真验证
为了验证DCQGA对PID控制器参数优化的有效性,选择如下被控对象[18]:
(21)
令输入为一阶跃信号,采样时间取为1 ms。为了获得满意的过渡过程动态特性,采用误差绝对值时间积分性能指标作为参数选择的最小目标函数。另外,把控制量的平方也加入到目标函数中,这样可以避免控制能量过大,如下式:
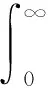
(22)
式中:e(t)——系统误差;u(t)——控制器输出;tu——上升时间;ω1,ω2,ω3——权值。
为避免超调,采用惩罚机制,即一旦产生超调,将超调量也加入目标函数中,即:
ife(t)<0
(23)
式中:ω4——权值,且ω4≥ω1。
在用PID控制器对该模型进行控制时,为优化PID控制器参数,各算法参数选择如下:kP的取值范围为[0,20];kI,kD的取值范围为[0,1];取ω1=0.999,ω2=0.001,ω3=2.0,ω4=100;限定代数为160代。GA,QGA和DCQGA三种算法针对以上的参数取值均一致。
另外,DCQGA的种群规模取为10,量子位数为3,转角步长为0.002π,变异概率为0.05;GA的交叉概率取为0.9,变异概率为0.033;而QGA的种群规模取为40。
5 结果分析
经过160代的优化,三种算法得到的最优结果见表1所列。
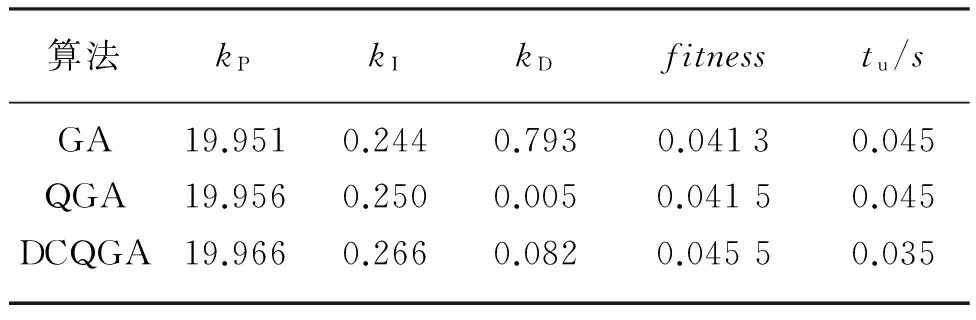
表1 GA,QGA,DCQGA算法结果比较
表1中,fitness表示最优适应度值,tu表示对应算法的上升时间。由表1可知DCQGA的bestfit最大,同时从适应度函数变化曲线图2中也可以看出DCQGA的曲线明显高于另外2条,表明DCQGA可以更快地跳出局部最优解,获得更好的全局最优解,优化结果更准确。
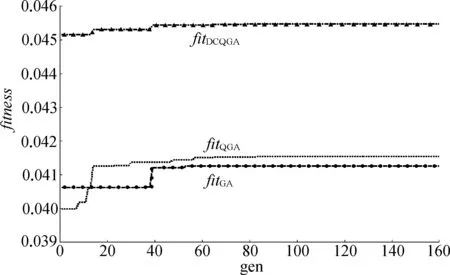
图2 适应度函数变化曲线
图3为DCQGA与QGA和GA优化的PID控制器的阶跃响应的比较。从图3和表1可以看出DCQGA得到的曲线上升时间最快,上升时间tu最小,说明DCQGA的响应速度最好。
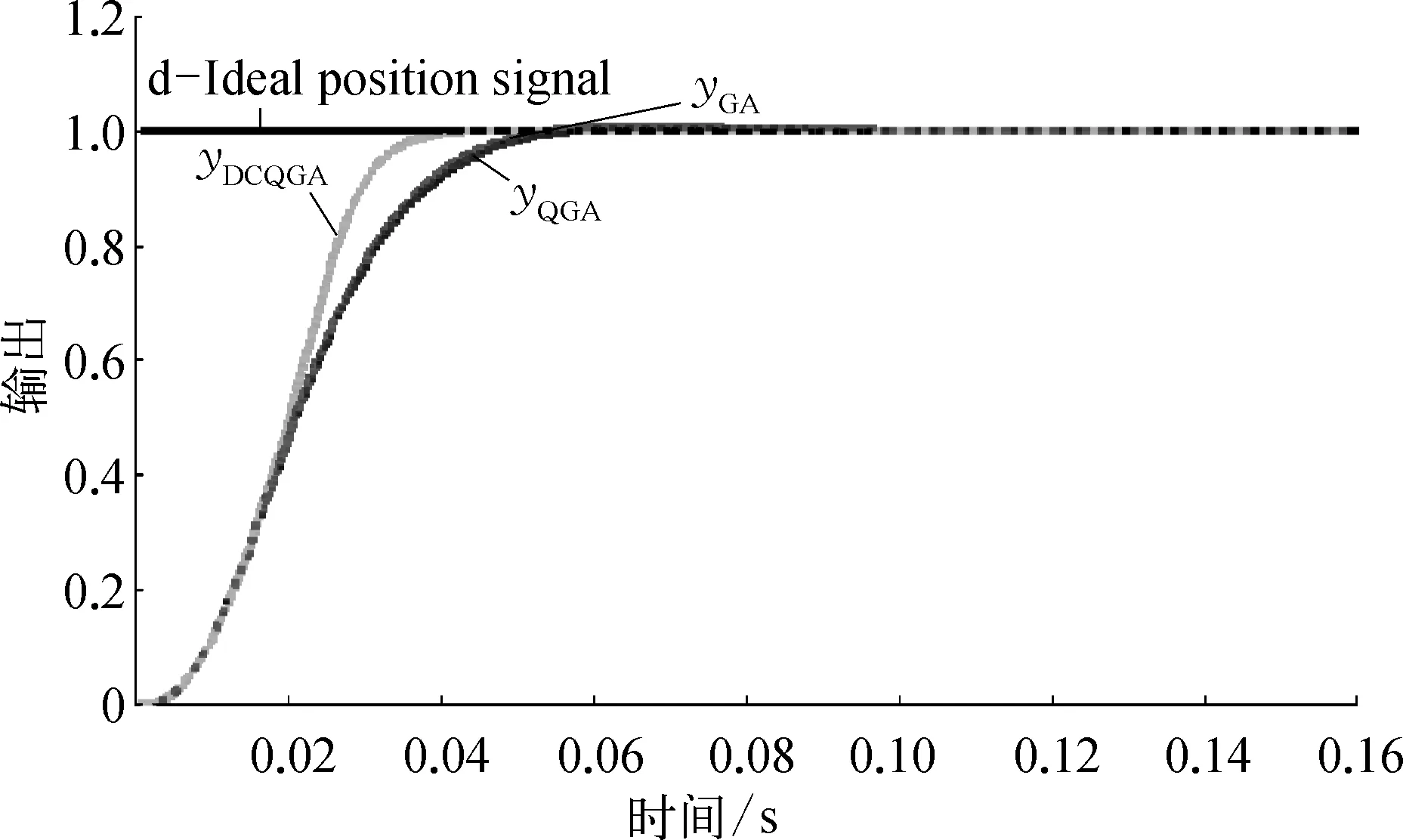
图3 PID控制器的阶跃响应示意
6 结 论
本文提出一种基于实数编码的双链量子遗传算法的PID参数自整定方法,该方法用量子比特构成染色体,用实数对量子比特进行编码;相比QGA,该方法将每个量子位看做上下2个并列的基因,每条染色体包含2条并列的基因链,每条基因链代表1个最优解。通过与普通遗传算法和标准量子遗传算法整定效果的对比,结果验证了DCQGA可以更快地跳出局部最优解从而获得更好的全局最优解,同时具有更快的响应速度和更好的多目标寻优能力。
参考文献:
[1] Ziegler J, Nichols N. Optimum Settings for Automatic Controllers[J].Transaction of ASME, 1942(64): 759-768.
[2] 刘屿,田联房,毛宗源.一种新型人工免疫算法的PID自整定研究[J].计算机应用研究,2007,24(04): 84-87.
[3] 陈书谦,张丽虹.蚁群算法在 PID 控制器参数优化中的应用研究[J].计算机应用研究,2011, 28(01): 177-181.
[4] 余丽莹,焦嵩鸣.基于鱼群算法的PID优化[J].计算机仿真,2014,31(03): 155-158.
[5] 彭勇,施宁,林浒.佳点遗传算法集及其在 PID控制中的应用[J].计算机应用研究,2009,26(02): 524-526.
[6] 周明,孙树栋.遗传算法原理及应用[M].北京: 国防工业出版社,1999
[7] Narayanan A, Moore M.Quantum-inspired Genetic Algorithms[C]//Proceedings of IEEE International Conference on Evolutionary Computation. Nagoya, Japan, 1996: 61-66.
[8] Han Kuk-Hyun, Kim Jong-Hwan.Quantum-inspired Evolutionary Algorithm for a Class of Combinatorial Optimization[J].IEEE Trans Evolutionary Computation, 2002, 6(06): 580-593.
[9] Zhang G X, Gu Y J, Hu L, et al. A Novel Genetic Algorithm and Its Application to Digital Filter Design[C]//Proc on IEEE Intelligent Transportation System. New Jersey: IEEE Press, 2003, 2: 1600-1605.
[10] 商允伟,裘聿皇.适应值共享对遗传算法选择概率的影响分析[J].控制与决策,2003,18(06): 708-711.
[11] Li P C,Li S Y.Quantum-inspired Evolutionary Algorithm for Continuous Spaces Optimization[J].Chinese Journal of Electronics, 2008, 17(01): 100-104.
[12] 李士勇,李盼池.基于实数编码和目标函数梯度的量子遗传算法[J].哈尔滨工业大学学报,2006(08): 1216-1218.
[13] 张兴华,朱筱蓉,林锦国.基于量子遗传算法的PID控制器参数自整定[J].计算机工程与应用,2007(21): 218-220.
[14] 郁磊,史峰.Matlab智能算法30个案例分析[M].北京: 北京航空航天大学出版社,2015.
[15] 曾成,赵锡均.改进量子遗传算法在PID参数整定中的应用[J].电力自动化设备,2009(10): 125-127,139.
[16] 李士勇,李盼池.量子计算与量子优化算法[M].哈尔滨: 哈尔滨工业大学出版社,2009.
[17] Zhang G X,Li N,Jin W D.A Novel Quantum Genetic Algorithm and Its Application[J].ACTA Electronic Sinica, 2004, 32(03): 476-479.
[18] 刘金琨.先进PID控制Matlab仿真[M].北京: 电子工业出版社,2011: 25-26.
猜你喜欢
中学生数理化·七年级数学人教版(2021年9期)2021-11-20 06:11:52
科学之谜(2019年3期)2019-03-28 10:29:44
科学之谜(2018年8期)2018-09-29 11:06:46
海峡姐妹(2017年10期)2017-12-19 12:26:20
三联生活周刊(2017年33期)2017-08-11 04:35:44
解放军健康(2017年5期)2017-08-01 06:27:42
银行家(2017年1期)2017-02-15 20:27:20
知识经济·中国直销(2016年2期)2016-11-07 09:33:42
恋爱婚姻家庭·养生版(2016年9期)2016-09-07 11:25:01
中央民族大学学报(自然科学版)(2015年2期)2015-06-09 08:45:16
