
遮阴和干旱对荩草生理代谢及抗性系统影响的协同作用
2018-04-19 01:23:53张小晶刘金平游明鸿郭碧花
生态学报 2018年5期
孙 帅,张小晶,刘金平,*,游明鸿,郭碧花
1 西华师范大学生命科学学院,南充 637009 2 四川省草原科学研究院,成都 611731
紫色土丘陵区属典型的亚热带季风性湿润气候,该区土壤发育时间短、土质松散、土层薄、土壤贫瘠、可溶性盐含量较高,受地形地势、日照、温度、降雨等自然因子影响,是侵蚀度仅次于黄土高原的极易水土流失的生态脆弱区[1]。受过度耕作、工业发展、开山修路、地产开发等人类活动影响,原有植被频遭破坏,形成了大量的裸露边坡,边坡土壤与基岩粘结性能差、抗蚀能力弱,崩解风化速度快,水土流失极为严重,易由面蚀发展为沟蚀[2]。为了恢复植被、提升边坡景观效果和水土保持能力,常通过工程技术、客土喷播技术、挂网技术,建植固土护坡草坪,进行边坡治理与生态重建。引进的暖季型草坪草枯黄期长达100d左右,为实现四季常绿的造景效果,常采用冷季型草坪草建植单一或混播边坡草坪。由于夏季高于35℃的酷热天气频现,8月绝对高温常超过40℃[3],冷季型草坪草夏眠严重,且水热同期致高温高湿常引发严重病害,使引进草种面临巨大生存风险。加之边坡草坪养护成本高、难度大,仅2—5年草坪退化殆尽[4],难以起到植被恢复和景观构建的预期效果,造成人力物力浪费的同时,引进草种还会带来潜在的生态危机。目前建植集绿化、观赏和固土功能一体的边坡草坪时,受建坪目的、立地条件和养护技术限制,几无合适引进草种可供选择。所以,筛选、培育具有极强适应能力和抗逆性的本土固土护坡草坪植物,是急需解决的实际问题。开发利用丰富的野生乡土草坪植物资源,是该区绿化、美化事业和生态治理的可持续发展的必由之路。
荩草[Arthraxonhispidus(Thunb.)Makino.]为紫色土丘陵区常见的禾本科荩草属一年或多年生草本乡土植物,别名竹叶草、绿竹、马耳草和中亚荩草等,有药用、饲用、染用和绿化价值。荩草野生资源丰富、生态型多、生长期长、生长速度快和再生性好,有广泛生态适应[3],具株秆细而硬、多分枝、基部倾斜、着地后节易生根等特点。有极强的繁殖能力,通过种子自播和无性拓展常入侵退化边坡草坪,逐渐成为优势种。有极强的耐贫瘠能力,甚至在大于70°断面坡上形成局部单一种群,在固土护坡草坪建植及生物修复方面有巨大开发潜力[5]。目前仅对荩草适应性和耐阴性进行了浅显分析[6-7],开展荩草生态学特点及开发利用途径研究,对植物资源保护与利用、生态环境治理、绿化草种选育及中草药持续性开发具有重要意义。
因立地条件和乔灌草结合的配置模式,边坡草坪常受遮阴和干旱胁迫。遮阴和干旱使光和水分资源分配失衡,草坪植物的生理代谢及抗性系统对立地条件的响应能力,必将影响植株的生长发育、能量积累和扩繁能力,决定着草坪外观质量、景观价值及寿命长短。本研究将荩草幼苗放置于5种遮阴度和4种干旱度交互形成的20个处理中,研究光照和水分变化对荩草生理代谢和抗性系统的影响,分析荩草的抗逆潜力和生态学特点,以期为野生荩草以边坡草坪开发利用提供依据。拟回答以下科学问题:(1)荩草光合生理对遮阴和干旱胁迫如何响应?(2)荩草糖代谢、氮代谢和呼吸作用对遮阴和干旱胁迫的如何应对?(3)荩草抗氧化酶系统对遮阴和干旱胁迫如何启动?(4)荩草细胞膜伤害程度对遮阴和干旱胁迫如何变化?
1 材料和方法
1.1 试验材料
以四川省南充市顺庆区的荩草(Arthraxonhispidus)为材料。在野生荩草种群内,选取当年生健壮匍匐茎,剪取基部茎段,去除次生根,选含2节、节间4cm、长约6cm为插穗,置阴凉处保湿备用。
以营养土∶紫色土1∶3为基质,除杂、晾晒、消毒、拌匀后,每盆2.5kg装于口径20cm,高28cm的花盆中,浇水沉降稳定后,细平整表层进行扦插。共60盆,每盆10插穗,在室温约27℃、相对湿度约30%的通风条件下养护管理。
1.2 试验设计
养护1月后,以生长健壮、长势一致、2片真叶为标准,每盆5株定苗。以自然光为基础,借助有效辐射计(GLZ-C),用遮阴网围成六面体遮阴罩,设置0、10%、30%、50%、70%等5个遮阴梯度;在自然蒸发下,用土壤含水量测定仪(TDR300)测浇水515mL/盆(饱和持水量)24、96、192h和288h后,土壤水分分别为饱和含水量的81.75%、65.18%、43.64%和27.88%基础上,设每24、96、192h和288h浇水515mL/盆为水分充足(W0)、轻度干旱(W1)、中度干旱(W2)、重度干旱(W3)4个水分梯度;60盆分为20个遮阴和水肥协同处理组,每组3盆,共15株。置1.5m高钢架,遮阴罩高出盆面50cm培养。处理36d后,进行下列测定。
1.3 测定指标
光合指标测定:用LI-6400光合仪,LED-红蓝光源叶室,自然光源下(约330Lux),每处理随机选取成熟叶片,测净光合速率(Pn),气孔导度(Gs),胞间CO2浓度(Ci)和蒸腾速率(Tr)等光合参数,每点连续采集3个值,3次重复。
代谢指标测定:用“苏州科铭生物有限公司”生产的试剂盒[8],测定可溶性糖(SS)、丙酮酸(PA)含量和硝酸还原酶(NR)活性。
抗性指标测定[9]:超氧化物歧化酶(SOD)活性测定采用氮蓝四哇(NBT)光化还原法,过氧化物酶(POD)活性测定采用愈创木酚法,过氧化氢酶(CAT)活性测定紫外吸收法。
受伤害指标测定[9]:丙二醛(MDA) 采用硫代巴比妥酸(TBA)法,脯氨酸(Pro)采用酸性茚三酮法,可溶性蛋白(SP)含量测定采用考马斯亮蓝G—250染色法,过氧化氢(H2O2)含量采用钼酸法。
1.4 数据分析
用SAS 10.0软件对所有数据进行方差分析和析因分析,并用Duncan法对各参数进行0.05水平显著性检验。
2 结果与分析
2.1 遮阴和干旱对光合指标的影响
遮阴、干旱和协同作用对Pn、Tr、Ci和Gs均有显著影响(P<0.05)(表1)。遮阴下,Pn、Tr和Gs随遮阴度增加表现出先增后减。10%遮阴显著增加Tr和Gs,显著降低Ci。30%遮阴Pn、Tr和Gs均达最大值,Ci达最小值。≥50%遮阴使Pn、Tr和Gs显著降低,Ci达最大值。70%遮阴时Pn、Tr和Gs分别仅为30%的27.18%、36.51%和39.61%,仅为W0-0时的44.07%、78.11%、60.70%。
干旱下,Ci和Tr随干旱度增加逐步下降,Pn和Gs先增后减。W1时Tr显著低于W0,Pn和Gs显著大于W0。W2和W3下Pn、Tr、Ci和Gs显著低于W0和W1。W3下Pn、Tr、Ci和Gs仅为W0时的37.44%、53.25%、13.59和57.64。
遮阴和干旱对光合参数有显著的协同作用(P<0.05)(表1)。≤30%遮阴可缓解干旱使Pn、Tr、Ci、Gs下降的影响,>30%遮阴则加重干旱对光合参数的影响。干旱可减弱或加重遮阴的影响,W1对10%遮阴下的Ci有显著提升作用,W2显著提升10%遮阴下Pn。W1使遮阴下Tr显著下降。干旱越重对遮阴下光合参数的协同作用越大,W3+70%的光合参数仅为W0+0的20%左右。
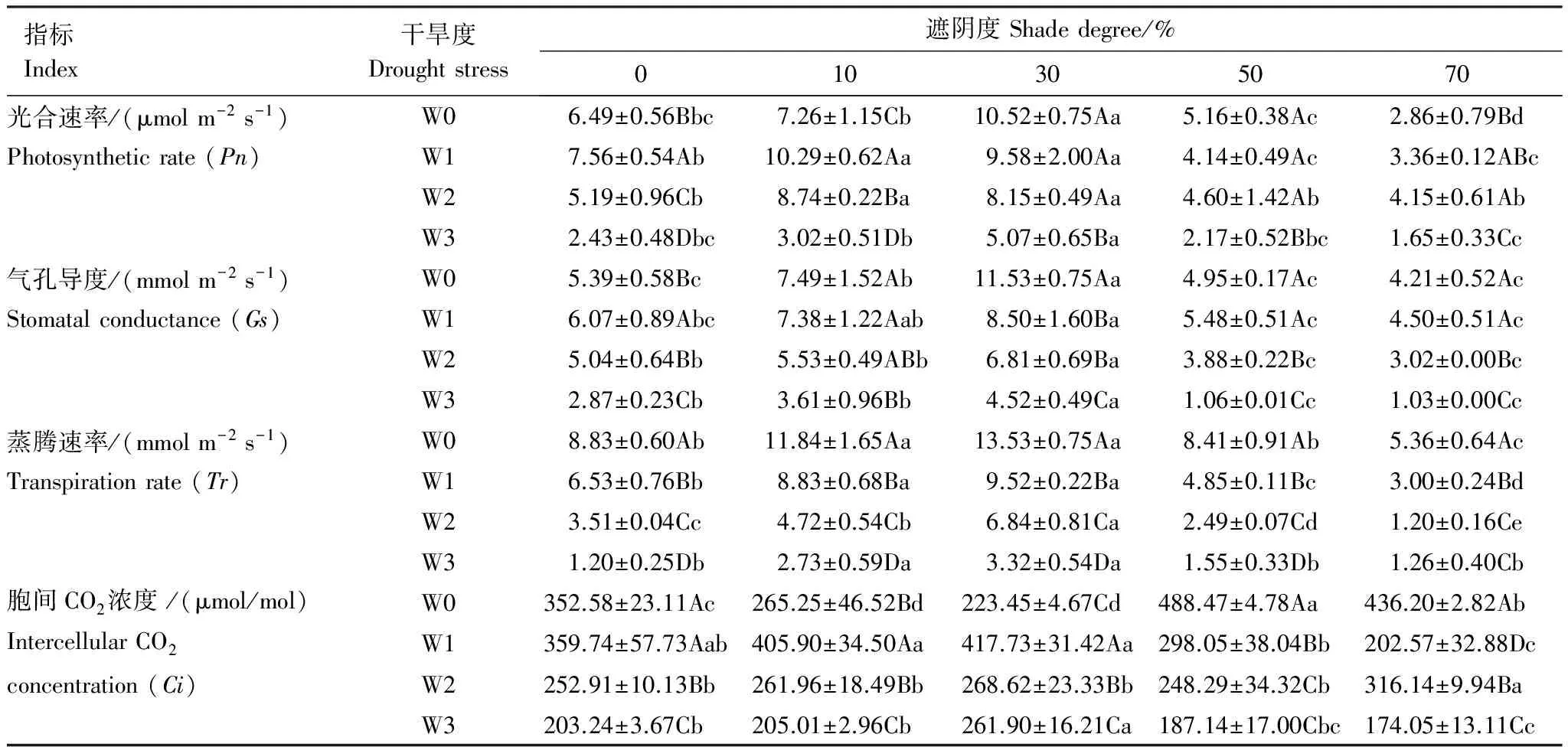
表1 遮阴和干旱对光合指标的影响
数据为平均值±标准误;同一性状的同列不同大写字母表示干旱间差异显著(P<0.05),同行不同小写字母表示遮阴间差异显著(P<0.05);W0:水分充足,Enough moisture;W1:轻度干旱,Mild drought;W2:中度干旱,Moderate drought;W3:重度干旱,Severe drought
方差分析表明,遮阴或干旱对光合参数均有极显著影响(P<0.01)(表2)。由F值可见,遮阴影响顺序为Ci>Tr>Pn>Gs,干旱影响顺序为Ci>Tr>Gs>Pn。遮阴和干旱对光合参数有极显著协同作用(P<0.01),协同作用为Tr>Ci>Pn>Gs。遮阴为影响Pn的主要因子,干旱为影响Gs、Ci和Tr的主要因子,遮阴和干旱对Tr的协同作用最大。
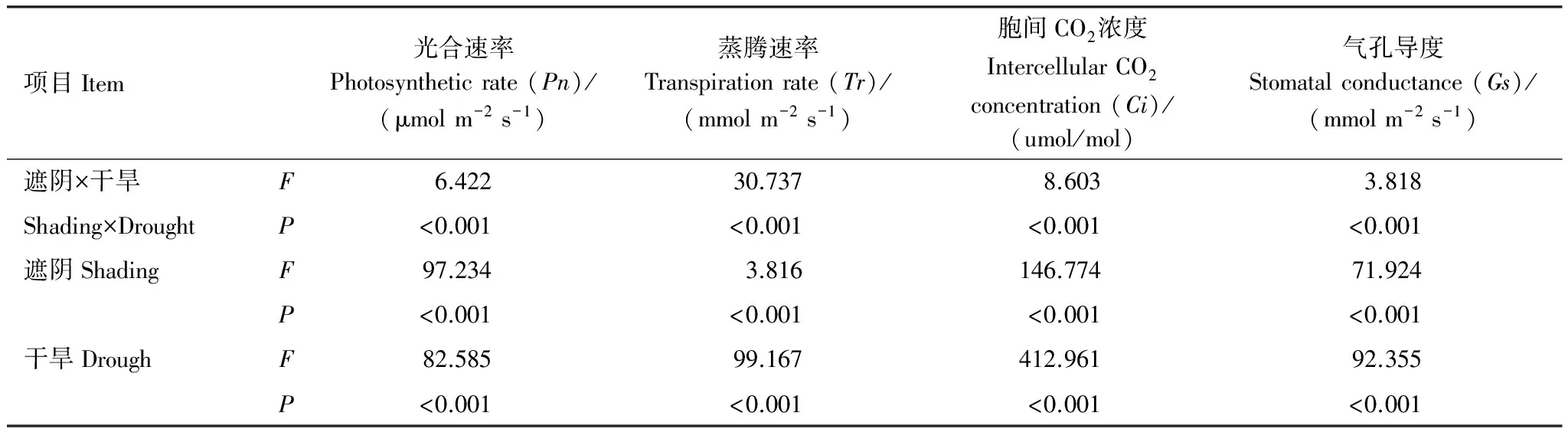
表2 光合指标差异的双因子方差分析
F值表示F检验的显著性,F越大表示越显著,P值表示概率值
2.2 遮阴和干旱对代谢指标的影响
遮阴、干旱和协同处理对SS和PA含量及NR活性有显著影响(P<0.05)(表3)。遮阴下,SS含量随遮阴度增大逐步降低,PA含量逐步增加,NR活性先增后减。10%遮阴使PA和NR显著增大;30%遮阴下NR显著高于其他遮阴;50%遮阴下SS和NR显著降低(P<0.05),PA则达最大值。70%遮阴下SS显著低于其他遮阴,PA无显著变化,NR显著下降,但显著大于CK(W0+0)。
干旱下,SS和PA随干旱加重而增大,NR活性先增后减。W1下SS变化较小,PA和NR则高于W0;W2下SS显著增加达最大值,PA和NR进一步提高;W3下SS无变化,PA显著增加,NR显著降低。
遮阴和干旱对糖代谢(SS)、呼吸作用(PA)和氮代谢(NR)有显著的协同作用(P<0.05)。遮阴可缓解干旱使SS和NR增加的幅度,加重干旱对PA增加的影响。干旱使遮阴下SS减小,使PA和NR增大。
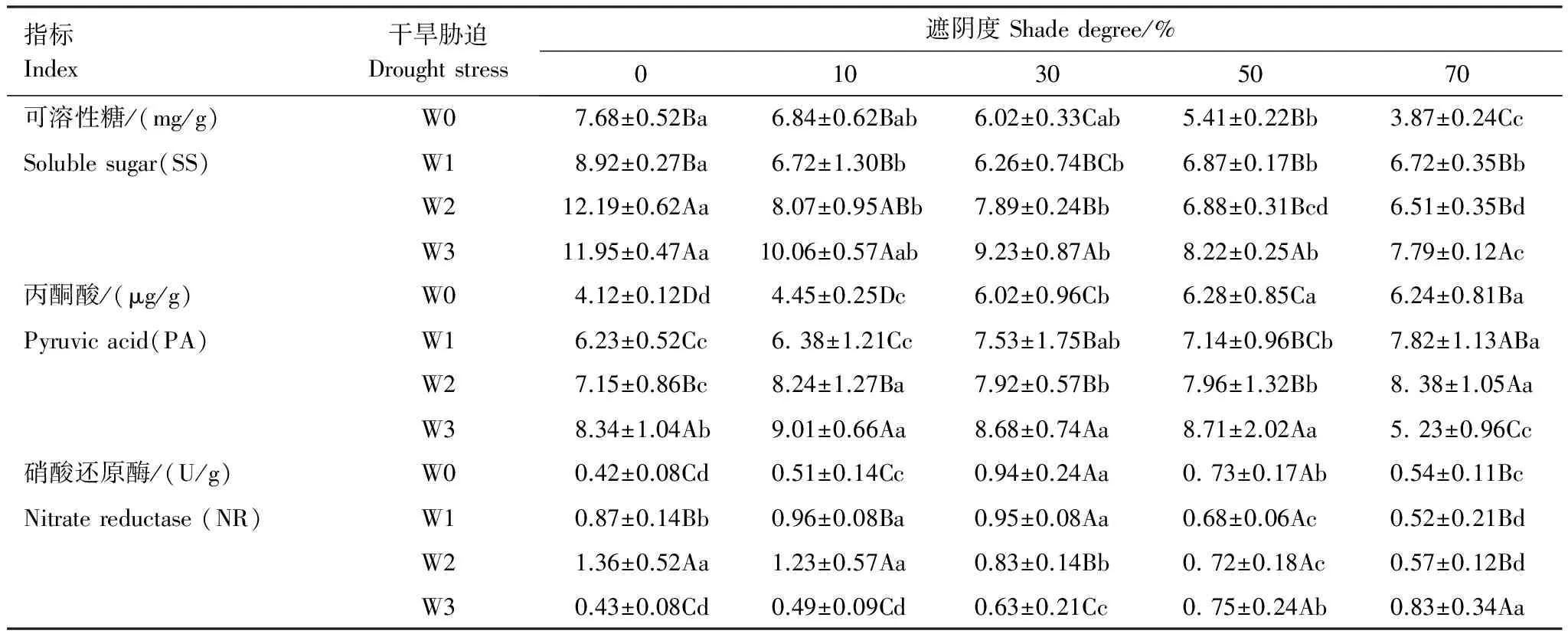
表3 遮阴和干旱对代谢指标的影响
方差分析表明,遮阴或干旱对SS、PA和NR有极显著影响(表4)。遮阴影响顺序为NR>PA>SS,干旱影响顺序为NR>SS>PA。遮阴和干旱对SS、PA和NR有极显著协同影响(P<0.01),大小为PA>NR>SS。干旱是影响代谢指标的主要因子,协同作用次之,遮阴较小。氮代谢受干旱或遮阴胁迫的影响最大,糖代谢次之,呼吸作用较小。
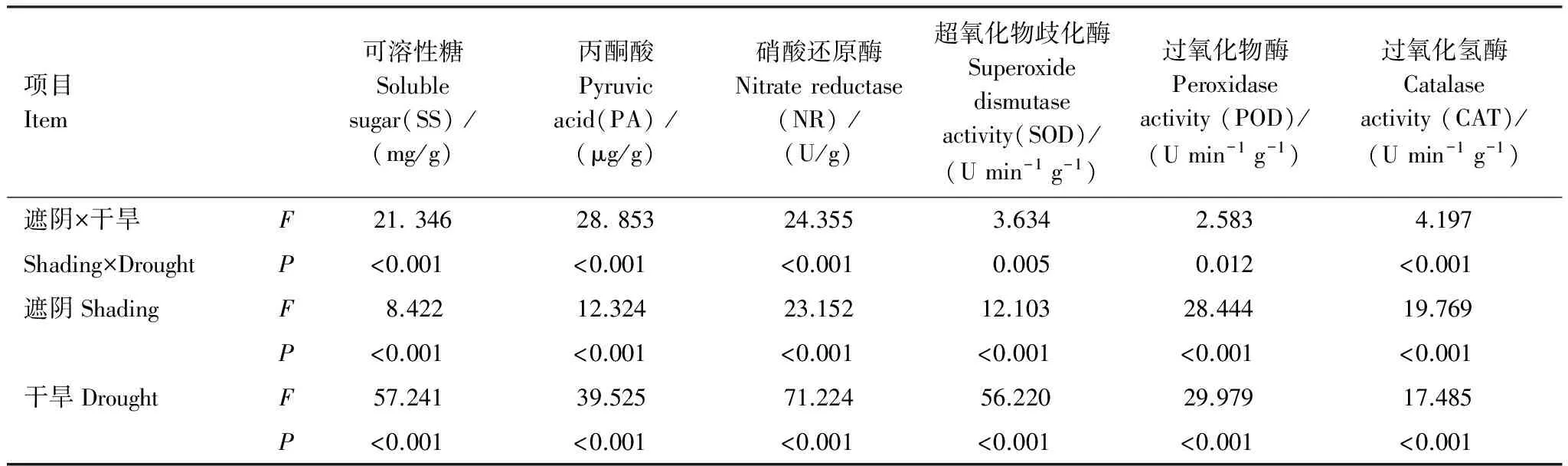
表4 代谢指标和抗性指标差异的双因子方差分析
2.3 遮阴和干旱对抗性指标的影响
遮阴、干旱和协同处理对SOD、POD和CAT活性有显著影响(P<0.05)(表5)。遮阴下,抗氧化酶活性随遮阴度增加先显著增加又显著降低。10%遮阴使POD显著增加;30%遮阴使SOD和CAT显著增加;50%遮阴下POD和SOD达最大值;遮阴度10%—50%间POD和CAT无显著差异,70%遮阴下SOD、POD和CAT活性则显著下降。
干旱下,抗性指标随干旱度增加先显著增加又显著下降。W1使SOD和POD显著增加(P<0.05),W2下SOD、POD和CAT显著增大到最大值,W3下显著下降到W0水平。
遮阴和干旱对SOD、POD和CAT活性有显著协同作用(P<0.05)。10%—30%遮阴提高了W1和W2下酶活性,提高了W3下SOD和POD活性。≥50遮阴对干旱下的酶活性影响差异较大。重度遮阴和重度干旱协同作用使SOD、POD和CAT均显著下降,W3+70%下SOD、POD和CAT分别为W0+0的73.08%、78.78%和42.94%,为各自最大值的35.06%、28.61%和28.32%。

表5 遮阴和干旱对抗性指标的影响
方差分析表明,遮阴或干旱对SOD、POD和CAT活性有极显著影响(P<0.01)(表4)。遮阴影响顺序为POD>CAT>SOD,干旱影响顺序为SOD>POD>CAT。遮阴和干旱协同处理对POD有显著影响,对SOD和CAT有极显著影响,协同作用大小为CAT>SOD>POD。干旱是影响抗性指标的主要因子,遮阴次之,协同作用较小。SOD受干旱影响大于遮阴,POD和CAT受遮阴和干旱影响差异较小。
2.4 遮阴和干旱对受伤害指标的影响
遮阴、干旱和协同处理对MDA、Pro、SP和H2O2含量有显著影响(P<0.05)(表6)。遮阴下,MDA、Pro、SP和H2O2含量随遮阴度增大逐步上升。10%遮阴使Pro含量显著增加,30%遮阴使MDA和H2O2含量显著增加(P<0.05);50%遮阴下MDA和Pro含量显著大于≤30%遮阴(P<0.05);70%遮阴时MDA、Pro、SP和H2O2含量均达最大值。
干旱下,MDA、Pro和H2O2含量随干旱度增加逐步上升,SP含量先增后降。W1时MDA、Pro、SP和H2O2含量显著大于W0(P<0.05),W2时H2O2和Pro含量显著高于W1,MDA和SP与W1下无显著差异;W3下MDA和H2O2含量显著大于W1和W2,SP含量显著低于W1和W2,Pro含量与于W1和W2下无显著差异。
遮阴和干旱对MDA、Pro、SP和H2O2含量有显著协同作用(P<0.05)。遮阴加重了干旱使MDA、Pro和H2O2含量上升的影响,干旱加重遮阴对MDA、Pro和H2O2含量的影响,遮阴对干旱下SP含量影响较小。≥30遮阴对干旱下受伤害指标的影响大于10%遮阴。W2+30%下SP含量达最大值,W2+70%下Pro含量达最大值。重度遮阴和重度干旱使MDA、Pro和H2O2含量显著增加,W3+70%下MDA、Pro和H2O2含量分别为W0+0的7.74、1.84、4.31倍。

表6 遮阴和干旱对受伤害指标的影响
方差分析表明,遮阴或干旱对MDA、Pro、SP和H2O2含量有极显著影响(P<0.01)(表7)。遮阴影响顺序为H2O2>MDA>Pro>SP,干旱影响顺序为H2O2>MDA>SP>Pro。遮阴和干旱对SP无协同作用,对Pro有显著协同作用,对H2O2和MDA有极显著协同作用(P<0.01),作用大小为H2O2>MDA>Pro>SP。干旱是影响细胞膜伤害指标的主要因子,遮阴次之,协同作用较小。干旱对H2O2、Pro和SP的影响大于遮阴,遮阴对MDA影响大于干旱。
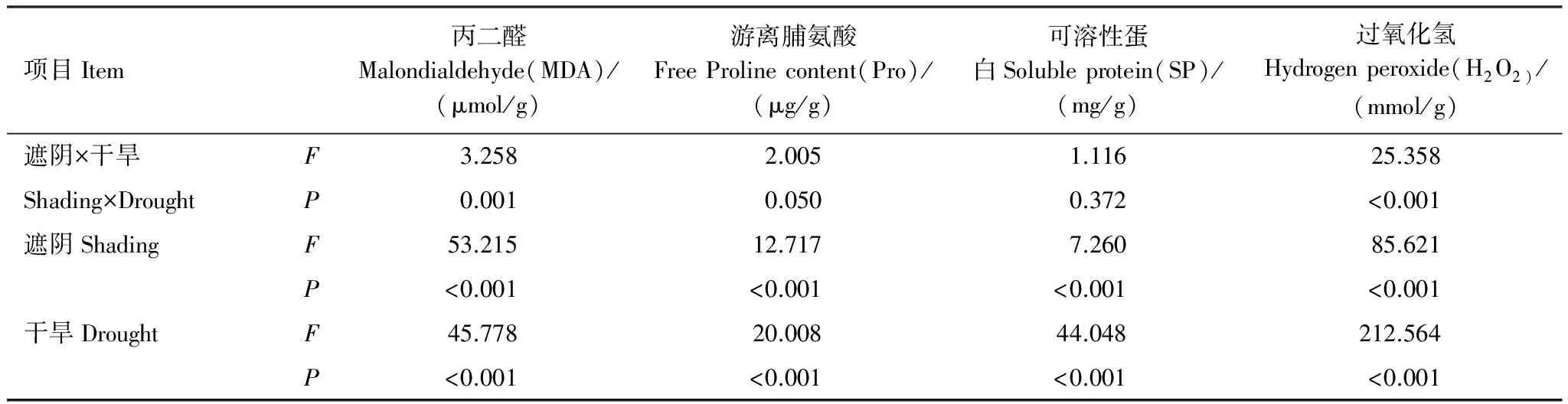
表7 受伤害指标差异的双因子方差分析
3 讨论
3.1 遮阴和干旱与光合作用的关系
光是植物生命活动的原初能量和动力来源,也是诱导、促进与调控植物的生长发育的基本信号。植物通过抬高叶片位置、变薄叶片厚度、增大叶片面积、改变叶片张角[10-13]、增加叶绿素含量,降低光补偿点、光饱和点和暗呼吸率等适应策略[14-15],增加遮阴下捕获光能和提升光合效益的能力。遮阴不仅降低光照强度和光照时间,也改变了微生境的气温、土温和叶表温度,空气湿度、土壤水分含量及叶片蒸腾速率等因素,影响了叶片的气孔大小、光合色素含量及光敏度和光响应能力,致使光合能力受到遮阴的影响[16]。遮阴下荩草叶性状、叶绿素含量、光响应能力发生变化[6],对荩草Pn、Tr、Ci和Gs等光合参数有显著影响。遮阴度引起荩草的叶绿体、气孔等自身微结构及温湿度等微生境产生差异,使Tr和Gs更易受遮阴影响。30%遮阴下Pn、Tr和Gs达最大值,表明荩草属喜阴植物。水是植物生命之源,是物质合成、转化和运输的重要载体。干旱直接影响物质和能量的吸收与合成及在功能间的分配,间接使叶片结构、色素含量、激素和酶活性发生变化,从而影响光合物质基础和光合效率。植物通过减少叶数量和叶面积、降低叶片位置[17-20]、关闭气孔等方式降低水分蒸发,通过改变光合器官和光合色素活性[21- 23]及光电子传递方式[24],提高吸收和转化光能效率和水分利用效率[25]应对干旱胁迫。本试验中,轻度干旱显著提高了Pn和Gs,中重度干旱使Pn、Tr、Ci和Gs显著降低,与短期或轻度干旱使光合作用降低[26]的观点不符,符合适度干旱可提高植物的光合能力[27-28],表明荩草具有较强的抗旱性。适度遮阴和适度干旱协同作用可提高荩草的光合能力,重度干旱和重度遮阴协同作用则显著降低了荩草的光合参数。W3+70%下光合速率仅为CK的20%,但植株不仅存活且无衰老表现,或许荩草采取了更为复杂的系统性光合策略,在水分和光照同时受限下,荩草调控光合作用及维持生长的机理,待于深入研究。
3.2 遮阴和干旱与物质代谢的关系
可溶性糖(SS)是碳水化合物代谢、暂时贮藏和运输的主要型式,也是呼吸作用的主要底物,与植物的抗性有密切关系。遮阴或干旱对SS含量的影响,因植物种类、生长阶段、测定器官、胁迫强度和持续时间等不同[16- 18]。本试验中,SS含量随遮阴或干旱变化与Pn变化趋势不同,表明遮阴或干旱不仅通过光合作用影响糖代谢,也通过SS的合成或分解、运转和累积,影响呼吸作用和细胞渗透压。随遮阴或干旱加重,PA含量显著上升,表明呼吸作用为抵御伤害提供能量。遮阴下SS含量减小和PA含量增大,SS主要参与呼吸作用。干旱下SS和PA含量同时增加,SS不仅参与呼吸作用,还要参与调节渗透压以维持细胞的完整性。硝酸还原酶(NR)对无机氮的还原、同化和氨基酸、蛋白质的合成等氮代谢过程起关键的调控作用[29],氮代谢与糖代谢和呼吸作用密切相关[30]。遮阴或干旱下NR活性与SS和PA含量变化趋势不一致,氮代谢与糖代谢和呼吸作用同步性较差。或许胁迫条件下,氮代谢主要参与维持生命运转的遗传物质和生理活性物质合成,而降低了与糖代谢和呼吸作用的相关性。遮阴或干旱下NR活性存在显著的阈值效应,超过范围的遮阴或干旱胁迫使NR活性下降。遮阴和干旱对荩草物质代谢有显著协同作用,使糖代谢和氮代谢比单一胁迫有所降低,但显著提高了呼吸作用。生境胁迫对糖代谢、氮代谢及呼吸作用的影响是极其复杂的系统性过程,本文通过SS和PA含量及NR活性变化,仅说明胁迫强度对荩草代谢速度的影响,表明遮阴和干旱对糖代谢、氮代谢及呼吸作用的有协同影响,但难以论述和推断NR活性、SS和PA含量与具体代谢过程的关系,与代谢速度与生长发育及抗逆性之间的关系。
3.3 遮阴和干旱与抗氧化酶活性
正常植物细胞内自由基的产生和清除处于动态平衡状态。当受到生境胁迫时,代谢速度加快引起活性氧积累,抗氧化系统则提高清除自由基酶的活性。SOD主要使O2-转化为H2O2和O2,POD和CAT将H2O2分解为H2O,三者共同协调作用消除过多的活性氧保护植物免受伤害[31-32]。SOD、POD和CAT活性的变化顺序和增长幅度受植物抗性、植物种类、胁迫强度等诸多因素的影响。本试验中,10%遮阴或轻度干旱使POD显著增加,说明POD活性易受胁迫诱导。50%遮阴或中度干旱使SOD、POD和CAT均活性达最大值,表明荩草抗氧化酶系统有显著的阈值限制,难以清除70%遮阴或重度干旱下产生的活性氧。遮阴与干旱对荩草的胁迫机理不同,导致糖代谢、氮代谢及呼吸作用强度不同,代谢中产生活性氧的种类与数量差异,使遮阴对抗氧化酶系统的影响为POD>CAT>SOD,干旱的影响顺序为SOD>POD>CAT。遮阴和干旱对SOD、POD和CAT活性有显著协同作用,10%—30%遮阴提升干旱下荩草抗氧化酶的活性,≥50遮阴对干旱度下荩草的SOD、POD和CAT活性影响表现出多样化,表明遮阴和干旱共同处理使抗氧化酶系统作用过程更加复杂。20个遮阴和干旱协同作用下,荩草抗氧化酶活性存在数倍差异,SOD、POD和CAT活性相互此消彼长,使抗氧化酶总量维持较高水平,说明荩草具有极其强大的抗氧化酶系统及适应环境能力。
3.4 遮阴和干旱与细胞膜完整性的关系
当产生的活性氧超过保护酶系统的清除能力,引起细胞膜透性增大、大分子生命物质受损,导致植物生理紊乱和功能受伤。随遮阴或干旱胁迫加重,荩草体内H2O2含量急剧上升约5倍,而抗氧化酶活性仅增2倍左右,导致活性氧大量累积对细胞膜造成伤害。植物通过生理生化变化,积累有机小分子物质,降低细胞内的渗透势,保护细胞膜的完整性[33]。可溶性糖(SS)、可溶性蛋白(SP)、游离脯氨酸(Pro)和丙二醛(MDA)是减轻逆境伤害的渗透调节物质,几种物质的来源与作用机理不尽相同。SS和SP既是营养物质也能提高细胞的保水能力和调节组织渗透势,对生命物质及生物膜起保护作用,SS和SP受光合生产、碳代谢及氮代谢的共同影响[32]。随遮阴度增大SS含量减少、SP含量增加,表明氮代谢对遮阴下荩草的渗透势调节贡献比糖代谢更大。轻中度干旱时SS和SP含量同步增加,重度干旱时SP含量急剧下降,说明干旱下糖代谢比氮代谢对渗透压的调节能力更强。Pro对稳定原生质胶体、生物大分子结构及组织内的代谢过程、降低细胞酸性和凝固点、防止细胞脱水、解除氨毒及调节细胞氧化还原势等方面起重要作用,Pro含量明显受植物抗性的影响[34-35]。荩草中Pro含量随遮阴和干旱度增大逐步显著增加,表明强胁迫下Pro充当重要的渗透调节物质。遮阴和干旱对SS和Pro有显著协同作用,对SP无协同作用,表明渗透性物质产生与累积是相当独立的复杂过程。MDA是膜脂过氧化最重要的产物,MDA随遮阴和干旱度增加而不断增大,反映出胁迫越大对膜系统受损越大。本试验发现,遮阴或干旱或协同处理使荩草中H2O2含量快速提高,虽抗氧化酶系统和渗透调节系统积极响应清除自由基以维持细胞完整性,但MDA对胁迫的敏感度与提升量高于Pro、SP和SS等调节物质,遮阴和干旱对H2O2和MDA有极显著协同作用,使细胞膜损伤度随协同胁迫组合加重而加大。虽一定范围的遮阴和干旱叠加处理,可提高荩草细胞中Pro、SS和SP等维持渗透压物质的含量,以减少对细胞的伤害,但同时也提高了H2O2和MDA含量,可见胁迫生境对细胞膜有不可逆性的伤害。
4 结论
遮阴和干旱对荩草光合作用、物质代谢、抗性系统和细胞膜完整性有极显著协同作用。荩草可依据遮阴和干旱的胁迫程度,通过调整光合参数,充分利用有限光照和水分资源完成光合作用,为植株生长发育提供能量。通过改变糖代谢、氮代谢及呼吸作用速度,使物质与能量在生长发育和抵御胁迫功能间进行分配。通过激活抗氧化酶系统及酶活性,改变渗透压物质组成与含量,减少胁迫伤害和保持细胞完整性。荩草具有极强的抗旱性和耐阴性,通过光合策略、代谢策略、抗性策略使植株能逐步适应或应对生境胁迫,适度遮阴和干旱利于荩草生长发育,重度遮阴和重度干旱虽可存活,但对细胞膜有不可逆的潜在伤害。遮阴和干旱对生理代谢及抗性系统影响的协同作用,提高了荩草的抗逆潜力和生态适应能力。
参考文献(References):
[1] 赵艳, 蔡捡, 李莹, 刘金平. 3种冷季型草坪草对温度与遮荫协同作用响应的差异性分析. 草业与畜牧, 2015, (2): 28- 33.
[2] 赵燮京, 刘定辉. 四川紫色丘陵区旱作农业的土壤管理与水土保持. 水土保持学报, 2002, 16(5): 6- 10, 16- 16.
[3] 刘金平. 坡度对野生荩草分株特征及生殖分配的影响. 草业科学, 2013, 30(10): 1602- 1607.
[4] 曾晓琳, 王大伟, 刘金平, 王思思, 范宣. 坡向对3种冷季型草坪草表观性状及叶绿素含量的影响. 草业科学, 2015, 32(11): 1823- 1831.
[5] 刘金平, 张海燕. 南充地区野生荩草种质资源形态多样性研究. 安徽农业科学, 2008, 36(12): 4926- 4928.
[6] 张小晶, 蔡捡, 刘金平, 李莹, 曾晓琳. 不同遮阴度对荩草构件性状和生物量分配影响的差异性分析. 西南农业学报, 2015, 28(6): 2720- 2725.
[7] 周旭, 齐代华, 蒋宣斌, 贺丽, 闵鹏, 梁洪海, 卢峰. 不同程度石漠化生境中矛叶荩草Arthraxonprionodes茎形态适应性研究. 西南大学学报: 自然科学版, 2016, 38(11): 40- 49.
[8] 刘金平, 范宣, 游明鸿, 王思思, 宗人旭. 老芒麦种子发育过程中生殖枝中糖分、丙酮酸含量及硝酸还原酶活性变化. 草业学报, 2016, 25(5): 69- 77.
[9] 王学奎. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2006: 176- 286.
[10] Ferree D C, McArtney S J, Scurlock D M. Influence of light on fruit set of French hybrid grapes. HortScience, 1998, 33(3): 510- 511.
[11] Bell G E, Danneberger T K, Mcmahon M J. Spectral irradiance available for turfgrass growth in sun and shade. Crop Science, 2000, 40(1): 189- 195.
[12] 陶建平, 钟章成. 光照对苦瓜形态可塑性及生物量配置的影响. 应用生态学报, 2013, 14(3): 336- 340.
[13] 张哲, 杜桂娟, 马凤江, 杨姝, 刘洋. 遮阴对5种豆科牧草形态和生理指标影响的初探. 草业科学, 2011, 28(7): 1296- 1300.
[14] 尹慧, 安莹, 陈雅君, 陈阳, 孙佳林, 李静静, 李艳侠. 不同遮阴强度下白三叶形态特征和生长动态. 中国草地学报, 2015, 37(5): 86- 91.
[15] 张会慧, 张秀丽, 许楠, 贺国强, 金微微, 岳冰冰, 李鑫, 孙广玉. 外源钙对干旱胁迫下烤烟幼苗光系统Ⅱ功能的影响. 应用生态学报, 2011, 22(5): 1195- 1200.
[16] 王艺, 韦小丽. 不同光照对植物生长、生理生化和形态结构影响的研究进展. 山地农业生物学报, 2010, 29(4): 353- 359, 370- 370.
[17] 王齐, 孙吉雄, 安渊. 水分胁迫对结缕草种群特征和生理特性的影响. 草业学报, 2009, 18(2): 33- 38.
[18] 金不换. 干旱胁迫对不同品种早熟禾形态和生理特性影响的研究[D]. 哈尔滨: 东北农业大学, 2009.
[19] 刘金平, 游明鸿, 段婧, 张丽慧. 水分胁迫下雌雄异株植物葎草繁殖策略的可塑性调节. 草业学报, 2015, 24(3): 226- 232.
[20] 温翠平, 李威, 漆智平, 唐树梅. 水分胁迫对王草生长的影响. 草业学报, 2012, 21(4): 72- 78.
[21] 张金政, 张起源, 孙国峰, 何卿, 李晓东, 刘洪章. 干旱胁迫及复水对玉簪生长和光合作用的影响. 草业学报, 2014, 23(1): 167- 176.
[22] 赵瑾, 白金, 潘青华, 金洪. 干旱胁迫下圆柏不同品种(系)叶绿素含量变化规律. 中国农学通报, 2007, 23(3): 236- 239.
[23] 陈暮初. 朱砂根种子萌发生理及苗木对干旱和光照胁迫的生理响应[D]. 雅安: 四川农业大学, 2009.
[24] 韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性. 生态学报, 2010, 30(15): 4019- 4026.
[25] 陆燕元, 马焕成, 李昊民, 邓西平. 土壤干旱对转基因甘薯光合曲线的响应. 生态学报, 2015, 35(7): 2155- 2160.
[26] Dai Y J, Shen Z G, Liu Y, Wang L L, Hannaway D, Lu H F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content ofTetrastigmahemsleyanumdiels et gilg. Environmental and Experimental Botany, 2009, 65(2): 177- 182.
[27] 邓云鹏, 雷静品, 潘磊, 王晓荣. 不同种源栓皮栎光响应曲线的模型拟合及参数比较. 生态学杂志, 2016, 35(2): 387- 394.
[28] 王永健, 方兴, 钟章成. 不同生境对蝴蝶花花部与果实特征的影响. 生态学报, 2010, 30(17): 4628- 4635.
[29] 马春晖, 韩建国, 孙洁峰, 王栋. 结缕草种子发育过程中生理生化变化的研究. 草业学报, 2009, 18(6): 174- 179.
[30] Ching T M, Crane J M, Stamp D L. Adenylate energy pool and energy charge in maturing rape seeds. Plant Physiology, 1974, 54(5): 748- 751.
[31] de Azevedo Neto A D, Prisco J T, Enéas-Filho J, de Abreu C E B, Gomes-Filho E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environmental and Experimental Botany, 2006, 56(1): 87- 94.
[32] 李霞, 阎秀峰, 于涛. 水分胁迫对黄檗幼苗保护酶活性及脂质过氧化作用的影响. 应用生态学报, 2005, 16(12): 2353- 2356.
[33] Ramanjulu S, Bartels D. Drought-and desiccation-induced modulation of gene expression in plants. Plant, Cell and Environment, 2002, 25(2): 141- 151.
[34] 唐钢梁, 李向义, 林丽莎, 李磊, 鲁建荣. 骆驼刺在不同遮阴下的水分状况变化及其生理响应. 植物生态学报, 2013, 37(4): 354- 364.
[35] 刘红云, 梁宗锁, 刘淑明, 董娟娥. 持续干旱及复水对杜仲幼苗保护酶活性和渗透调节物质的影响. 西北林学院学报, 2007, 22(3): 55- 59.
猜你喜欢
氯碱工业(2021年6期)2021-12-25 19:46:27
中成药(2021年5期)2021-07-21 08:38:32
天津医科大学学报(2021年1期)2021-01-26 00:57:20
科学大众(2020年23期)2021-01-18 03:09:08
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:18
上海包装(2019年2期)2019-05-20 09:10:52
汽车观察(2019年2期)2019-03-15 06:00:50
中成药(2018年2期)2018-05-09 07:19:49
中国卫生(2016年5期)2016-11-12 13:25:26
大型铸锻件(2015年5期)2015-12-16 11:43:22
