
模拟放牧强度与施肥对青藏高原高寒草甸群落特征和物种多样性的影响
2018-04-16 05:58:34张璐璐王孝安朱志红李英年
生态环境学报 2018年3期
张璐璐,王孝安,朱志红*,李英年
1. 陕西师范大学生命科学学院,陕西 西安 710062;2. 中国科学院西北高原生物研究所,青海 西宁 810008
草地生态系统是陆地生态系统的重要组成部分,刈割作为放牧的一种人为替代方式,已逐渐成为草地生态系统的主要利用方式(王国良等,2012)。适度刈割提高了草地群落空间异质性并降低了物种间的竞争强度,从而提高物种多样性与丰富度(Yamauchi et al.,2004;Hooper et al.,2005;高本强等,2014),并可以在保证草地群落稳定的前提下,充分发挥草地生态系统的再生性能,提高草地群落生产潜力(刘美丽等,2016)。养分是陆地生态系统的重要限制因子,同时也是土壤养分含量和植物群落特征的重要决定因子(魏金明等,2011)。增加土壤肥力可显著提高初级生产力、解除土壤对植物生长的营养限制作用、改变物种间固有的关系(Rajaniemi,2002)。因此,刈割与施肥能够通过改变土壤肥力与植物生长状态而改变群落结构及其物种多样性水平,以达到维持生态系统的生产力并促进其可持续发展的目的(高本强等,2014)。然而,目前大量研究表明,过度放牧会引起物种多样性和初级生产力的同时降低(韩国栋等,2007;卢黎黎,2010;高本强等,2014),不利于草原生态系统的维持和更新(刘美丽等,2016),最终导致草地退化(石红霄等,2014)。同样地,施肥在提高初级生产力的同时,会使草地植物群落物种组成贫乏,群落结构趋于简单,物种多样性减少(李晓彤,2016)。因此,研究不同刈割与施肥梯度下草地物种多样性和生产力的变化规律,对生态环境的保护、恢复和重建具有实际意义。
高寒草甸是中国天然草地中最重要的区域性类型之一,但近几十年来,由于气候变化、人类干扰的影响,使得种群生境恶化,群落种类成分多样性降低,结构简单化,生产力下降,草地退化问题也日趋严重(张靖乾等,2008)。而超载过牧或不合理利用正是导致草地普遍退化的主导因素(Kim et al.,2006),且随着人类活动干扰的加剧,物种多样性的丧失已经成为最严重的生态问题之一,并受到了生态学者的广泛关注(黄丽娜等,2009)。其中,变化环境中群落物种多样性和生态系统初级生产力的关系又是当代生态学研究中最有争论的问题之一(杜家丽,2016)。鉴于此,对不同刈割和施肥处理下的高寒草甸植物群落展开研究,旨在探讨不同刈割强度和土壤资源对高寒草甸群落特征、多样性水平、生产力水平的影响,以深入理解物种多样性和草地生态系统生产力的变化及相互关系,为高寒草甸天然草场的合理利用和生物多样性维持提供基础数据。
1 材料与方法
1.1 研究区概况
本研究在中国科学院海北高寒草甸生态系统定位站(37°29′~37°45′N,101°12′~101°23′E)进行。该定位站位于青海省海北藏族自治州门源回族自治县,海拔为 3200 m。定位站气温极低,全年无绝对无霜期,相对无霜期约为20 d,年平均气温仅为-1.6 ℃,年平均降水量为578.1 mm,主要分布在5—9月,占年降水量的79%(李英年等,1998)。土壤为高山草甸土,由于微生物活动较弱,土壤有机质分解缓慢(乔安海等,2006)。研究样地属典型的矮篙草(Kobresia humilis)草甸,是在长期放牧利用下形成的放牧偏途顶极群落(周兴民等,1982),优势种为矮嵩草,次优势种有异针茅(Stipa aliena)、美丽风毛菊(Saussurea superba)、紫羊茅(Festuca rubra)、垂穗披碱草(Elymus nutans)、麻花艽(Gentiana straminea)、青海风毛菊(Saussurea kokonorensis)、柔软紫苑(Astar flaccidus)、异叶米口袋(Gueldenstaedtia diversifolia)等(李英年等,1998)。
1.2 实验设计
本研究样地建于2007年4月,占地0.6 km2(100 m×60 m)。试验设置3个刈割水平,2个施肥水平、3区组的裂区嵌套设计,每区组3个4 m×4 m的大样方,共计27个大样方。试验主区为刈割处理(HC:重度刈割留茬1 cm;MC:中度刈割留茬3 cm;NC:不刈割),于每年6月中旬进行。嵌套在主区之中的副区处理为施肥,包含4个2 m×2 m的副区样方,利用2 m×0.25 m雪花铁皮十字形纵切嵌入草地进行隔离隔,其中包括2个施肥(F)处理和2个不施肥(NF)处理。刈割和施肥在副区样方中央 1.5 m×1.5 m范围内进行。在每个副区样方内再设4个0.5 m×0.5 m的小样方,其中1个为永久样方,用于群落调查;在剩余的3个小样方中随机选取1个进行取样。施肥分别于每年5月、6月和7月中旬进行,施肥量为建植人工草地时的最优施肥量(乔安海等,2006)。每次施用尿素4.25 g·m-2,磷酸二铵1.02 g·m-2,年添加 N 2.782 g·m-2,P 0.857 g·m-2。
1.3 取样与测定
2012年 6月中旬刈割后,将收获的部分置于60 ℃烘箱中烘72 h后称重。8月中旬采用目测法测定每个样方的总盖度及样方内各物种的盖度,同时测定各样方的分株密度和高度。8月下旬对所有处理进行刈割收获,将刈割部分置于 60 ℃烘箱中烘 72 h,称重。以6月中旬和8月下旬收获的生物量之和记作群落地上生物量,用于估计群落生产力。
本研究比较了群落中豆科、禾本科、莎草科与其他杂草4个功能群的重要值变化。豆科植物包括青海苜蓿(Medicago archiducis-nicolai)、甘肃棘豆(Oxytropis kansuensis)、黄花棘豆(Oxytropis ochrocephala)、宽苞棘豆(Oxytropis latibracteata)、高山豆(Tibetia himalaica)、祁连山黄芪(Astragalus chilienshanensis)、披针叶黄华(Thermopsis lanceolata)和小花棘豆(Oxytropis glabra);禾本科包括异针茅、早熟禾(Poa annua)、垂穗披碱草、羊茅(Festuca ovina)、双叉细柄茅(Ptilagrostis dichotoma)、洽草(Koeleria cristata)和太白细柄茅(Ptilagrostis concinna);莎草科包括矮嵩草、粗喙薹草(Carex scabrirostris)和双柱头藨草(Scirpus distigmaticus);其他杂草包括菊科、龙胆科、玄参科、蔷薇科以及毛茛科等除豆科、禾本科和莎草科外的所有物种。
1.4 数据计算与统计分析
1.4.1重要值
物种重要值(IV)=(相对高度+相对密度+相对盖度)/3(王国良等,2012)
功能群重要值=(功能群相对高度+功能群相对密度+功能群相对盖度)/3;
1.4.2物种多样性
采用丰富度指数(R)、Simpson优势度指数(D)和Shannon-Weaner多样性指数(H′)评价研究区物种多样性水平,其计算公式(马克平等,1994)分别为:

式中,S为物种数;N为群落内所有物种的个体数之和;Pi为群落内属于种i的物种个体数与所有物种总个体数的比例。
1.4.3统计分析
采用SPSS 13.0-GLM-Univariate-ANOVA,以嵌套裂区设计的混合模型分析刈割、施肥及其交互作用对所测指标的影响。模型的主区变异来源为刈割和区组,副区变异来源为施肥、刈割与施肥及刈割与区组的交互作用。多重比较采用Duncan氏检验。
采用 SPSS 13.0-Pearson correlate以及 Linear regression对不同刈割和施肥处理群落地上净初级生产力和物种多样性进行相关性和回归分析。
方差分析前对数据进行对数转化,并进行数据的正态性和方差齐性检验。统计检验的显著水平设为0.05。
2 结果与分析
2.1 群落组成及重要值
经过连续6年的刈割和施肥处理,共观察到75种植物,隶属25个科、54个属,其中以菊科物种数最多,占总物种数的17.33%,毛茛科、豆科、禾本科、龙胆科、玄参科、蔷薇科和莎草科中出现的物种在总物种数中也占一定比例,分别为 12%、10.67%、9.33%、8%、6.67%、5.33%和 4%,其他的大多数科则仅有1~2个植物种出现(表1)。
不同的刈割强度对物种组成存在一定的影响,不刈割处理区共发现 69种植物,群落以异针茅、早熟禾、钝苞雪莲瑞苓草(Saussurea nigrescens)和垂穗披碱草为主,同时伴生种有矮嵩草、美丽风毛菊、柔软紫菀、圆萼刺参(Morina chinensis)、麻花艽等莎草和杂草类(表 1)。而中度刈割区发现群落中仅有 63种植物,其中以异针茅、早熟禾等禾草以及美丽风毛菊、矮嵩草为主,伴生种有矮火绒草(Leontopodium nanum)、直梗高山唐松草(Thalictrum alpinum)、青海苜蓿(Medicago archiducis-nicolai)等杂草类(表1)。重度刈割区发现了 65种植物,其主要物种组成与中度刈割区相同,但伴生种主要为矮火绒草、蒲公英(Taraxacum mongolicum)、钝苞雪莲瑞苓草、疏齿银莲花(Anemone obtusiloba)、青海苜蓿等(表1)。此外,刈割还导致了一些新的物种出现,如小花棘豆、露蕊乌头(Aconitum gymnandrum)、黑柴胡(Bupleurum smithii)、蓝苞葱(Allium atrosanguineum)等,而太白细柄茅、黄帚橐吾(Ligularia virgaurea)和美花草(Callianthemum pimpinelloides)均在刈割后消失(表1)。

表1 不同刈割和施肥处理下植物群落物种组成及其重要值Table 1 Species composition and their importance value of plant communities under different clipping and fertilizing intensity

续表1Continued Table 1
2.2 功能群重要值
如图 1所示,刈割显著降低了禾本科重要值(F(2,4)=9.665,P=0.029),却使豆科和杂草的重要值显著增加(F(2,4)=9.651,P=0.029;F(2,4)=8.112,P=0.039);施肥在增加禾本科重要值的同时(F(1,6)=26.134,P=0.036),显著降低了豆科(F(1,6)=281.062,P=0.000)和杂草(F(1,6)=22.761,P=0.041)的重要值;刈割(F(2,4)=0.162,P=0.856)和施肥(F(1,6)=0.045,P=0.851)对莎草科重要值均无显著影响。
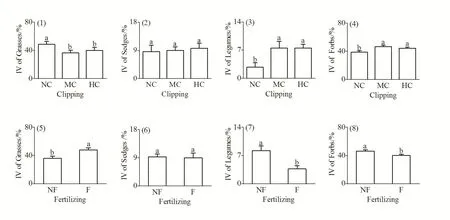
图1 刈割、施肥对禾本科、莎草科、豆科及杂草功能群重要值的影响Fig. 1 Effects of clipping, fertilizing on the importance value of functional group of Grasses, Sedges, Legumes and Forbs
2.3 群落数量特征
如图2所示,中度和重度刈割下群落盖度分别比不刈割处理增加了23.8%和19.3%(F(2,4)=11.649,P=0.021),群落密度分别比不刈割处理增加了36.7%和43.6%(F(2,4)=7.880,P=0.041),而群落高度却分别比不刈割处理降低了 60.7%和 69.2%(F(2,4)=58.950,P=0.001);重度刈割处理的群落生产力显著高于不刈割处理(F(2,4)=12.519,P=0.019);施肥使群落盖度(F(1,6)=107.314,P=0.000)、高度(F(1,6)=61.180,P=0.000)及生产力(F(1,6)=174.102,P=0.000)显著增加;;,却对群落密度影响不显著(F(1,6)=0.028,P=0.873)。
2.4 群落物种多样性
方差分析结果(图 3)显示,群落丰富度指数(F(2,4)=8.192,P=0.039)、Simpson指数(F(2,4)=13.300,P=0.002)和Shannon-Wiener指数(F(2,4)=13.448,P=0.017)均在刈割后增加;,且丰富度指数和Simpson指数在中度刈割后最高,但物种多样性在施肥后均显著降低(F(1,6)=49.614,P=0.000;F(1,6)=15.354,P=0.008;F(1,6)=60.726,P=0.000)。
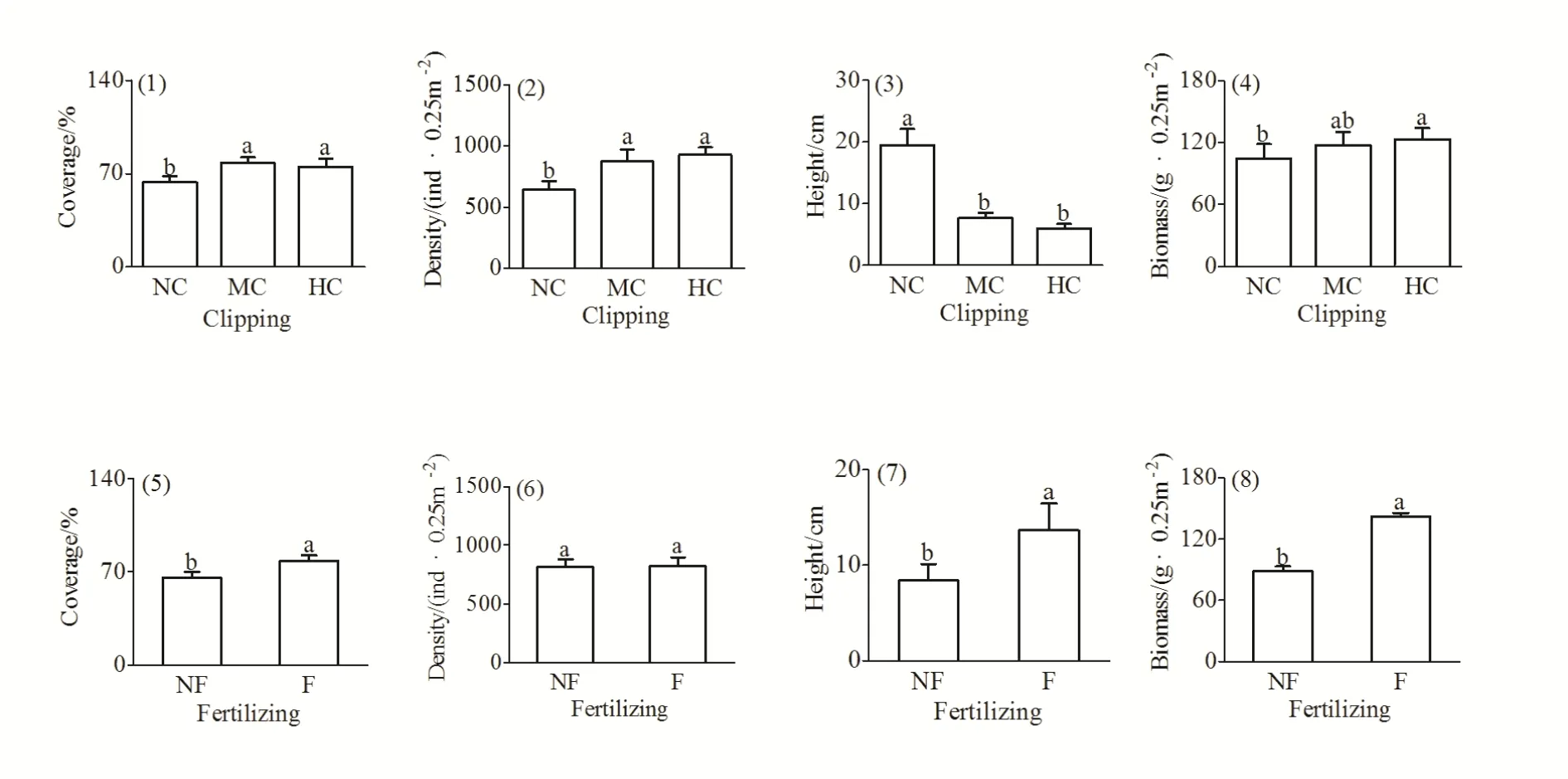
图2 刈割、施肥对群落盖度、密度、高度及地上生物量的影响Fig. 2 Effects of clipping and fertilizing on the coverage, density, height and the aboveground biomass of community
2.5 生产力与物种多样性的关系
如图4所示,在不刈割(NC)与施肥(NF、F)、中度刈割(MC)与施肥(NF、F)以及重度刈割(HC)与施肥(NF、F)3个处理梯度上,群落丰富度指数和 Shannon-Wiener指数与生产力的关系完全一致,均表现出极显著负相关关系;而Simpson指数与生产力的关系在不刈割(NC)与施肥(NF、F)处理梯度上则表现为微弱的负相关,在中度刈割(MC)与施肥(NF、F)和重度刈割(HC)与施肥(NF、F)处理梯度上分别表现出极显著负相关和显著负相关。
由图4还可知,在不施肥(NF)与刈割(NC、MC、HC)处理梯度上,群落丰富度指数、Shannon-Wiener指数和Simpson指数均与生产力表现出无显著相关性,而在施肥(F)与刈割(NC、MC、HC)处理梯度上,生产力与丰富度指数之间表现出微弱的正相关关系,与Shannon-Wiener指数和Simpson指数表现出无显著相关。
3 讨论
3.1 刈割、施肥对群落特征的影响
刈割对植物群落的影响主要表现为种群密度、群落种类组成、结构和生物量等的变化(钟秀琼等,2007),首先体现在草地植物群落特征的变化上(段敏杰等,2010)。根据天然草原优良禾本科牧草及主要毒杂草重要值的变化,可以看出每种植物在群落中的重要性,同时也在一定程度上反映天然草原偏离顶级群落的程度(赵成章等,2004)。本研究结果表明,在不同刈割处理下群落的物种组成发生了明显的变化,与对照相比,刈割后草地中矮嵩草的优势地位明显增强,同时一些适口性差的毒杂草的优势地位也相对增强(表 1)。从功能群重要值来看,刈割后,禾本科植物的重要值表现出降低的趋势,豆科和杂草的重要值显著增加,这与前人的研究结果一致(段敏杰等,2010;靳茜等,2015)。由于禾本科牧草具有根系发达、分蘖能力较强、植株高大、冠幅宽大等生物学特性,其在不刈割群落中占绝对优势(石红霄等,2014),但刈割后由于高大草类优势消失,使矮生植物得到充足光照,而中下层植物本身耐牧性强,因此刈割使草层中低矮牧草的优势度显著提升(Wu et al.,2004)。石红霄等(2014)对三江源地区植物群落的特征分析表明,莎草科植物由于具有强烈的克隆生殖特性和较强的耐牧性使得其在刈割后重要值显著增加(石红霄等,2014),但本研究中莎草科植物的重要值在刈割后没有发生明显变化,可能是因为莎草科中某些物种(如矮嵩草)的重要值在刈割后增加而另一些物种(如粗喙苔草)重要值降低导致相抵消(表1)。由此可见,刈割使草地中一些优良牧草的优势地位大大降低,而适口性差的毒杂草的优势地位相对增加,如果长期进行重度刈割,势必会导致草地质量下降,甚至使整个群落向以劣质牧草为主的方向演替。
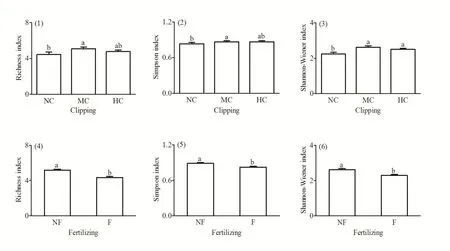
图3 刈割、施肥对群落丰富度指数、Simpson指数和Shannon-Wiener指数的影响Fig. 3 Effects of clipping, fertilizing on the Richness index, Simpson index and Shannon-Wiener index of community
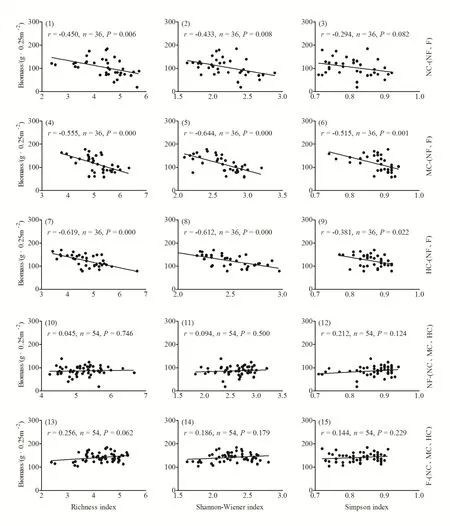
图4 不同刈割、施肥处理梯度下群落地上生物量与物种多样性之间的关系Fig. 4 Correlations between the aboveground biomass and species diversity of community for different clipping and fertilizing levels
草地群落数量特征与放牧强度密切相关(仁青吉等,2008),以往的诸多研究证明,不同强度的放牧或刈割扰动对草地群落密度、高度、盖度及地上生物量都会产生一定影响,并随干扰强度的加大其影响程度也逐渐加重(卢黎黎,2010;倪悦涵,2015;李晓彤,2016)。本研究中,刈割后群落盖度增加、密度增大、高度降低的结果与卢黎黎(2010)的研究一致(图2(1)~(3))。密度调节是植物响应刈割的主要机制之一(李晓彤,2016),刈割解除了顶端优势,许多植株“放弃”了有性繁殖,而选择增强其无性繁殖以适应刈割干扰,同时,刈割给杂类草物种的生长创造了机会,杂类草盖度的增大,导致刈割后草地总体盖度大于不刈割(卢黎黎,2010)。但群落刈割之后,株高显著下降,这是植物避免被啃食和刈割的一种响应策略(朱璧如等,2011;倪悦涵,2015)。生产力是生态系统最基本的数量特征之一,草地群落生产力的变化研究可以为人们了解草地生态系统的物质能量的分布格局提供基本资料,是实现草地持续利用和管理的重要理论依据(卢黎黎,2010)。周晓松等(2011)对矮嵩草草甸植物群落补偿生长的研究结果表明,刈割后多数不耐牧的高大禾草的生长受到抑制,表现出低补偿,而耐牧性强的莎草科植物以及大量中下层阔叶类杂草将获得充足的光照从而引发超补偿,最终导致群落地上净初级生产力的增加。本研究结果与之相符,刈割后群落生产力显著增加(图 2(4)),说明对草地进行适度的刈割管理,可以提高其空间异质性,有利于提高草地群落生产力(Yamauchi et al.,2004;Hooper et al.,2005)。
土壤养分通常是限制植物生长生殖的主要营养因素之一(李晓彤,2016),施肥增加地上净初级生产力的效应已得到广泛印证(高本强等,2014;李晓彤,2016;温超等,2017),特别是在高寒草甸,由于环境温度较低,土壤N、P养分的矿化率较低,可利用N、P含量仅占土壤全N、全P的1%左右,限制了植物的生长(杜家丽,2016),而施肥能有效解除土壤的养分限制,增加群落的盖度和高度(图 2(5),2(7)),最终导致群落生产力增加(图2(8))。卢黎黎(2010)对松嫩草地物种多样性与生产力关系的研究结果显示,施肥当年群落高度和群落盖度显著增加,而群落密度变化不显著,直到施肥第3年群落密度才显著增加(卢黎黎,2010),说明密度对土壤养分敏感度较差,以至于本研究中群落密度在施肥处理间未表现出显著差异(图2(6))。此外,施肥可以通过提高土壤中有效资源的水平,降低物种对限制资源竞争的强度,增加群落功能群的重要值(Schoolmaster et al.,2014)。施肥可使禾本科重要值得以提升的结论已得到多数学者的认同(李晓彤,2016;代景忠等,2017),当添加的氮、磷元素足够充分,禾本科植物对氮元素的竞争能力强于豆科对磷元素的竞争(Silvertown et al.,1987),从而导致重要值增加(图1(5)),使豆科功能群丧失主导地位,重要值出现下降(图1(7)),此外,禾本科植物高大、数量密集,对一些杂草类植物具有一定的遮阴作用,使它们对光的竞争能力减弱,所占重要值下降(图1(8)),这一结果与代景忠等(2017)研究相同,说明施肥量的增加有利于优势物种的生长,而且在一定程度上抑制了杂类草的生长,促进退化草地的正向演替(温超等,2017)。
3.2 刈割、施肥对物种多样性的影响
物种多样性的变化是对草地退化评价的一个重要指标,草地生态系统的稳定性发展和生产力的维持在很大程度上依赖于草地植物群落的生物多样性(Tilman et al.,2001;段敏杰等,2010)。在刈割扰动群落结构的机制中,不得不提到中度干扰假说,Grime(1977)将中度干扰假说定义为:外界的扰动(刈割)减弱了植物物种间的排斥过程,尤其是减弱了优势种对种内其他物种的竞争排斥作用,导致群落外物种的迁入和群落内物种的迁出。对植物群落进行中等程度干扰,可使植物群落物种丰富度和多样性达到最大值(Grime,1977;李晓彤,2016)。本研究结果表明,高寒矮嵩草草甸植物群落物种多样性在刈割后均增加,且丰富度指数和Simpson指数均在在中度刈割处理下达最大值(图 3(1)~(2)),这与前人研究结论一致(段敏杰等,2010;王国良等,2012;刘美丽等,2016),即群落物种多样性在中度刈割处理中最高,支持中度干扰假说,使中度干扰理论在高寒草甸中得到了证实。
土壤肥力既是影响生产力的因素,同样也是影响物种丰富度的重要因素,施肥可使得对土壤利用率高的优势种获得更大的优势,在群落中竞争能力增强,从而影响植物群落物种多样性(李晓彤,2016)。诸多研究表明,对草地长期施肥,在提高生产力的同时,会使植物群落的空间异质性或生态位维数降低,从而加剧物种间竞争并导致物种多样性降低(高本强等,2014;李晓彤,2016;温超等,2017)。Rajaniemi(2002)以不同的角度分析并解释施肥使物种多样性下降的原因。杨路存等(2015)认为施肥处理对高寒草甸物种丰富度、Simpson指数、Shannon-Weiner指数和Pielou均匀度指数影响变化一致,具有显著降低作用,本研究也得到了相同的结果,施肥后,高寒草地物种丰富度、Simpson指数和Shannon-Weiner指数均降低(图3(4)~(6)),因此在草地管理过程中应严格控制施肥量,以达到保护物种多样性的目的。
3.3 生产力与物种多样性的关系
生产力是生态系统功能的重要表现形式,物种多样性与生产力间的相互关系一直是生态学家们争执的热点,但至今没得出一致的结论(Hooper et al.,2005;韩国栋等,2007;王国良等,2012)。Gough et al.(2000)对美国6个LTER站点的研究结果的总结显示,大尺度范围内,只有一项研究表明草地生产力与物种多样性为单峰曲线关系;在小尺度范围内,45%的研究显示为无相关模式,20%表现为单峰关系,20%表现为正相关关系,15%表现为负相关(Gough et al.,2000;杜家丽,2016)。中国部分学者认为群落多样性指数与地上生物量之间呈“单峰型”曲线关系(韩国栋等,2007),但也有学者认为生产力与物种多样性呈负相关关系或者无相关性(靳茜等,2015)。总体而言,物种多样性与生产力间的关系主要表现为单调上升、单调下降和单峰格局,具体的机制还需在更广泛的领域进行研究和论证(王国良等,2012)。
从本研究结果来看,不同干扰下,群落生产力与物种多样性之间表现出4种关系模式,即显著负相关、微弱负相关、微弱正相关或不相关。其中,在不同施肥处理下,刈割使生产力与物种多样性呈现出微弱正相关或不相关(图 4(10)~(15)),而在不同刈割处理下,施肥使生产力与物种多样性呈现显著负相关或微弱负相关关系(图 4(1)~(9)),这一结果基本支持邱波等(2003)对生产力与多样性关系的观点,即生产力与多样性关系不同可能是环境因素的干扰导致的,当环境因素对生产力与多样性都起促进作用时,生产力与多样性表现为正相关;当环境因素对生产力与多样性中的一个起促进作用,一个起负面作用时,生产力与多样性就表现为负相关或没有明显关系。结合本研究结果,刈割使生产力、物种丰富度、Simpson指数和Shannon-Weiner指数均增加(图 2(4);图 3(1)~(3)),故生产力与物种多样性的关系表现为微弱正相关或无相关,而施肥使生产力显著增加(图2(8)),却使物种丰富度、Simpson指数和Shannon-Weiner指数显著降低(图 3(4)~(6)),故生产力与物种多样性的关系表现为显著负相关或微弱负相关。需要补充说明的是,当环境因素对生产力与多样性都起促进作用时,生产力与多样性也可能表现为不相关。因此,物种多样性-生产力关系可能是多变的,与群落所受扰动因素的类型和强度有关。
4 结论
通过对高寒矮嵩草草甸群落特征和物种多样性进行分析可知,刈割管理可以提高群落的盖度、密度及生产力,但是刈割后群落的物种组成发生了明显的变化,一些优良牧草的优势地位大大降低,而适口性差的毒杂草的优势地位相对增加。如果长期进行重度刈割,势必会造成草地质量下降,甚至使整个群落向以劣质牧草为主的方向演替。施肥能有效解除土壤的养分限制,增加群落盖度、高度和生产力,且施肥后,禾本科重要值增加、豆科和毒杂草重要值下降,说明施肥量的增加有利于优势物种的生长,而且在一定程度上抑制了毒杂草的生长,促进退化草地的正向演替。本结果有力支持了中度干扰假说,并暗示了物种多样性与生产力的关系可能是多变的,且这种关系取决于群落所受扰动因素的类型和强度。值得注意的是,施肥后,物种多样性显著降低,因此在草地管理过程中,放牧强度和营养添加应控制在合适的范围内,以达到保护物种多样性和合理利用的双重目的。
参考文献:
GOUGH L, OSENBERG C W, GROSS K L, et al. 2000. Fertilization effects on species density and primary productivity in herbaceous plant communities [J]. Oikos, 89(3): 428-439.
GRIME J P. 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory [J].American naturalist, 111(982): 1169-1194.
HOOPER D U, CHAPIN III F S,EWEL J J, et al. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge [J]. Ecological Monographs, 75(1): 3-35.
KIM E S, PARK D K, ZHAO X Y, et al. 2006. Sustainable management of grassland ecosystems for controlling Asian dusts and desertification in Asian continent and a suggestion of Eco-Village study in China [J].Ecological Research, 10(21): 907-911.
RAJANIEMI T K. 2002. Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses [J]. Journal of Ecology, 90(2): 316-324.
SCHOOLMASTER D R, MITTELBACH G G, GROSS K L. 2014.Resource competition and community response to fertilization: the outcome depends on spatial strategies [J]. Theoretical Ecology, 7(2):127-135.
SILVERTOWN J, LAW R. 1987. Do plants need niches? Some recent developments in plant community ecology [J]. Trends in Ecology andEvolution, 2(1): 24-26.
TILMAN D, REICH P B, KNOPS J, et al. 2001. Diversity and productivity in a long-term grassland experiment [J]. Science, 294(5543): 843-845.WU N,LLU J, YAN Z L. 2004. Grazing intensity on the plant diversity of alpine meadow in the eastern Tibetan plateau [J]. Rangifer, 15(4): 9-15.YAMAUCHI A, YAMAMURA N. 2004. Herbivory promotes plant production and reproduction in nutrient-poor conditions: effects of plant adaptive phenology [J]. American Naturalist, 163(1): 138-153.
代景忠, 闫瑞瑞, 卫智军, 等. 2017. 施肥对羊草(Leymus chinensis)割草场功能群物种丰富度和重要值的影响[J]. 中国沙漠, 37(3): 453-461.
杜家丽. 2016. 物种多样性和功能群多样性对地上净初级生产力的相对贡献[D]. 西安: 陕西师范大学: 1-46.
段敏杰, 高清竹, 万运帆, 等. 2010. 放牧对藏北紫花针茅高寒草原植物群落特征的影响[J]. 生态学报, 30(14): 3892-3900.
高本强, 袁自强, 王斌先, 等. 2014. 施肥和刈割对亚高山草甸物种多样性与生产力及其关系的影响[J]. 植物生态学报, 38(5): 417-424.
韩国栋, 焦树英, 毕力格图, 等. 2007. 短花针茅草原不同载畜率对植物多样性和草地生产力的影响[J]. 生态学报, 27(1): 182-188.
黄丽娜, 李文宾, 廉振民. 2009. 生物多样性的丧失及其保护[J]. 安徽农业科学, 37(5): 2217-2219.
靳茜, 安沙舟, 刘兴义, 等. 2015. 刈割对山地草甸草原群落特征及多样性的影响[J]. 新疆农业科学, 52(9): 1723-1730.
李晓彤. 2016. 施肥和刈割对羊草草地生产力与物种多样性的影响[D].长春: 东北师范大学: 1-42.
李英年, 王启基, 周兴民, 等. 1998. 高寒草甸植物群落的环境特征分析[J]. 干旱区研究, 15(1): 54-58.
刘美丽, 梁秀梅, 乌仁其其格. 2016. 刈割制度对围封羊草草地群落特征及生物量的影响[J]. 草业科学, 33(11): 2319-2325.
卢黎黎. 2010. 刈割对松嫩草地物种多样性——生产力关系及土壤特性的影响[D]. 长春: 东北师范大学: 1-28.
马克平, 刘玉明. 1994. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J]. 生物多样性, 2(4): 61-66.
倪悦涵. 2015. 黄河三角洲植物群落及功能性状对盐分和刈割的响应[D]. 济南: 山东大学: 1-61.
乔安海, 韩建国, 巩爱岐, 等. 2006. 氮肥对垂穗披碱草种子产量和质量的影响[J].草地学报,14(1):48-51.
邱波, 王刚. 2003. 生产力与生物多样性关系研究进展[J]. 生态科学,22(3): 265-270.
仁青吉, 崔现亮, 赵彬彬. 2008. 放牧对高寒草甸植物群落结构及生产力的影响[J]. 草业学报, 17(6): 134-140.
石红霄, 范月君, 侯向阳, 等. 2014. 三江源区围栏与放牧高山嵩草草甸植物群落特征分析[J]. 中国草地学报, 36(3):67-72.
王国良, 吴波, 杨秋玲, 等. 2012. 刈割对灌草丛草地植被物种多样性和生产力的影响[J]. 草地学报, 20(6): 1020-1025.
魏金明, 姜勇, 符明明, 等. 2011. 水、肥添加对内蒙古典型草原土壤碳、氮、磷及pH的影响[J]. 生态学杂志, 30(8): 1642-1646.
温超, 单玉梅, 高丽娟, 等. 2017. 施肥对科尔沁羊草割草场植物群落多样性和生产力的影响[J]. 江苏农业科学, 45(16): 250-254.
杨路存, 刘何春, 李长斌, 等. 2015. 氮磷钾不同施肥配方对退化高寒草原植物群落结构的影响[J]. 生态学杂志, 34(1):25-32.
张靖乾, 张卫国, 江小雷. 2008. 刈割频次对高寒草甸群落特征和初级生产力的影响[J]. 草地学报, 16(5): 491-496.
赵成章, 樊胜岳, 殷翠琴, 等. 2004. 毒杂草型退化草地植被群落特征的研究[J]. 中国沙漠, 24(4): 507-512.
钟秀琼, 钟声. 2007. 刈割对牧草影响的研究概况[J]. 草业与畜牧, (5):22-25.
周晓松, 朱志红, 李英年, 等. 2011. 刈割、施肥和浇水处理下高寒矮嵩草草甸补偿机制研究[J]. 兰州大学学报(自然科学版), 47(3): 50-57.
周兴民, 李键华. 1982. 海北高寒草甸生态系统定位站的主要植被类型及其地理分布规律.高寒草甸生态系统(第一集)[M]. 兰州: 甘肃人民出版社: 9-18.
朱璧如, 徐冰, 张大勇. 2011. 草地植物功能性状变异的程度和来源[J].北京师范大学学报(自然科学版), 47(5): 485-489.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
活力(2019年21期)2019-04-01 12:17:10
红土地(2016年3期)2017-01-15 13:45:17
水生生物学报(2015年1期)2015-02-28 16:01:05
海峡姐妹(2015年9期)2015-02-27 15:13:07
杭州科技(2014年3期)2014-02-27 15:26:46
