
烟青虫胚胎发育经历影响初孵幼虫的嗅觉偏好性
2018-04-10 09:35吴广庆郭线茹杨淑芳
安徽农业科学 2018年10期
吴广庆,郭线茹,杨淑芳*
(1.聊城大学东昌学院,山东聊城 252000;2.河南农业大学植物保护学院,河南郑州 450002)
昆虫的卵在发育生物学、生理学、生态学和进化等多学科的交叉点上占据独特的研究地位。卵是一个由卵壳包被的相对封闭的系统,其中幼虫的外周神经系统是随着胚胎发育的过程逐渐出现的。Woods[1]认为,胚胎发育经历的环境驱动了随后虫态尤其是幼虫阶段的表型可塑性。鳞翅目的初孵幼虫容易受到各种生物因子和非生物因子的胁迫[2],并且寄主植物被取食完或者从初生寄主植物上掉落地面之后,幼虫能够识别和评测一株陌生的寄主植物[3-4]。初生经历或许可以为初孵幼虫提供能够促进其寄主选择决策的信息[5]。目前,关于取食经历对随后幼虫取食行为以及成虫产卵行为的影响的研究报道较多[6-11]。海灰翅夜蛾(Spodopteralittoralis)成虫的求偶行为也会被自身幼虫期的取食经历调控[12]。然而,有关胚胎发育经历影响随后初孵幼虫寄主选择的试验证据相对比较缺乏。
烟青虫[Helicoverpaassulta(Guenée)]是一种寡食性昆虫,主要在茄科植物如烟草、辣椒和酸浆属(Physalis)的植物上取食。其幼虫嗜好取食烟草和其他寄主植物的花蕾等繁殖器官,造成比较严重的经济损失[13]。笔者以2种主要寄主作物辣椒和烟草作为供试植物,每种植物进一步分为营养器官和繁殖器官,将这些材料的乙醇提取物涂布在纱布上作为产卵基质,测试了从中孵化的烟青虫幼虫的嗅觉偏好性,旨在为透视经历在昆虫生活史中的功能提供参考。
1 材料与方法
1.1材料
1.1.1供试昆虫。烟青虫幼虫采集于郑州市毛庄绿色蔬菜基地的辣椒果实上,带回室内后放在指形管(10 cm×2 cm)中,用新鲜辣椒果实饲养(15 L∶9 D,光照期温度为30 ℃,暗期温度为28 ℃,相对湿度为60%~70%)。末龄幼虫转移到湿润洁净的细沙上化蛹。羽化后的成虫饲喂10%(W/V)的蔗糖溶液。 随后世代的幼虫用混入麦胚基人工饲料[14]饲养。室内种群用人工饲料纯化取食背景达到12个世代之后用于测试。
1.1.2供试植物。烟草[Nicotianatabacum(var.NC89)]和辣椒[Capsicumannum(var.Zhongshu No.6)]常规种植。测试用的株高约30 cm。4种植物材料(烟草花器、烟草叶片、辣椒果实和辣椒叶片)采集后放在隔离的自封袋中并带回实验室当天使用。
1.2方法
1.2.1卵期经历的诱导。供试材料用无菌水冲洗、晾干并称重。经过匀浆后加入与相应材料相等重量的无水乙醇,在室温下提取48 h之后过滤。棉纱布剪裁成约30 cm×30 cm的方块,分别用上述4种材料的乙醇提取物饱和(每块纱布大约可以吸收10 mL提取物),在通风橱中挥发溶剂20 min。无水乙醇浸渍并晾干处理的纱布作为对照组。上述5个处理组分别称为TL(烟叶粗提物经历组)、TF(烟草花器粗提物经历组)、HF(辣椒果实粗提物经历组)、HL(辣椒叶片粗提物经历组)和CK(对照组)。
试验时,将5对羽化后3 d的烟青虫蛾释放到锥形的塑料笼(上口直径20 cm,底部直径15 cm)中,迅速将各种粗提物或者无水乙醇处理过的棉纱布罩在上口,然后将笼子转移到和上述幼虫饲养条件相同的人工气候箱中。每天早上移除携带新产卵的纱布并更换相应处理的纱布,实时观察并分离出孵化的个体,立即进行嗅觉偏好性测试。
1.2.2嗅觉偏好性测试。嗅觉偏好性在4项选择的条件下测试。将滤纸剪成直径1.5 cm的小圆碟,取4个滤纸碟以“十”字交叉的方式放在1个底部灌装2%琼脂胶的培养皿(直径14 cm)中,分别在4个滤纸碟中央滴加40 μL上述4类提取物。培养皿底部灌装琼脂胶的目的是促进幼虫爬行选择,另外由于初孵幼虫取食量很小,故仅测试了嗅觉趋向反应。待溶剂挥发后,将来自特定经历诱导组的1头初孵幼虫释放到培养皿底部的中央,在均匀的光照下观察并记录幼虫首次做出的选择。5 min内未做出选择反应的个体放弃记录并测试下一头。为了避免孵化之后发生的渐进性学习现象,每头幼虫仅测试1次。不同气味源的位置随机化。每个处理组重复测试100头。
1.3统计分析采用χ2测验分析幼虫的选择频次。从4个不同角度进行统计分析:①整体差异显著性;②处理组之间选择趋势的差异;③相同处理组在不同气味源之间选择频次的差异;④每个处理组和无经历组之间的差异。为了鉴别出显著性的处理×气味源组合,进一步将测试数据分为6个亚组,每个亚组含有2个气味源及其相应的寄主组织粗提物处理组,采用χ2测验分辨卵期发育经历效应是正面的还是负面的。为了分析究竟是寄主种类的经历还是寄主植物组织的经历对初孵幼虫的寄主选择影响更大,相应的频次合并之后用χ2测验(针对组内差异)或χ2似然比测验(针对组间差异)。在上述分析中,凡是自由度只有1的案例,χ2测验采用Yates连续性校正法。
2 结果与分析
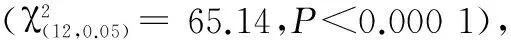
表1卵期经历不同寄主化学刺激的烟青虫初孵幼虫选择频次的组内差异与组间差异
Table1Within-andbetween-groupdifferencesamongchoicefrequenciesofH.assultaneonatesafterexperienceddifferenthostchemicalstimuliduringeggstage

卵期经历组Groups不同气味源被选择频次ChoicefrequencyofdifferentodorsourcesTFTLHFHLχ2P显著性SignificanceTF1418511736.40<0.0001***TL233121252.240.5241nsHF4015212413.680.0034**HL1615531641.84<0.0001***CK222026323.360.3394nsχ220.3612.0133.0511.6965.14P0.00040.0173<0.00010.0198<0.0001显著性Significance***********
注:TF.烟草花器;HF.辣椒果实;TL.烟草叶片;HL.辣椒叶片;ns.无显著性差异(P>0.05);*.0.01>P>0.05;**.0.01 >P>0.001;***.P<0.001
Note:TF.Tobacco flower;HF.Hot pepper fruit;TL.Tobacco leaf;HL.Hot pepper leaf.ns.No significant difference(P>0.05);*.0.01>P>0.05;**.0.01>P>0.001;***.P<0.001
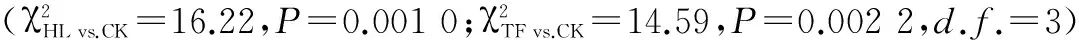
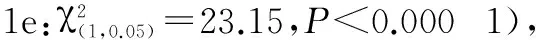
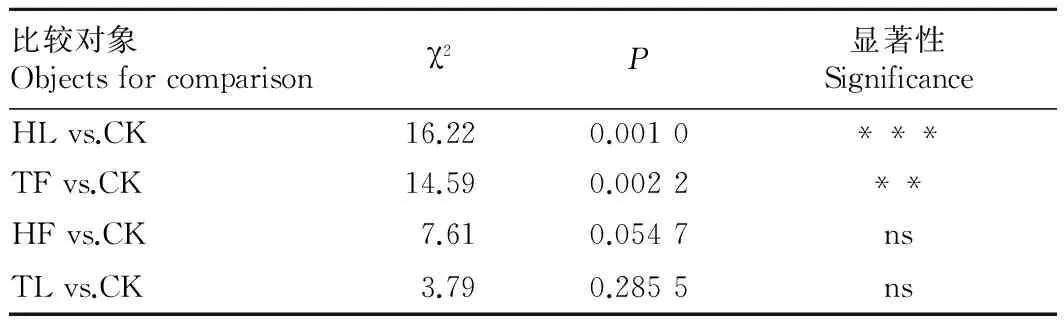
表2 各个经历组分别与对照组之间寄主选择反应的比较
注:TF.烟草花器;HF.辣椒果实;TL.烟草叶片;HL.辣椒叶片;ns.无显著性差异(P>0.05);*.0.01>P>0.05;**.0.01>P>0.001;***.P<0.001
Note:TF.Tobacco flower;HF.Hot pepper fruit;TL.Tobacco leaf;HL.Hot pepper leaf;ns.No significant difference(P>0.05);*.0.01>P>0.05;**.0.01>P>0.001;***.P<0.001
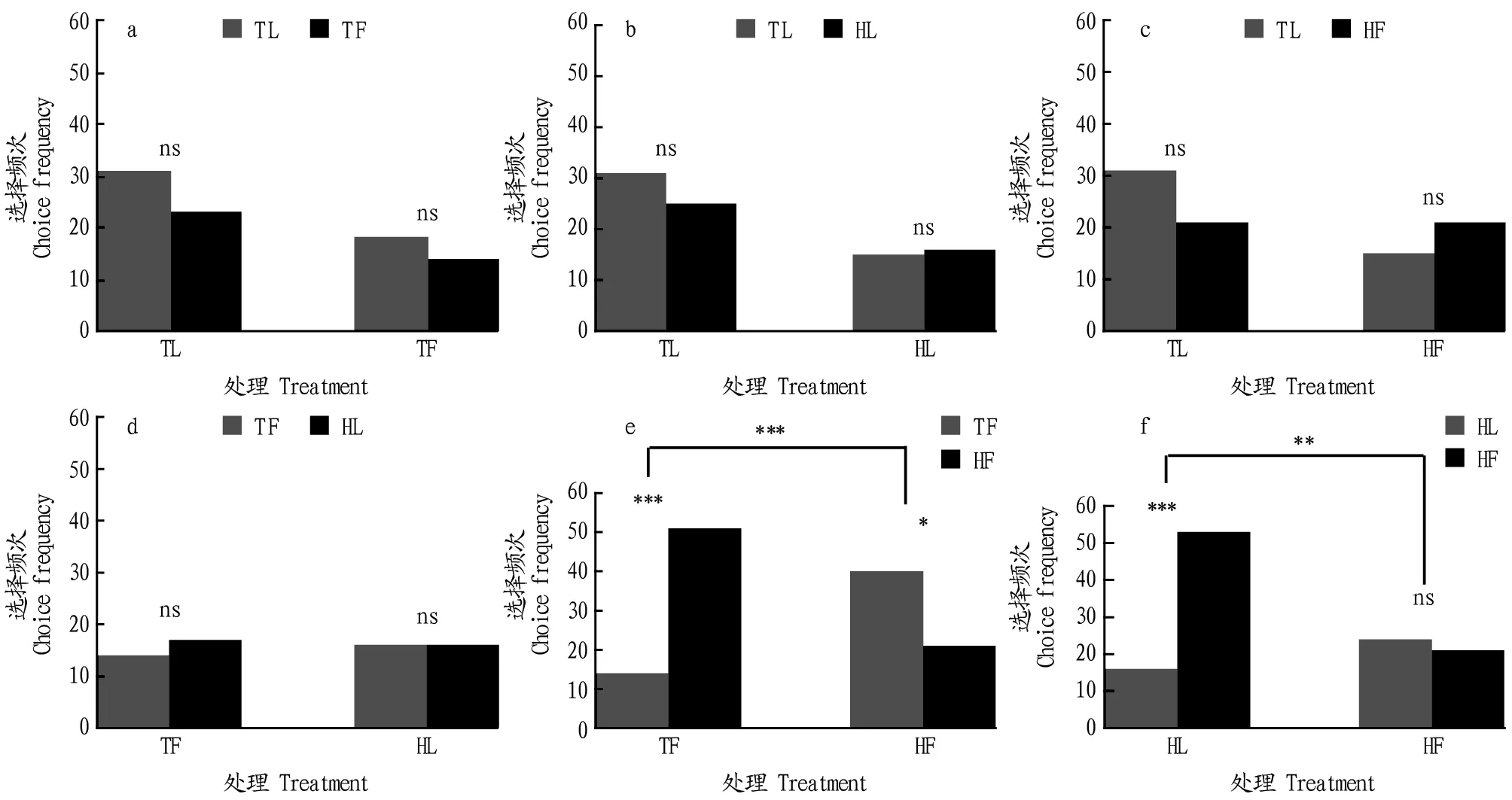
注:TF.烟草花器;HF.辣椒果实;TL.烟草叶片;HL.辣椒叶片;ns.无显著性差异(P>0.05);*.0.01>P>0.05;**.0.01>P>0.001;***.P<0.001Note:TF.Tobacco flower;HF.Hot pepper fruit;TL.Tobacco leaf;HL.Hot pepper leaf;ns.No significant difference(P>0.05);*.0.01>P>0.05;**.0.01>P>0.001;***.P<0.001图1 烟青虫初孵幼虫对有经历寄主和与其配对的无经历寄主选择频次比较Fig.1 Choice frequencies of H.assulta neonates with different egg experiences to selected paired options with corresponding egg experiences

3 结论与讨论
在影响植食性昆虫初孵幼虫嗅觉偏好性的因素中,卵期经历的寄主化学刺激物对随后寄主选择影响的信息相对比较缺乏,尤其是不了解寄主种类、寄主植物器官或者是二者特定的组合经历效应的大小。该研究发现,烟青虫初孵幼虫的嗅觉偏好性受卵期发育环境的显著影响,但这些幼虫并非对经历过的寄主植物或者其组织嗅觉偏好性增强。
在4类测试材料中,只有卵期经历过来自辣椒叶片和烟草花器的化学刺激物,初孵幼虫的嗅觉偏好性才和对照组相比有显著差异。这些结果恰好和自然雌虫的产卵位置选择一致。Han等[15]在辣椒田调查了不同器官上烟青虫的落卵量,发现叶片上的产卵比例占76.2%,而辣椒果实、茎干和花上的产卵百分比仅是16.8%、6.5%和0.4%。然而在烟草上,开花期是对烟青虫雌蛾产卵引诱力最强的生育期[13]。田间调查表明,6月和7月烟草花器上的烟青虫卵所占的比例分别为88.71%和94.44%[16]。
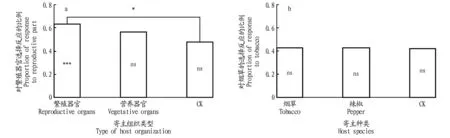
注:ns.无显著性差异(P>0.05);*.0.01>P>0.05;***.P<0.001Note:ns.No significant difference(P>0.05);*.0.01>P>0.05;***.P<0.001图2 寄主种类或者寄主组织类型合并之后的经历效应分析Fig.2 Experience effect after host species or part types pooled
卵期经历过辣椒叶片或烟草花器化学刺激的初孵幼虫,总是诱导出对辣椒果实气味源的嗅觉偏好性,但是卵期经历过辣椒果实化学刺激的初孵幼虫却并没有对其嗅觉偏好性增强。也就是说,卵期经历的效应并不是正面的,这和此前的多数报道是相反的[17-18]。当前仅报道过东方云杉卷叶蛾(Choristoneurafumiferana)有类似的现象,雌成虫偏好在幼虫期没有取食经历的寄主植物上产卵[19]。然而,尽管该研究结果也发现经历的效应是负面的,但供试对象却是不食不动的卵。
该研究发现,卵发育期间经历的寄主植物组织而不是寄主植物的种类,对初孵幼虫的嗅觉偏好性才有显著的影响。尤其明显的是,卵期经历过繁殖器官化学刺激的初孵幼虫,对繁殖器官的嗅觉偏好性非常强。这可以用化学遗产假说来解释,而不是霍普金斯寄主选择原理,因为卵期并不会摄入食物,来自卵发育环境的痕量化学物质可能存在于初孵幼虫的血淋巴中或者在卵壳之外。后续研究需了解寄主繁殖器官中究竟是哪些活性成分诱导了这种嗅觉偏好性。
总之,烟青虫卵期经历过烟草花器或者辣椒叶片的化学刺激,会导致初孵幼虫嗅觉偏好性的变化,二者也正是自然雌蛾偏好产卵的部位。卵发育期间经历过繁殖器官化学刺激的烟青虫初孵幼虫对繁殖器官的嗅觉偏好性明显增强。尽管烟青虫的卵产在烟草花器和辣椒叶片上,但是幼虫可以自行运动到繁殖器官上,为该害虫卵与幼虫分布的不一致性提供了一个崭新的解释。
[1] WOODS H A.Water loss and gas exchange by eggs ofManducasexta:Trading off costs and benefits[J].Journal of insect physiology,2010,56(5):480-487.
[2] ZALUCKI M P,CLARKE A R,MALCOLM S B.Ecology and behavior of first instar larval Lepidoptera[J].Annu Rev Entomol,2002,47(1):361-393.
[3] PERKINS L E,CRIBB B W,HANAN J,et al.Where to from here? The mechanisms enabling the movement of first instar caterpillars on whole plants usingHelicoverpaarmigera[J].Arthropod-plant interact,2008,2:197-207.
[4] SADEK M M.Complementary behaviors of maternal and offspringSpodopteralittoralis:Oviposition site selection and larval movement together maximize performance[J].J Insect Behav,2011,24(1):67-82.
[5] STAMPS J A,DAVIS J M.Adaptive effects of natal experience on habitat selection by dispersers[J].Anim Behav,2006,72(6):1279-1289.
[6] KAROWE D N.Facultative monophagy as a consequence of prior feeding experience:Behavioral and physiological specialization inColiasphilodicelarvae[J].Oecologia,1989,78(1):106-111.
[7] RENWICK J A A.Variable diets and changing taste in plant-insect relationship[J].J Chem Ecol,2001,27(6):1063-1076.
[8] AKHTAR Y,ISMAN M B.Larval exposure to oviposition deterrents alters subsequent oviposition behavior in generalist,Trichoplusianiand specialist,Plutellaxylostellamoths[J].J Chem Ecol,2003,29(8):1853-1870.
[9] CHOW J K,AKHTAR Y,ISMAN M B.The effects of larval experience with a complex plant latex on subsequent feeding and oviposition by the cabbage looper moth:Trichoplusiani[J].Chemoecology,2005,15(3):129-133.
[10] WANG H,GUO W F,ZHANG P J,et al.Experience-induced habituation and preference towards non-host plant odors in ovipositing females of a moth[J].J Chem Ecol,2008,34:330-338.
[11] SHIKANO I,ISMAN M B.A sensitive period for larval gustatory learning influences subsequent oviposition choice by the cabbage looper moth[J].Anim Behav,2009,77(1):247-251.
[12] ANDERSON P,SADEK M M,LARSSON M,et al.Larval host plant experience modulates both mate finding and oviposition choice in a moth[J].Anim Behav,2013,85(6):1169-1175.
[13] SUN J G,HUANG L Q,WANG C Z.Electrophysiological and behavioral responses ofHelicoverpaassulta(Lepidoptera:Noctuidae)to tobacco volatiles[J].Arthropod-plant interact,2012,6(3):375-384.
[14] AHN S J,BADENES-PÉREZ F R,HECKEL D G.A host-plant specialist,Helicoverpaassulta,is more tolerant to capsaicin fromCapsicumannuumthan other noctuid species[J].J Insect Physiol,2011,57(9):1212-1219.
[15] HAN M W,LEE J H,SON J S.Intra-and inter-plant distribution ofHelicoverpaassulta(Lepidoptera:Noctuidae)eggs in red pepper and tobacco fields[J].Korean J Appl Entomol,1994,33:6-11.
[16] 郭线茹,罗梅浩,马继盛.烟青虫/棉铃虫混合种群卵在烟田的空间分布特征研究.Ⅰ.混合种群卵在春烟田的空间分布及其密度估测技术[J].中国烟草学报,1995(2):1-7.
[17] ANDERSON P,ANTON S.Experience-based modulation of behavioural responses to plant volatiles and other sensory cues in insect herbivores[J].Plant,Cell & Environ,2014,37(8):1826-1835.
[18] DAVIS J M.Patterns of variation in the influence of natal experience on habitat choice[J].The quarterly review of biology,2008,83(4):363-380.
[19] MADER B J,DAOUST S P,CARDINAL-AUCOIN M,et al.Larval experience induces adult aversion to rearing host plants:A novel behaviour contrary to Hopkins’ host selection principle[J].Ecol Entomol,2012,37(3):204-211.
猜你喜欢
世界竹藤通讯(2022年2期)2022-05-07
农业科技通讯(2022年3期)2022-03-26
植物保护(2022年1期)2022-02-10
紫禁城(2020年9期)2020-10-27
紫禁城(2020年9期)2020-10-27
紫禁城(2020年9期)2020-10-27
现代农村科技(2018年3期)2018-04-02
四川农业科技(2015年5期)2015-04-04
湖南中医药大学学报(2011年11期)2011-03-20
