
海拔梯度对西藏高原3种禾本科牧草种子大小和萌发特性的影响
2018-04-09 03:27,,,,
种子 2018年2期
, , , ,
(西藏自治区农科院草业科学研究所, 拉萨 850009)
青藏高原有世界“第三极”之称,其高寒草原生态系统极其脆弱,正面临着草地退化的危机[1]。时下,高原植被恢复已成为社会各界人士关注的问题之一。西藏野生牧草种质资源丰富,全区可食牧草种类有2 672种[2],其中高寒草原以禾本科早熟禾属(Poa)、披碱草属(Elymus)、针茅属(Stipa)为主要的建群植物,其拥有较强的适应性和高度的抗逆性等优质性状,是畜牧业发展和退化草地恢复和重建的关键。
表1种子采集地概况

植物名称编号北纬(N)东经(E)海拔(m)年均温度(℃)采集地垂穗披碱草ElymusnutansE⁃131°51′94°24′42001.4那曲地区巴青县雅安镇E⁃231°08′92°12′4566-1.9那曲县E⁃331°24′90°01′4700-1.2班戈县冷地早熟禾PoacrymophilaP⁃129°53′92°30′4234-3.8林芝地区工布江达县江达乡P⁃231°08′92°12′4566-2.2那曲县P⁃332°03′92°17′4848-10.0聂荣县聂荣镇5村丝颖针茅StipacapillaceaS⁃130°06′92°48′4045-3.8林芝地区工布江达县江达乡S⁃231°51′94°24′42001.4那曲地区巴青县雅安镇S⁃330°35′91°18′43601.3拉萨市当雄县龙仁乡
表2不同海拔3种植物发芽特性变化

植物名称编号千粒重(g)发芽率(%)发芽速率胚芽(cm)胚根(cm)披碱草ElymusnutansE⁃14.6±0.04a88.2±4.81a18.1±0.77a11.3±0.417.8±0.29E⁃23.8±0.10b75.6±3.75a14.2±0.84b9.8±0.297.9±0.35E⁃33.1±0.05c54.7±4.00b7.5±1.04c7.5±0.455.3±0.72冷地早熟禾PoacrymophilaP⁃10.18±0.005b59.8±3.46b10.8±0.70b2.4±0.142.8±0.35P⁃20.21±0.003a93.4±1.33a13.7±0.94a2.5±0.073.9±0.29P⁃30.11±0.002c49.3±5.81b7.7±0.76c2.2±0.103.6±0.39丝颖针茅StipacapillaceaS⁃17.5±0.19a84.0±4.06a15.9±0.65a6.4±0.183.8±0.16S⁃26.5±0.07b59.6±4.62b8.3±0.90b5.6±0.254.4±0.35S⁃33.3±0.08c70.7±4.81ab11.0±0.94b4.9±0.263.9±0.22
注:相同牧草不同海拔间做单因素方差分析,不同字母表示Duncan方差分析后的显著性差异。
种子萌发是植物生命活动过程中重要的环节,是新生命开始植物定居和种群延续的前提[3]。直接关系着物种维持、繁衍、扩散等生态过程。研究发现,许多植物种子萌发与其生长的地理环境的异质性有密切关系。如多年生草本植物种子萌发随海拔升高而降低[4]。也有研究指出,种子萌发率与海拔呈显著的负相关[5]。西藏高原海拔落差极大,不同海拔种子差异性对植物种子萌发的影响有待研究。本试验以西藏野生禾本科植物冷地早熟禾(Poacrymophila)、垂穗披碱草(ElymusnutansGriseb)、丝颖针茅为(StipacapillaceaKeng)材料,研究不同海拔牧草种子大小和萌发特性的差异,揭示高海拔条件下种子生存和萌发对策,旨在为西藏畜牧业生产和草地恢复提供优质种质资源。
1 材料与方法
1.1 材 料
不同海拔的牧草种子于2013年9月中旬在西藏自治区拉萨市、那曲地区、林芝地区境内采集(见表1)。为避免相同植株种子对发芽的干扰,将采集后的50株相同种群不同株丛的种子充分混合,自然风干。种子清选后置于牛皮纸信封中室温条件下贮藏备用。次年4月挑选成熟、均匀一致的种子进行萌发试验。
1.2 方 法
1.2.1种子千粒重测定
随机选取清选好的100粒种子,称百粒重,精确到0.000 1 g,重复5次,取其平均值,计算千粒重[6]。
1.2.2萌发试验
萌发试验于拉萨市农科院草业所实验室内进行。将不同种类和不同海拔的种子均匀排列于发芽床上,发芽床为垫有双层滤纸的直径为9 cm的培养皿,滤纸始终保持湿润而无明水,每皿放置100粒种子,3个重复。将装有发芽种子的培养皿放于25 ℃恒温,光照16 h/黑暗8 h的培养箱中。萌发试验期间,每天补充因蒸发而散失的水分,以尽量减少水势变动。每天统计发芽数(胚根长度达到与种子等长、胚芽长度达到种子一半作为发芽标准)[7],第14天后发芽结束。测定根长和芽长并计算发芽率、发芽速率(GS)。
发芽率(%)=发芽种子数/供试种子总数×100%;
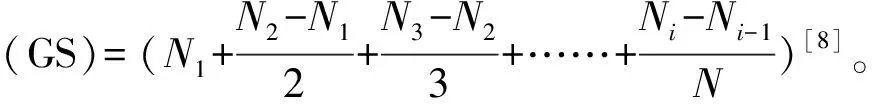
1.3 数据分析
试验数据采用Excel与SPSS 20.0软件进行方差分析,用Duncan法进行多重比较。
2 结果与分析
2.1 不同海拔3种植物发芽特性变化
2.1.1千粒重差异
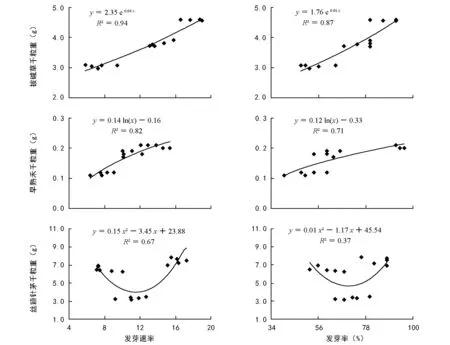
图1 种子大小与发芽率、发芽速率的回归分析
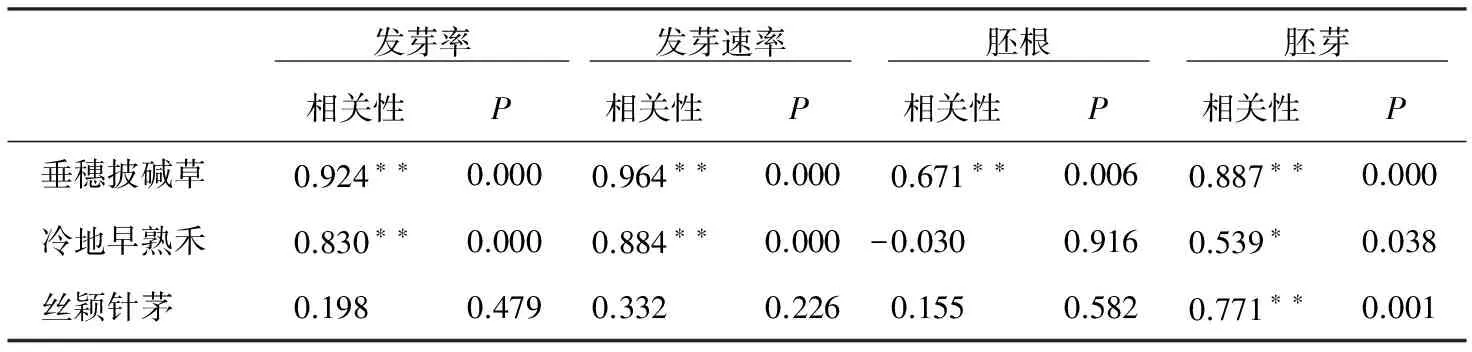
表3种子大小和发芽率、发芽速率、根长和芽长的相关性
发芽率 发芽速率 胚根 胚芽 相关性P相关性P相关性P相关性P垂穗披碱草0.924∗∗0.0000.964∗∗0.0000.671∗∗0.0060.887∗∗0.000冷地早熟禾0.830∗∗0.0000.884∗∗0.000-0.0300.9160.539∗0.038丝颖针茅0.1980.4790.3320.2260.1550.5820.771∗∗0.001
注:**(*)代表0.01(0.05)水平下相关显著性检验。
不同海拔野生垂穗披碱草、高原早熟禾、丝颖针茅种子的千粒重差异显著(p<0.05),如表2。垂穗披碱草、冷地早熟禾和丝颖针茅种子千粒重分别为3.05~4.58 g、0.11~0.21 g和3.34~7.46 g。垂穗披碱草和丝颖针茅种子千粒重随海拔升高而降低,早熟禾以P-2(海拔4 654 m)种子的千粒重最大,P-3(海拔4 848 m)最小。
2.1.2发芽率、发芽速率的变化
表2所示,不同海拔禾本科植物种子发芽率都有明显差异,垂穗披碱草、冷地早熟禾和丝颖针茅发芽变幅分别为54.7%~88.0%、49.3%~93.4%、60.0%~84.0%。披碱草发芽率随海拔升高而降低;早熟禾呈现单峰变化,P-2发芽率最高,达93.4%,是P-3发芽率的2倍;丝颖针茅在巴青县雅安镇(S-2)发芽率最低,为60.0%。相同牧草不同海拔间单因素方差分析表明:班戈县披碱草(E-3)的显著低于那曲县(E-1)和巴青县雅安镇(E-2);P-2显著高于P-1和P-3(p<0.05),工布江达县丝颖针茅(S-1)显著高于S-2(p<0.05)。
不同禾本科植物发芽速率随海拔的变化如表2所示,发芽速率与发芽率的变化趋势一致,单因素方差分析结果显示,不同海拔垂穗披碱草、早熟禾的发芽速率差异均显著(p<0.05)。S-1显著高于S-2和S-3(p<0.05)。
2.1.3胚根长和胚芽长的变化
由表2可见,牧草种子胚根和胚芽长度差异明显,垂穗披碱草随海拔升高胚根、胚芽长度缩短,相同海拔其胚芽长度>胚根长度,那曲县冷地早熟禾其胚根和胚芽最长,同一海拔其胚根长度>胚芽长度,丝颖针茅胚芽随海拔升高长度缩短。同一海拔其胚芽长>胚根长。
2.2 种子大小和发芽率、发芽速率、根长和芽长的相关性
不同海拔3种禾本科牧草种子大小与发芽率、发芽速率、胚根长和胚芽长的相关关系如表3所示,垂穗披碱草的种子大小与发芽率、胚根长和胚芽长极显著正相关(p<0.01),与发芽速率极显著负相关(p<0.01)、相关系数均在0.67以上。冷地早熟禾的种子大小与发芽率、发芽速率极显著正相关(p<0.01),与胚芽长显著相关(p<0.05)。丝颖针茅种子大小与胚芽长极显著正相关(p<0.01)。
2.3 种子大小与发芽率、发芽速率的回归分析
对不同海拔植物种子大小与发芽率、发芽速率进行回归拟合,结果见图1,垂穗披碱草种子大小与发芽速率、发芽率成指数拟合较好。拟合方程分别为y=2.35 e0.04 x、y=1.76 e0.01 x,决定系数0.94和0.87;冷地早熟禾种子大小与发芽速率、发芽率对数拟合较优,决定系数分别为0.82和0.71;丝颖针茅种子大小与发芽速率、发芽率呈2次拟合关系,拟合系数较低。
3 讨论与结论
植物种子在长期的进化过程中形成了一系列的适应生存环境的对策,种子大小的变异受植物遗传进化和自然选择的共同作用,系统发育是种子大小变异的内因,环境条件是其变化的外因。海拔是一个复合的环境梯度,造就了不同的水、气、热、光照和土壤营养的梯度,成为影响种子大小变异的重要因素之一。本研究中随海拔升高供试种子千粒重随之降低,这与郭淑青和杨霞等研究结果类似[9-10]。在高原地区,尤其是海拔4 000 m以上的区域,随着海拔的升高,植物生长环境条件严酷而多变,种子趋于微小化,王琳[11]等在研究古尔班通古特沙漠种子大小空间变异中提出,受沙漠极端环境的影响,植物更倾向产生小质量种子。这类种子具有较远的传播距离,能充分利用环境中的随机降水,使种群能够延续下来,也充分印证了本研究的结论。
种子萌发直接体现了植物的总体适应性。影响着幼苗的存活及个体适合度[12]。同种植物生长在不同海拔梯度下其萌发特性也不近相同,可能与生境气候条件存在差异有关[13],为了适应不同海拔多变的环境,每一植物都有各自的生存策略。西藏高原不同地区气候差异显著,年均降水变幅也较大[14],聂荣县年均温度仅为-10 ℃,而工布江达县年均温度为1.4 ℃。因此,本研究不同地区相同的垂穗披碱草、早熟禾和丝颖针茅其萌发特性是受各地气候条件决定的。
海拔决定的种子大小能够影响种子萌发能力[15],从而影响群落植物更新。到目前为止,在关于种子大小与萌发特性关系的研究中,并未得出一致性的结论。许静等[5]对高寒草甸32科524种植物种子的萌发研究,发现种子大小与萌发率显著相关,质量小的种子具有相对较高的萌发率和较快的萌发速度。Stanton[16]认为,较大种子萌发明显快于小种子,种子大小和种子的萌发率、萌发速率呈明显的负相关关系。本研究结果显示,垂穗披碱草大种子比小种子有着较高的发芽率、发芽速率、胚根长和胚芽长,早熟禾种子大小与发芽率、发芽速率和胚芽长正相关。丝颖针茅种子大小与萌发特性相关性不明显,高海拔小种子的萌发率不高。分析原因,西藏高寒草原区,高海拔与低海拔相比,年均温较低、生长季短,霜冻等恶劣的自然天气频繁,这将对种子的萌发产生不利的影响。种子为了适应环境,质量变小。小种子在高海拔生境下可能需要较长的寒冷需求[17],也会影响种子的萌发。
由此可见,种子的萌发是多变的,在不同海拔及温、湿度条件下同种植物或不同植物有着不同的萌发特性。所以关于植物种子生物学特征和种子萌发的研究,还应从发育学,生态学各个角度进行系统分析研究。
参考文献:
[1]鱼小军,徐长林,景媛媛,等.冬季层积处理对5种高寒草甸植物种子萌发特性的影响[J].草业科学,2015,32(3):427-432.
[2]边巴卓玛,呼天明,吴红新.依靠西藏野生牧草种质资源提高天然草场的植被恢复效率[J].草业科学,2006,23(2):6-8.
[3]崔现亮,罗娅婷,毕廷菊,等.储藏和萌发温度对青藏高原东缘12种灌木种子萌发的影响[J].生态学杂志,2014,33(1):23-32.
[4]王桔红,张勇,崔现亮,等.不同海拔梯度糙皮桦和紫果云杉种子的萌发变异[J].生态学杂志,2009,28(4):589-594.
[5]许静,杜国祯,李文龙,等.温度和海拔对高寒草甸植物种子萌发进化特性的影响[J].兰州大学学报(自然科学版),2013,49(3):377-383.
[6]Zhang S T,Du G Z,Chen J K.Correlates of seed size in a subalpine meadow on the east of the Tibetan plateau[J].Eco-science,2004,11:6-15.
[7]GB/T 2930.4—2001.牧草种子检验规程[S].北京:中国标准出版社,2001:3-4.
[8]Yu J B,Wang X H,Ning K,et al.Effects of salinity and water depth on germination of Phragmites australis in Goastal Wetland of the Yellow River Delta[J].Clean-Soil,Air,Water,2012,4040(10):1 154-1 158.
[9]郭淑青,齐威,王玉林,等.青藏高原东缘海拔对植物种子大小的影响[J].草业学报,2010,19(1):50-58.
[10]杨霞,梁艳,陈学林.青藏高原东缘地区常见植物种子大小变异研究[J].生态科学,2007,26(6):483-489.
[11]王琳,刘彤,韩志全,等.古尔班通古特沙漠植物种子大小变异的空间格局[J].生态学报,2014,34(23):6 797-6 806.
[12]Donohue K,Dorn L,Griffith C,et al.The evolutionary ecology of seed germination of Arabidopsis thaliana:Variable natural selection on germination timing[J].Evolution,2005,59:758-770.
[13]刘志民,李雪华,李荣平,等.科尔沁沙地15种禾本科植物种子萌发特性比较[J].应用生态学报,2003,14(9):1 416-1 420.
[14]朱文泉,高清竹,段敏捷,等.藏西北高寒草原生态资产价值评估[J].自然资源学报,2011,26(3):419-428.
[15]张蕾,张春辉,吕俊平,等,青藏高原东缘31种常见杂草种子萌发特性及其与种子大小的关系[J].生态学杂志,2011,30(10):2 115-2 121.
[16]Stanton M L.Seed variation in wild radish:Effect of seed size on components of seeding and adult fitness[J].Ecology,1984,65:1 105-1 112.
[17]崔现亮,王桔红,齐威,等.青藏高原东缘灌木种子的萌发特性[J].生态学报,2008,28(11):5 294-5 301.
猜你喜欢
安徽农业科学(2021年14期)2021-09-03
山西农业科学(2021年8期)2021-08-17
青年文学家(2021年4期)2021-03-18
农民致富之友(2020年19期)2020-07-18
山西农业科学(2020年1期)2020-02-27
学苑创造·A版(2019年11期)2019-12-05
家庭百事通·健康一点通(2019年11期)2019-09-26
小学生学习指导(低年级)(2018年9期)2018-09-26
读者·校园版(2018年1期)2017-12-16
少年科学(2015年2期)2015-08-12
