
不同树种与土地利用方式对土壤微生物生物量碳氮的影响
2018-04-09 02:08徐建峰赵家豪袁在翔金雪梅关庆伟
中南林业科技大学学报 2018年4期
徐建峰 ,赵家豪 ,袁在翔 ,金雪梅 ,陈 斌 ,关庆伟 ,高 洋
(1.江苏省太仓市林业站,江苏 太仓 215400;2.南京林业大学 生物与环境学院 南京林业大学南方现代林业协同创新中心,江苏 南京 210037)
土壤微生物量是指在土壤中体积小于5×103μm3的生物总量[1-3]。微生物生物量碳氮是其重要的表征指标。虽然微生物生物量碳氮仅占土壤有机碳1%~4%[4]、全氮0.5%~8.0%[5],但却是土壤中最活跃的碳氮组分,直接参与土壤碳氮矿化,调控陆地生态系统碳氮循环。此外,土壤微生物生物量碳氮受气候、土壤、植被和土地利用方式等因子影响显著[2-3],是土壤生态系统变化的敏感指标[6-7]。有研究表明,即使在相同的立地条件下,不同植被类型下土壤微生物生物量碳氮差异显著[3,8];并且当土地利用方式发生变化时,土壤微生物生物量碳的变化更加迅速,短期内(3~5 a)内就可以检测到,5~10 a就可达到新的平衡状态[2]。
近年来,国内外关于植被、土地利用方式对土壤微生物生物量碳氮的影响已有不少研究。在植被方面主要集中在森林类型[3,9-11]、植物群落[12]、林龄[13-14]、凋落物[15]、根系分泌物[16]、植被恢复模式[17]对土壤微生物生物量的影响,对于相同立地条件、相近林分特征及经营历史下因树种差异对土壤微生物生物量碳氮影响的研究相对较少;在土地利用方式方面主要集中在耕作方式[18-20]、退耕还林[21]对土壤微生物生物量碳氮的影响,而在对退耕还林的比较中弱化了树种差异。因此,本研究通过比较近同质条件下3个树种纯林及农田之间不同土壤层次微生物生物量碳氮含量的差异,旨在阐明树种差异与土地利用方式转变对土壤微生物量的影响,为造林树种选择提供基础数据。
1 研究地概况
试验地位于江苏省太仓市S339省道两侧的绿色通道内(121°31′24″E,31°26′39″N),属长江三角洲冲积平原。气候为北亚热带湿润季风气候,四季分明,年平均气温15.5 ℃,降水量1 078.1 mm,日照1 960.9 h,无霜期226 d。土壤类型为水稻土,平均土层厚度70 ~ 80 cm,中性至偏碱性。地下水位较高。林分类型均为人工纯林,林龄为10 a,林下鲜有植被。
2 研究方法
2.1 样地设置与试验设计
试验地在造林之前一直为水稻田,地势平坦均一,并具有相同的栽培耕作历史。3年生幼树于2008年均以3 m×3 m的种植密度栽植在近似等长等宽的矩形区域内(由太仓市农林局提供相关资料),并于2015年4月份进行了一次除草抚育措施,因此近似于同质试验区。此次研究根据实地踏勘情况选取了3段互不相邻且间隔100 m(即一个非试验林分长度)左右,规格大小皆为100 m×30 m的纯林。各纯林分别栽植水杉、香樟、重阳木,林分基本概况见表1。在100 m×30 m的纯林内随机设置3个20 m×20 m标准采样区,并且远离林分边界。对照农田相距试验林分较近,且标准采样区的设置与上述林分相同。
2015年11月中旬在每块20 m×20 m标准采样地内,除去地面凋落物和草本植物,按“S”型选7个点,为了减少浅表性地下水对土壤的非均质影响,故用土钻取0~10 cm、10~20 cm、20~40 cm以下共3层土样,每层取样点土样混合后作为该层待测土样。同时,挖取土壤剖面采用环刀法测定各土层容重。土样经冷藏箱带回实验室后分成2份,一份新鲜去杂、过2 mm钢筛后贮藏于4 ℃的冰箱内,并在一周内完成土壤微生物生物量碳(SMBC)、微生物生物量氮(SMBN)的测定,另一份自然风干、去杂、过筛后进行土壤全氮、有机碳、pH值等其他指标的测定

表1 三个树种纯林斑块基本概况Table 1 Site characteristics in the monocultures of three tree species
2.2 试验指标测定
微生物生物量碳(氮):采用氯仿熏蒸—K2SO4浸提法[22],浸提液经0.45 μm滤膜过滤用TOC—VCPN自动分析仪(岛津,日本)测定土壤提取碳(氮)。微生物生物量碳=(熏蒸后土壤提取碳-未熏蒸土壤提取碳)/ 0.45[15],微生物生物量氮=(熏蒸后土壤提取氮-未熏蒸土壤提取氮)/ 0.57[23]。
土壤全氮、有机碳:由于试验土壤样品偏碱性,故采用1 mol/L HCl充分去除风干土样中的无机碳,烘干并研磨过0.149 mm钢筛后用元素分析仪(Elementar Vario EL,德国)测定[24-25]。pH用电位法测定(水∶土 = 2.5∶1);土壤容重用环刀法测定;湿度用烘干法测定。
2.3 数据分析
用Excel 2003、Origin 8.5和SPSS 19.0统计分析软件进行数据分析和图表处理。其中,土壤微生物生物量碳氮与相关理化指标均进行One-way ANOVA 方差分析,LSD多重比较法进行差异显著性分析,Pearson相关系数评价土壤微生物生物量碳氮与相关因子之间的关系。
3 结果与分析
3.1 不同树种纯林及土地利用方式土壤理化性质比较
由表2可知,不同树种纯林土壤理化性质存在一定的差异,其中碳氮等相关指标差异性显著(P<0.05)。具体表现在,3个土壤层次的容重、pH等均值表现为:水杉>香樟>重阳木,但差异不显著;土壤含水率在18.78%~23.77%之间波动,0~10 cm土层无明显差异,10~20 cm、20~40 cm土层重阳木显著高于其他2个树种;土壤有机碳与全氮在重阳木纯林各土层中含量均为最高,且与其余的2树种均存在显著差异(P<0.05),水杉与香樟之间仅20~40 cm土层全氮含量存在明显的差异(P<0.05);各树种纯林内土壤容重、pH等指标在不同土壤层次之间差异性不显著,土壤含水率、有机碳、全氮等指标随土层加深而降低,其中表层土壤(0~10 cm)显著高于下层土壤(10~20 cm、20~40 cm,P<0.05)。
农田土壤容重变化范围为1.30~1.43 g·cm-3、pH为7.40~7.57,均与林地之间无显著差异;土壤含水率除0~10 cm表层土壤与林地差异不显著外,其余土层均显著高于3种林地(P<0.05);土壤有机碳与全氮显著高于林地,其中与水杉、香樟纯林之间差值最大。土壤容重、pH、含水率等指标随土层深度变化差异性不显著,土壤有机碳、全氮含量在0~10 cm与10~20 cm土层之间无明显差异,但都显著高于20~40 cm土层(P<0.05)。
3.2 不同树种纯林及土地利用方式微生物生物量碳、氮含量比较
由图1a可知,3个树种纯林土壤微生物生物量碳含量在174.22~373.34 mg·kg-1之间波动,其中重阳木纯林各土壤层次微生物生物量碳含量均显著高于水杉、香樟纯林(P<0.05),水杉纯林各土壤层次微生物生物量碳含量均值与香樟略有差异,但不显著。各树种纯林土壤微生物生物量碳的平均含量均随土层深度增加均呈现“高低高”的变化特征。其具体表现为,水杉0~10 cm、20~40 cm土层微生物生物量碳含量显著高于10~20 cm土层(P<0.05),0~10 cm与20~40 cm土层之间无显著差异;香樟各土壤层次之间均无显著差异;重阳木表层土壤(0~10 cm)显著高于下层土壤(10~20 cm、20~40 cm,P<0.05)。
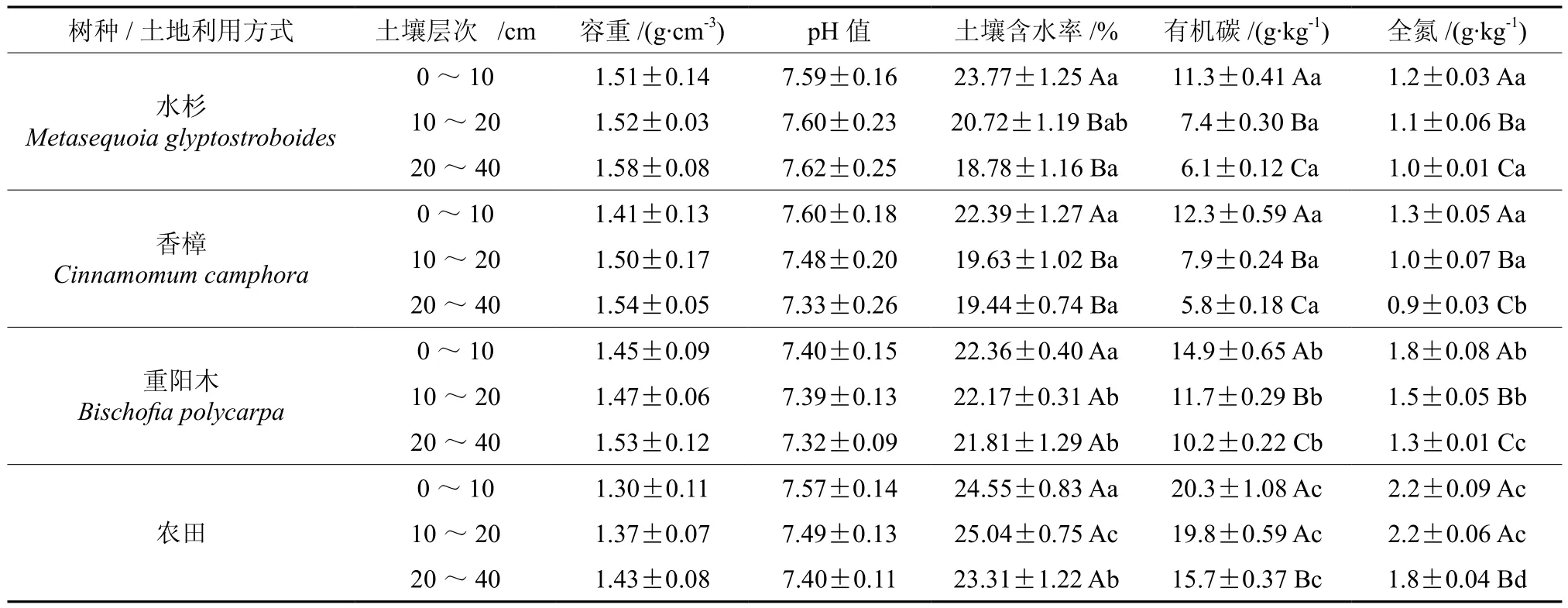
表2 不同树种纯林及土地利用方式土壤理化性质(平均值±标准误)†Table 2 Soil physicochemical properties in the monocultures of three tree species and landuse patterns (mean±SE)
如图1b所示,树种显著影响土壤微生物生物氮含量(P<0.05)。0~10 cm土壤微生物生物量氮含量变化范围在11.00~19.27 mg·kg-1之间,大小顺序为:重阳木>水杉>香樟;10~20 cm土壤微生物生物量氮含量波动范围为5.87~9.29 mg·kg-1,表现为重阳木>香樟>水杉;20~40 cm土层为8.28~11.79 mg·kg-1,与0~10 cm土壤微生物生物量氮含量的大小顺序相同。3树种纯林土壤微生物生物量氮平均含量随土层深度的变化趋势与微生物生物量碳相同,表现出“高-低-高”的变化特征。其中表层土壤微生物生物量氮含量(0~10 cm)显著高于下层(10~20 cm、20~40 cm,P<0.05)。

图1 不同树种纯林及土地利用方式土壤微生物生物量碳、氮含量比较Fig.1 Comparisons of soil microbial biomass carbon & nitrogen content in the monocultures of three tree species and landuse patterns
农田土壤微生物生物量碳含量在278.22~359.11 mg·kg-1之间,微生物生物量氮含量在6.80~29.48 mg·kg-1之间,除0~10 cm微生物生物量碳含量与重阳木纯林之间差异不显著外,其余的均与林地微生物生物量碳氮含量之间存在显著差异(图1,P<0.05)。具体表现为,农田土壤微生物生物量碳显著高于水杉、香樟纯林(P<0.05),表层含量与重阳木相近,中层含量显著高于重阳木,而底层含量则明显低于重阳木(P<0.05);微生物生物量氮含量0~10 cm、10~20 cm土层显著高于林地,20~40 cm土层则显著低于林地。微生物生物量碳氮平均含量随土层深度的变化趋势与林地显著不同,呈现出“低-高-低”的变化特征,其中0~10 cm、10~20 cm土层显著高于20~40 cm土层(P<0.05),而0~10 cm与10~20 cm土层之间无显著性差异。
3.3 土壤微生物生物量碳氮与相关因子之间的相关性分析
由表3可知,土壤微生物生物量碳氮含量之间存在极显著正相关关系(P<0.01),与土壤容重之间呈负相关关系,与土壤含水率、有机碳、全氮之间呈正相关关系,显著性因树种与土地利用方式的不同而有所差异。具体表现为,水杉除土壤微生物生物量碳氮含量之间极显著正相关外,仅土壤微生物生物量氮含量与土壤总有机碳含量之间显著正相关(P<0.05);香樟与农田除土壤微生物生物量氮含量与容重、pH值之间相关性不显著外,其余的均呈显著或极显著正负相关关系;重阳木纯林除土壤微生物生物量碳氮含量与容重、pH值、含水率之间相关性不显著外,其余均呈显著或极显著正相关关系。
4 结论与讨论
4.1 树种差异对土壤微生物生物量碳氮含量的影响
不同树种土壤微生物生物量碳氮平均含量存在一定差异,其中微生物生物量氮含量随树种变化差异性显著。重阳木各土层微生物生物量碳氮含量均显著高于其他树种,主要原因可能是重阳木纯林土壤容重较小,通气性较好,且有机质、全氮、水分含量较高(表2),凋落物分解速度快[26],有利于土壤微生物的生长与繁殖。水杉纯林各土层微生物生物量碳含量与香樟纯林无显著性差异,微生物生物量氮含量0~10 cm、20~40 cm土层显著高于香樟纯林,10~20 cm土层则低于香樟纯林,这与黄亮亮等[27]、王娟娟等[28]研究结果不一致,这可能归因于近均质条件下排除了树种之外的众多因素(如林分密度、林龄等)对土壤微生物生物量的影响。
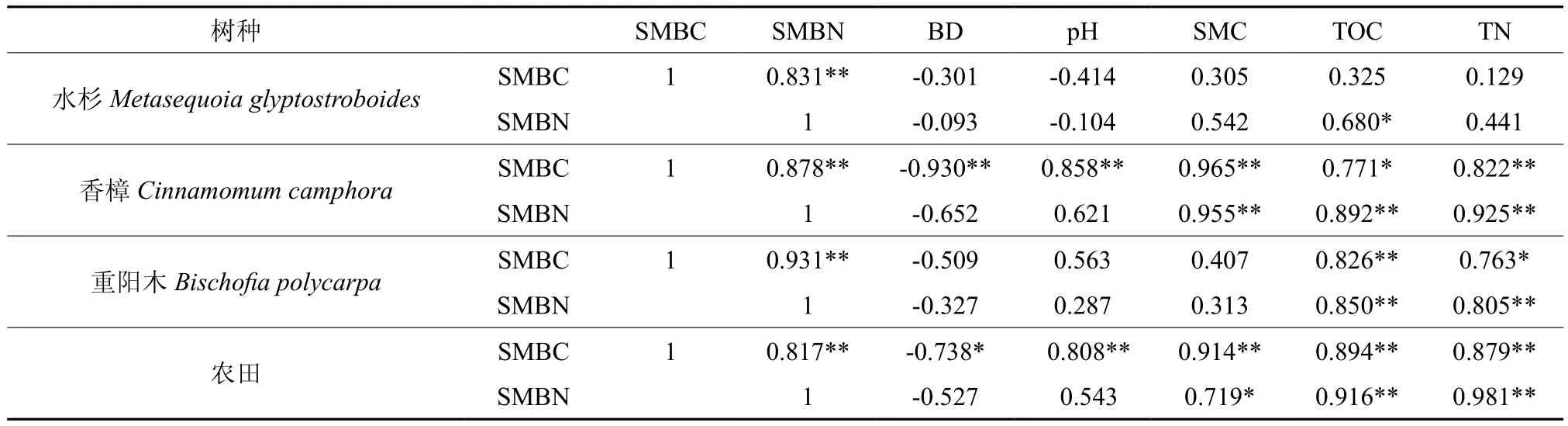
表3 不同树种纯林及土地利用方式土壤微生物生物量碳、氮与土壤理化性质之间的Pearson相关系数†Table 3 Pearson correlation coefficients between soil microbial biomass carbon & nitrogen and soil physicochemical properties in the monocultures of three tree species
有研究表明,不同树种凋落物分解速度影响表层土的微生物的数量,且细菌生物量氮含量高于真菌[29]。水杉等落叶树种的凋落物分解速度快,其土壤细菌数量较多;而香樟等常绿树种凋落物分解率低,土壤细菌数量较少,真菌含量较高[30],所以水杉纯林0~10 cm土层土壤微生物生物量氮含量显著高于香樟。研究发现,土壤微生物生物量峰值比林地凋落物峰值提前一个月,其原因可能是地上部分养分转移导致根系分泌物相对增加,从而促进微生物的生长[31]。水杉纯林凋落物峰值出现在12月[30],并且在叶片凋落之前养分已充分转移至地下根系部分,弱化植物根系与土壤微生物之间对氮素竞争关系,且根系分泌物也相对增加,进而促进20~40 cm土层的微生物生长,而香樟凋落物峰值出现在4、7月[30-32],11月中旬香樟地上部分仍处于非休眠状态,根系与土壤微生物对氮素仍具有一定的竞争关系,可能在一定程度上抑制土壤微生物生长及对氮素的吸收,所以20~40 cm土层土壤微生物生物量氮含量水杉纯林显著高于香樟纯林。各树种纯林土壤微生物生物量均呈现“高-低-高”的垂直分布特征,其中表层土壤(0~10 cm)总体上高于下层土壤(10~20 cm、20~40 cm),这与邵玉琴等[33]、王娟娟[34]等研究相似。其原因可能与土壤受林木干扰程度相关,0~10 cm表层土壤有林地凋落物分解产生的碳氮输入,20~40 cm土层一般是乔木树种根系层,也是根系凋落物、分泌物产生与外生菌根分布的主要区域,故0~10 cm与20~40 cm土层微生物生长直接受林木的影响。
4.2 土地利用方式对土壤微生物生物量碳氮含量的影响
不同土地利用方式下土壤微生物生物量碳氮含量存在一定的差异。本研究中3个树种纯林土壤微生物生物量碳氮含量总体上均与农田差异显著,且随树种的不同,差异性有所不同。重阳木等落叶阔叶树种土壤微生物生物量碳含量与农田土壤差异较小,其余树种则均显著低于农田土壤(图1a),这与黄亮亮等[27]研究结果相似。其原因可能是退耕还林改变了原有的土壤环境(表2),且在造林初期林分凋落物养分归还量较少[35],故而一定程度上抑制了土壤微生物的生长;而在此期间,水稻秸秆、稻茬田间分解以及肥料的使用保证了农田养分的输入,而耕作方式又无较大变化,故而可能存在的“主场优势效应(home- field advantage,HFA)”[36]使得农田土壤微生物变化较小。
4.3 不同树种及土地利用方式下土壤微生物生物量碳氮的影响因子
Pearson相关分析表明,土壤微生物生物量碳氮之间极显著正相关,且主要与有机碳、全氮、土壤水分显著或极显著正相关,这与黄亮亮等[27]、刘爽等[3]研究一致。其原因是土壤微生物生物量碳氮是微生物固有化学属性,土壤有机碳、全氮是微生物碳源与氮源的主要来源,水分则是土壤相关生物化学反应的催化剂。
综上所述,不同树种与土地利用方式下土壤微生物生物量碳氮含量存在一定差异,其中微生物生物量氮含量随树种及土地利用方式的改变差异性显著。不同土地利用方式改变了土壤微生物生物量的垂直分布特征。这些差异与改变可能是因为土壤理化性质、凋落物分解速率、根系分布区间、土壤扰动、植物与微生物之间的养分竞争强度等发生变化,影响了土壤微生物生长与繁殖,以及对土壤养分的吸收与利用。
4.4 不足之处与展望
本研究仅初步探究了树种及土地利用方式对土壤微生物生物量的影响,验证了影响土壤微生物生物量较为常见的相关因子,缺乏清晰地试验地土壤微生物生物量差异形成的机理性原因,而且所选树种亲缘关系相差较大,对于相同科、属的树种是否符合类似情况仍旧存疑,因此,未来有必要针对上述疑问进行进一步研究。
参考文献:
[1]Paul E A, Ladd J N. Soil Biochemistry[M]. New York: Marcel Dekker, 1981: 415-471.
[2]吴金水, 林启美, 黄巧云. 土壤微生物生物量测定方法及其应用[M].北京: 气象出版社,2006.
[3]刘 爽, 王传宽. 五种温带森林土壤微生物生物量碳氮的时空格局[J]. 生态学报, 2010, 30(12):3135-3143.
[4]Sparling G P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter[J]. Soil Research, 1992, 30(2): 195-207.
[5]Joergensen R G, Mueller T. The fumigation-extraction method to estimate soil microbial biomass: calibration of the k EN value.Soil Biology and Biochemistry, 1996, 28(1): 33-37.
[6]张成霞, 南志标. 土壤微生物生物量的研究进展[J]. 草业科学, 2010, 27(6): 50-57.
[7]Somova L A, Pechurkin N S. Functional, regulatory and indicator features of microorganisms in man-made ecosystems[J].Advances in Space Research, 2001, 27(9): 1563-1570.
[8]Wardle D A. A comparative assessment of factors which in fluence microbial biomass carbon and nitrogen levels in soil[J].Biological reviews, 1992, 67(3): 321-358.
[9]王国兵, 阮宏华, 唐燕飞,等. 北亚热带次生栎林与火炬松人工林土壤微生物生物量碳的季节动态[J]. 应用生态学报,2008, 19(1): 37-42.
[10]李胜蓝, 方 晰, 项文化,等. 湘中丘陵区 4 种森林类型土壤微生物生物量碳氮含量[J]. 林业科学, 2014, 50(5): 8-16.
[11]李 斌, 辜 翔, 方 晰,等. 湘中丘陵区土壤微生物指标对土地利用方式的响应[J]. 中南林业科技大学学报, 2015, 35(4):72-77.
[12]王晓龙, 徐立刚, 姚 鑫,等. 鄱阳湖典型湿地植物群落土壤微生物量特征[J]. 生态学报, 2010, 30(18): 5033-5042.
[13]杨 凯, 朱教君, 张金鑫,等. 不同林龄落叶松人工林土壤微生物生物量[J]. 生态学报, 2009, 29(10): 5500-5507.
[14]郭春兰, 张 露, 叶素琼,等. 赣西油茶人工林土壤微生物群落的多样性[J]. 经济林研究, 2015, 33(1):25-32.
[15]王春阳, 周建斌, 董燕婕,等. 黄土区六种植物凋落物与不同形态氮素对土壤微生物量碳氮含量的影响[J]. 生态学报,2010, 30(24): 7092-7100.
[16]李 娇, 蒋先敏, 尹华军,等. 不同林龄云杉人工林的根系分泌物与土壤微生物[J]. 应用生态学报, 2014, 25(2): 325-332.
[17]赵 溪, 李君剑, 李洪建. 关帝山不同植被恢复类型对土壤碳、氮含量及微生物数量的影响[J]. 生态学杂志, 2010, 29(11):2102-2110.
[18]马效国, 樊丽琴, 陆 妮,等. 不同土地利用方式对苜蓿茬地土壤微生物生物量碳、氮的影响[J]. 草业科学, 2005, 22(10):13-17.
[19]彭佩钦, 吴金水, 黄道友,等. 洞庭湖区不同利用方式对土壤微生物生物量碳氮磷的影响[J]. 生态学报, 2006, 26(7): 2261-2267.
[20]秦韵婷, 李建贵, 郭艺鹏,等. PGPR对灰枣土壤养分及微生物数量影响的主成分分析[J]. 经济林研究, 2015, 33(3):39-43.
[21]李新爱, 肖和艾, 吴金水,等. 喀斯特地区不同土地利用方式对土壤有机碳、全氮以及微生物生物量碳和氮的影响[J]. 应用生态学报, 2006, 17(10): 1827-1831.
[22]Schinner F, Öhlinger R, Kandeler D E,et al. Methods in Soil Biology[M]. Berlin:Springer-vetlag, 1995,93-110.
[23]Jenkinson D S. Determination of microbial biomass carbon and nitrogen in soil[J]. Advances in nitrogen cycling in agricultural ecosystems, edited by J. Wilson, 1988, pp.368-386. Wallingford:CAB International.
[24]王巧环, 任玉芬, 孟 龄,等. 元素分析仪同时测定土壤中全氮和有机碳[J]. 分析试验室, 2013, 32(10):41-45.
[25]Wang X, Wang J, Zhang J. Comparisons of three methods for organic and inorganic carbon in calcareous soils of northwestern China[J]. PlOS one, 2012, 7(8): e44334.
[26]许晓静. 不同环境下几种园林树木落叶分解及N、P动态的研究[D]. 合肥: 安徽农业大学, 2007.
[27]黄亮亮, 阮宏华, 王 莹,等. 围湖造田不同利用类型下土壤微生物量碳的特征[J]. 林业科技开发, 2010, 24(4):64-67.
[28]王娟娟, 朱 莎, 靳士科,等. 上海市3种森林类型土壤微生物生物量碳和氮的时空格局[J]. 生态与农村环境学报, 2016,32(4):615-621.
[29]曾全超, 李 鑫, 董扬红,等. 黄土高原不同乔木林土壤微生物量碳氮和溶解性碳氮的特征[J]. 生态学报, 2015,35(11):3598-3605.
[30]高志红. 上海几种主要绿化树种枯枝落叶的特征及生态功能研究[D]. 上海: 华东师范大学, 2005.
[31]Ruan H H, Zou X M, Scatena F N,et al.Asynchronous fluctuation of soil microbial biomass and plant litterfall in a tropical wet forest[J]. Plant and Soil, 2004, 260(1-2): 147-154.
[32]车文玉, 商侃侃, 王妍婷,等. 上海不同发育阶段香樟人工林凋落物及其养分动态[J]. 西北林学院学报, 2016,31(1):42-47.
[33]邵玉琴, 赵 吉, 岳 冰. 皇甫川流域人工油松林地土壤微生物的垂直分布[J]. 内蒙古大学学报(自然科学版), 2002,33(5): 541-545.
[34]王娟娟. 上海市五种绿化群落土壤微生物生物量的特征研究[D]. 上海: 华东师范大学, 2016.
[35]王春梅, 刘艳红, 邵 彬,等. 量化退耕还林后土壤碳变化[J].北京林业大学学报, 2007, 29(3):112-119.
[36]Prescott C E, Grayston S J. Tree species in fluence on microbial communities in litter and soil: current knowledge and research needs[J]. Forest Ecology and Management, 2013(309): 19-27.
猜你喜欢
现代装饰(2022年3期)2022-07-05
意林·少年版(2021年13期)2021-08-30
西安交通大学学报(社会科学版)(2021年2期)2021-04-06
参花·青春文学(2021年3期)2021-03-04
北方音乐(2019年6期)2019-07-16
作文成功之路·作文交响乐(2019年10期)2019-01-10
共产党员·下(2018年6期)2018-12-05
方圆(2018年23期)2018-01-07
小小说月刊·下半月(2015年12期)2015-05-14
发明与创新·中学生(2014年11期)2014-11-24
