
钟萼木当年生生长性状与不同生长类型适应性探讨
2018-03-30 02:33:00郭祥泉钱国钦涂年旺郑经池郭祥堆黄建兰施向东
安徽大学学报(自然科学版) 2018年2期
郭祥泉,钱国钦,涂年旺,郑经池,郭祥堆,黄建兰,施向东
(1.福建省三明市林业局,福建 三明365000;2.福建省世界银行贷款造林项目办公室,福建 福州 350001;3. 福建省永安市林业局,福建 永安 366000;4. 福建省永安市第六中学,福建 永安 366000;5.福建省闽江源国家级自然保护区管理局,福建 建宁 354500;6.福建省大田县赤头坂国有林场,福建 大田366101)
前人研究结果表明,林木的高与地径生长性状由多基因控制,杂交子代的高与地径生长性状表现为多样性生长类型,在早期生长已表现多样性分化,是物种为适应不同生态环境条件进化形成的[1-5].如林木天然更新过程,因一些环境因子(如病原菌侵染、水分胁迫、动物危害等因子)对种子与幼苗生长所产生的影响,通过自然选择把适宜生长的基因类型保存下来.不同环境条件保留下不同的生长类型,经长期自然选择形成多样性生长类型[6].由基因重组理论可知,生长类型遗传多样性,是隐性基因与显性基因在基因重组后形成,多基因控制的生长性状在子代表现多样性生长类型.自然采种与人工筛选优良个体采种基本都是属于天然授粉杂交,在子代中表现多种基因组合的生长类型.钟萼木(BretschneiderasinensisHems1.)为异花授粉濒危物种,天然分布的物种所保存的个体属于能适宜环境生长的个体,自然授粉的杂交子代高与地径生长性状遵循基因重组规律[1,6-8].根据目前自然生长效果与前人的记载,人们对该树种不同个体生长差异与潜在的优势了解很少,作者对该树种苗木与多年生长特性等研究,认为该树种个体生长表现早期遗传分化,优劣势个体年生长量相差较大,为了更好地了解该树种生长特性与潜在的生长[9-13].笔者试图通过群体生长性状分化所表现的不同个体生长类型,探讨它的生长性状多样性遗传和不同生长类型生长规律,以期揭示该树种生长特性和生长性状遗传潜能,为更好利用该物种提供参考.
1 试验材料
1.1 试验地气候条件
试验圃地位于大田县赤头坂国有林场,地处戴云山西侧的闽中低山带,为中亚热带季风气候,年平均气温15.3~19.6℃,无霜期255~275d,年降水量1491.2~1809.6mm,海拔约450m,坡向为南坡,坡度20~32°.土壤以红壤为主.区域内冬季时间较短、夏季较长,气候温和,雨量充沛,温暖湿润,自然条件优越(气象资料来源于大田县气象局).
1.2 试验概况
钟萼木苗木种子来源于福建省永安天宝岩自然保护区天然分布的采种母树,试验圃地海拔450m,圃地为农田,沙壤土;于2015年2月底整地,100kg/亩过磷酸钙作为基肥,畦面宽90cm、沟深20cm,播种前用600倍多菌灵消毒.经休眠催芽后的种子,于2015年3月下旬播种,采用点播形式,沟深10cm、间隔20cm,点播间隔20cm,播种后用细黄心土盖面,即时喷水达畦面潮湿为宜.该树种属于耐荫树种,高温与强光照都对嫩芽与叶产生伤害,所以出芽后搭盖遮阳网(透光度在50%),直到当年的9月上旬左右才能揭去遮阳网[13].该树种个体生长在2.0m以下不分生侧枝,苗木冠幅大小由它的羽状复叶决定,在苗木期,个体叶片数平均达9片,由早期的3出复叶到后期的9~11出复叶间呈轮生,7月份后叶片大小变化不大,通常冠幅的水平半径在15~25cm.整体冠幅较稀疏,个体叶片表现相互交叉,且因成苗率影响和该树种较强的耐荫特性,个体竞争不大,密度效应弱于遗传对生长的影响.
2 调查处理与研究方法
2.1 遗传多样性调查
遗传多样性根据正态分布标准差分类法原理[5],在当年苗木生长结束后(苗木落叶在当年11月底),选择苗木高生长优势木作为调查群体中心,设置调查地块,每个调查地块所调查数量不少于500株,有多株优势木处于同一调查地块的,则设置同一个调查地块处理,所有不在同一调查地块的优势木都另设置调查地块,分别调查苗木地径与苗高生长量,于2015年12月8日调查,共调查6个群体,以高生长优势木作为调查群体中心可以避免好的生长类型不被遗漏.所调查群体处于同一圃地,生长条件基本一致.
调查工具:地径测量采用游标卡尺,苗高采用钢卷尺,分别测量地径与苗高值.
2.2 研究方法
2.2.1遗传多样性

2.2.2不同遗传品质个体生长适应性
苗木的高与地径年节律生长,是从苗木发芽整齐后,在同一苗圃地选择100株定期记录个体生长,生长调查记录从子叶完全展开后开始,此后按每月的1、16日定株定期调查个体高与地径生长,并记录各生长期管理措施.根据上述分类方法划分的不同生长类型,确定各生长类型的平均值,依据各生长类型平均值与定株记录个体生长值比较,从100株定株生长记录中选择相应的生长类型,各取5株观察记录平均值(有的区间不足5株的,以现有株数计算平均数),利用5株记录的生长值平均数作为不同生长类型生长表现.
3 结果与分析
3.1 苗木高生长性状遗传品质多样性
试验苗木为自然授粉的杂交种子代,根据前人研究结果,高生长类型遵循多基因重组规律.通过大样本抽样调查统计分类,群体高生长分布于[13,92] cm 区间,群体的高生长平均值、方差、标准差和波动系数分别是38cm、100、10、0.263,依据上述划分方法,钟萼木当年生苗木高生长划分为如图1所示的8个不同生长类型.
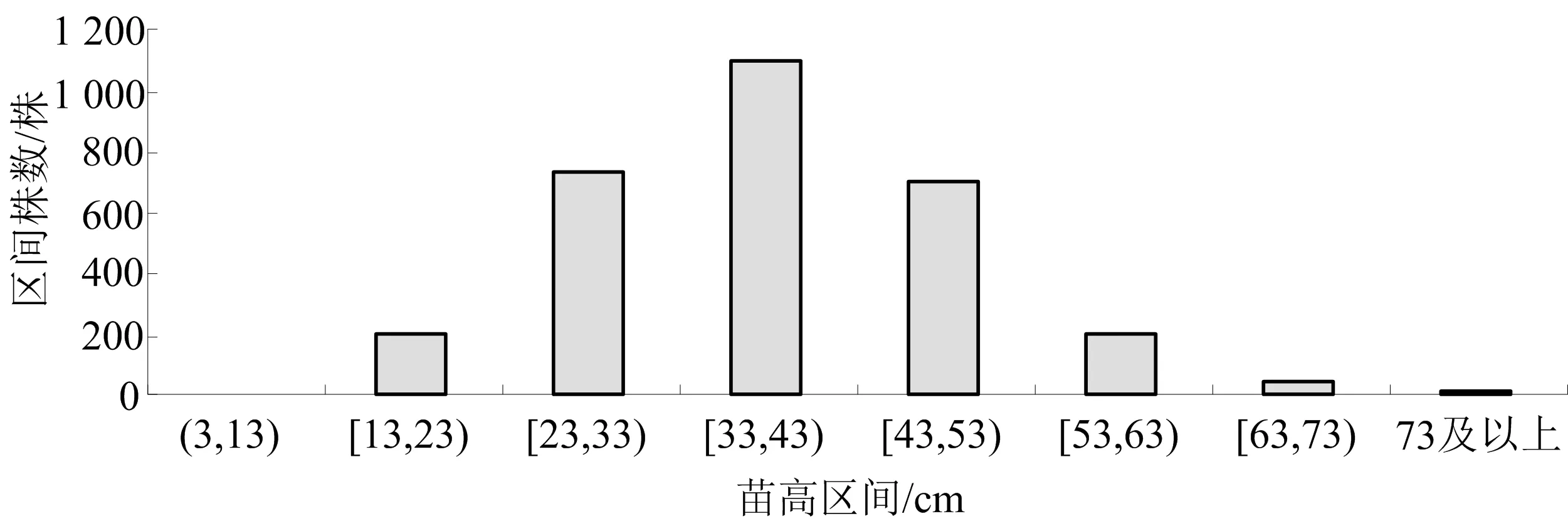
图1 苗高生长不同遗传品质分布

3.2 苗木地径生长性状遗传多样性
良种壮苗的评价,地径生长量是苗木的重要生长指标,直接影响造林后个体生长量.前人研究表明,地径与苗木高生长存在相关性,在生长分化上遵循基因重组规律[1,5].经对钟萼木苗木地径生长调查数据统计,苗木地径生长分布于(0.28,2.02) cm区间,群体的地径生长量平均值、方差、标准差和波动系数分别是1.12cm、0.06、0.24、0.214,群体生长性状分化明显,根据正态分布标准差分类法,以钟萼木当年生苗木地径生长分布将其划分为8个不同生长类型,如图2所示.
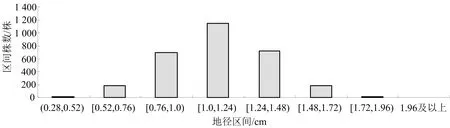
图2 地径生长不同遗传品质分布

3.3 不同生长类型苗木高年生长节律
物种多样性遗传品质在逆境时表现不同抗逆性生长.苗木高生长是林木适应性生长的重要因子,生长性状在多基因控制下表现为多样性生长[1,5,15-16],该调查群体测量结果如图3所示.
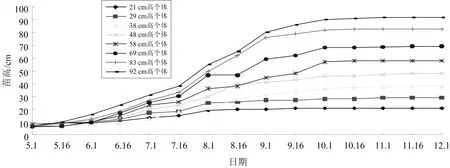
图3 不同遗传品质苗木高年生长适应性
由图3可知,高年生长量低的,生长期很短,个体年苗高21cm,8月份以后就没有高生长了,个体苗高为29、38、48cm的生长类型,在6—8月份是主要生长期,8月16日以后,基本没有或很小生长量,而个体苗高58、69、83、92cm的生长类型,生长期一直持续到10月份,且一直有良好的生长量,特别是在8—9月份还有一较高的生长量,生长期的长短是不同生长类型对环境条件的适应性,适应性强的个体有更长的生长期,有更好的年生长量,适应性差的个体年生长期相对短,也表现较小的年生长量,由不同生长类型的适应性可以较好了解个体生长特性,为该树种去濒危与遗传品质改良提供相关参考.
3.4 不同生长类型苗木地径年生长节律
地径与苗高生长存在相关,苗木地径与苗高生长都是受遗传控制的,通过光合作用与净光合作用积累表现地径生长量,光合作用受冠幅生长和光合作用效率影响,冠幅生长与苗木高生长相关,高生长好的个体,个体冠幅相对大,光合作用面积也较大,直接影响地径生长.高生长表现的个体在空间竞争不是很激烈的条件下,个体的冠幅相对大,净光合作用积累量大,直接表现为木质部的生长.不同个体光合作用率受遗传影响,光利用率高则净光合作用积累量大,表现为良好的地径生长量;光利用率相对低则光合作用净积累量相对小,表现为地径生长量小[18].该调查群体地径生长适应状况如图4所示(图例中括号的数字代表地径所对应的苗高生长量).

图4 地径不同遗传品质生长适应性
由图4可知,地径生长期基本一致,地径生长差异主要受冠幅与光合作用的影响,由遗传因子决定.当然,环境条件中的密度对苗木的地径生长也会产生影响,当苗木培育的密度较大时,因冠幅生长空间竞争而得不到充分生长,地径生长量就偏小,高茎比相对大,显得瘦弱,但这可以通过人为措施进行调控,环境条件产生的可人为干扰,而遗传引起的人为干扰则没有效果[1,5].该树种在苗木期生长没有分生侧枝,株间距相对合理,苗木在当年生长过程空间竞争小,密度对地径生长的影响不大,地径生长的差异主要来源于遗传.由图4可知,苗木高分别为83、92cm的个体处于群体的上层,有良好的生长空间,地径生长量较大,从7—11月份一直保持良好的地径生长,11月以后基本停止生长.8月份以后在个体高生长分化加剧后,生长差的个体地径生长增加量也小,这表明遗传因子对个体生长的影响.
4 结束语
多样性生长类型是物种适应不同生态环境时生存的需要,前人对多样性研究,多数是进行不同物种的生物多样性研究,对于同一物种的多样性生长类型主要是进行基因亲源性研究[1,5,13-14],对于生长性状多样性研究少见相关资料[5].林木的高与地径生长是由多基因控制,遵循基因重组规律并表现不同生长类型[1-2,5],试验划分不同生长类型群体所占比例与基因重组后子代分化比例基本一致.钟萼木是濒危树种,所保留下来的个体能适应所处环境,进行遗传多样性生长适应性研究,能更好地了解该树种遗传特性与不同生长类型的适应性生长规律.
应用正态分布标准差分类法划分不同生长类型,表明好的生长类型具有良好的遗传品质,个体表现良好的年生长量,差的生长类型表现差的遗传品质.试验研究较科学地分析了该物种生长特性与生长潜能,能较好地说明物种生长潜能与对环境的适应能力.前人对该物种评价以群体平均值说明该物种的生长特性的研究,较难清楚了解物种生长潜能与遗传品质.对钟萼木进行不同生长类型适应性研究,可较好地揭示不同生长类型年生长规律与优良遗传个体生长潜能,也让人们知道,虽然该树种处于濒危状态,但现存的天然群体中包含有优良基因资源,可以通过不同生长类型划分,筛选优良个体,利用优良个体遗传品质改良现有资源状况,可为物种保护与利用提供依据[7-11].
研究表明,不同生长类型苗木高与地径生长表现不同的生长适应性,该树种生长类型差的个体高生长期较短,优良生长类型个体有更长的高生长期,但地径生长期基本是一致的,主要受高生长期与光合作用率影响,不同生长类型光合作用效率与净光合作用积累的潜能存在差异[18],进行生长类型分类,可以揭示不同生长类型的遗传差异,了解该物种不同生长类型净光合作用积累潜能,为生理生化与基因研究提供参考.
参考文献:
[1] 郭祥泉.邓恩桉栽培学[M]. 北京: 中国林业出版社,2015:207-247.
[2] 梁一池. 树木育种原理与方法[M]. 厦门: 厦门大学出版社,1998:98-139.
[3] 王明麻. 林木育种学概论[M]. 北京: 中国林业出版社,1989:67-145.
[4] 郭祥泉, 洪伟, 吴承祯, 等. “t”检验法则在闽北抗寒速生邓恩桉优株筛选的应用[J]. 应用与环境生物学报,2009,15(3):385-389.
[5] 郭祥泉. “正态分布标准差分类法”对杉木养分供求响应的研究[J]. 西北林学院学报,2016,31(5):132-137.
[6] 陈远台, 郭祥泉, 郭奥捷. 近自然林业生态丰产机理与生产应用模式探讨[J]. 西南林业大学学报,2013,33(6):7-13.
[7] 宋朝枢. 中国珍稀濒危保护植物[M]. 北京: 中国林业出版牡,1989:204-205.
[8] 张若慧. 浙江珍稀濒危植物[M]. 杭州: 浙江科学技术出版社,1994:188-189.
[9] 郭祥泉, 李延群, 孙晓冬, 等. 应用“t”检验选择法筛选钟萼木幼树优株的探讨[J]. 亚热带植物科学,2011,40(4):38-42.
[10] 李铁华, 周佑勋, 段小平. 钟萼木种子生理休眠特性的初步研究[J]. 中南林学院学报,1997,17(2):41-44.
[11] 郭治友, 龙应霞, 肖国学. 钟萼木的组织培养和快速繁殖[J]. 植物生理学通讯,2007,43(1):127-130.
[12] 郭祥泉, 周立华, 熊自华, 等. 孑遗树种钟萼木多年生幼树生长特性探讨[J]. 亚热带植物科学,2012,41(2):32-36.
[13] 郭祥泉, 钱国钦, 施向东, 等. 濒危树种钟萼木苗木年生长节律与适应性调查[J]. 亚热带植物科学,2016,45(4):351-356.
[14] 洪伟, 吴承祯. 试验设计与分析[M]. 北京: 中国林业出版社,2004:7-90.
[15] 王雷存, 万怡震, 高华, 等. “富士×嘎拉”F1代杂种苗早期选择试验[J]. 西北林学院学报,2010,25(2):80-82.
[16] 李鹏丽, 申凤莲, 毛永民, 等. 果树性状遗传规律研究进展[J]. 河北农业大学学报,2003,26(5):53-57.
[17] 陈存及, 陈伙法. 阔叶树种栽培[M]. 北京: 中国林业出版社,2000:141-329.
[18] 李合生. 现代植物生理学[M]. 北京: 高等教育出版社,2002:178-246.
猜你喜欢
区域治理(2022年40期)2022-11-27 04:01:54
绿色科技(2021年21期)2021-11-26 09:12:06
内蒙古林业调查设计(2021年3期)2021-07-06 14:12:00
动漫界·幼教365(小班)(2019年10期)2019-10-28 02:04:20
动漫界·幼教365(大班)(2019年10期)2019-10-28 01:54:09
动漫界·幼教365(中班)(2019年10期)2019-10-28 01:53:17
现代园艺(2018年2期)2018-03-15 08:00:35
现代园艺(2018年1期)2018-03-15 07:56:13
中国林业产业(2016年5期)2016-04-03 00:33:02
中国林业产业(2016年5期)2016-04-03 00:32:38
