
物体认知
——基于动物自发行为的一种学习记忆评价方法的研究
2018-03-29 06:35吕静薇宋广青董黎明陈善广刘新民
中国比较医学杂志 2018年3期
吕静薇,宋广青,董黎明,姜 宁,陈善广,刘新民*
(1.中国医学科学院/北京协和医学院药用植物研究所,北京 100193;2.中国航天员科研训练中心人因工程重点实验室,北京 100094)
物体认知实验方法是1988年由Ennaceur提出,它是基于啮齿类动物天生具有的新奇偏爱性,用于检测动物的学习记忆能力与传统的检测方法相比,不需给予正向(奖赏)或反向(惩罚)的刺激,也不需冗长的训练,动物分别根据“新旧物体”特征、位置、出现的顺序和所处的背景对“新旧物体”进行辨别,能够检测从简单的物体识别记忆到复杂的空间、时序和情景记忆[1],对动物学习记忆行为实现精细化区分,实验简单易操作。在学习记忆发生发展机制、认知障碍防护药物和保健品研发等方面具有重要应用价值[2-3]。但目前国内尚未见对其四种实验模式及操作影响因素的系统研究。
本文选择两种常用品系小鼠,对新物体识别、物体位置识别、时序记忆和情景记忆四个模式的实验方法操作步骤,以及测试期时间、熟悉期与测试期间隔时间等因素展开研究,为物体认知的应用提供稳定可靠、客观规范的操作方法。
1 材料和方法
1.1 实验动物
SPF级,ICR雄性小鼠108只和KM雄性小鼠48只,24~28 g,购于北京维通利华实验动物技术有限公司[SCXK (京) 2012-0001]。动物自购入后先进行一周的适应性饲养,实验期间给予动物充足的食、水,实验室保持安静,温度(23±2)℃,湿度(50±10)%,12 h照明/12 h黑暗环境(上午8:00开灯)[SYXK (京) 2013-0023]。实验遵守国际实验动物伦理学要求,实验过程中按实验动物使用的3R原则给予人道主义关怀。每天进行实验的时间固定,实验开始前将动物由动物房移入实验室中适应约20 min,实验装置调节灯光强度在60 lx左右。
1.2 主要仪器
物体认知实验设备:由中国医学科学院药用植物研究所、中国航天员科研训练中心、北京康森益友有限公司联合研制。设备包括四个测试箱和不同的物体,并于箱体顶部装置摄像头进行实时监测。
1.3 实验方法
1.3.1 新物体识别实验
适应期实验中将动物放入没有任何物体的实验箱中,每只每天10 min,每天1次,连续3 d。第4天进行熟悉期实验,开始时箱中不放入任何物体,将动物放入再次适应2 min,动物取出后,放入两个完全相同的物体,分别记录每只动物5 min内对左侧物体的探索时间(exploration time for left object,Tl)和对右侧物体的探索时间(exploration time for right object,Tr)。
熟悉期后将动物放回饲养笼中,10 min后开始测试期实验。测试期时放入两个不同的物体,其中一个物体与熟悉期的物体完全相同,另外一个是新奇物体(与熟悉期物体匹配),动物放入后计时5 min,分别记录动物在不同时间段对熟悉物体的探索时间(exploration time for familiar object,Tf)和对新奇物体的探索时间(exploration time for novel object,Tn)。
探索活动定义为:小鼠的鼻子近距离指向物体或者直接嗅、舔物体。当其他部位接触物体而鼻子没有指向物体或站在物体上时非对物体的探索活动[4]。
1.3.2 物体位置识别实验
适应期与熟悉期实验过程同新物体识别实验,而测试期与新物体识别实验不同,只是将熟悉期中两个相同物体中的一个移到新的位置,动物放入后计时5 min,分别记录动物在不同时间段对两不同物体的探索时间。
1.3.3 时序记忆实验
时序记忆实验程序和上述两种方法有所不同,实验流程见图1,包括2个熟悉期,即熟悉期1和熟悉期2,其他过程基本相同。适应期同新物体识别。
第4天进行熟悉期实验,开始时箱中无任何物体,将动物放入再次适应2 min,动物取出后放入两相同的物体,进行熟悉期1实验,分别记录5 min内对两个完全相同物体的探索时间;将动物取出后放入饲养笼中,10 min后进行熟悉期2实验,在同一个实验箱中放入另外两个完全相同的物体(与熟悉期1中物体匹配),分别记录5 min内对两个完全相同物体的探索时间。熟悉期2后将动物放回饲养笼中,10 min后开始测试期实验。测试期时放入两个不同的物体,分别是熟悉期1和熟悉期2中各一个物体,动物放入后计时5 min,分别记录在不同时间段对两不同物体的探索时间。
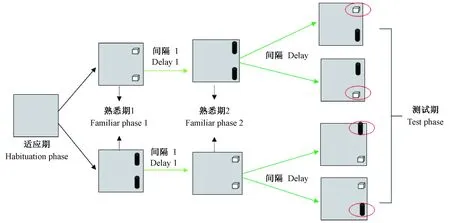
注:以红色圆圈标记物体为新奇物体。图1 时序记忆实验操作流程图Note. Red circles indicate novel objects.Fig.1 Schematic diagram of the experimental process of temporal order memory task

注:以红色圆圈标记物体为新奇物体。为保持配对平衡,A、B两种放置方式均需考虑。图2 情景记忆实验操作流程图Note. Red circles indicate novel objects. Objects should be placed as A and B for counterbalance.Fig.2 Schematic diagram of the experimental process of object context recognition task
1.3.4 情景记忆实验
情景记忆实验程序与时序记忆实验类似,也包括2个熟悉期,但在不同背景的实验箱中进行,实验流程见图2。
适应期实验将动物放入具有第一种背景的实验箱中,适应完成10 min后,再放入第二种背景的实验箱中,每种实验箱适应过程与新物体识别中适应期实验相同。第4天开始熟悉期实验,在具有第一种背景的实验箱中放入两相同的物体,进行熟悉期1实验,分别记录5 min内对两个完全相同物体的探索时间;将动物放回饲养笼中,10 min后进行熟悉期2实验,在具有第二种背景的实验箱中放入另外两个完全相同的物体(与熟悉期1中的物体匹配),分别记录5 min内对两相同物体的探索时间。熟悉期2后将动物取出放入饲养笼中,10 min后进行测试期实验,计时5 min,分别记录在不同时间段对两不同物体的探索时间。
1.3.5 不同时间间隔时的物体辨别记忆能力考察
将动物随机分为5组,熟悉期与测试期间隔分别设置为10 min,30 min,60 min,90 min,120 min。实验步骤与新物体识别实验相同,仅每组动物检测时熟悉期与测试期间隔不同。记录每组动物在5 min测试期中不同时间段对两不同物体的探索时间。
1.4 统计学方法
2 结果
2.1 新物体识别实验结果分析
由图3可知,KM和ICR小鼠熟悉期对两完全相同物体的探索时间差异无显著性,动物对物体放置的位置未表现出偏爱性。由图4A和4B可知,测试期实验时,KM小鼠在1~5 min和ICR小鼠在2~5 min时Tn显著多于Tf,DI与0相比有显著性(P< 0.01),两品系小鼠可识别新旧物体。
2.2 物体位置识别实验结果分析
相较于原位置的物体,动物对移动到新位置的物体更感兴趣。Tn表示测试期移动到新位置物体的探索时间,Tf表示测试期对原位置物体的探索时间。由图5A可知,KM和ICR两种小鼠在物体位置识别实验中,测试期实验时间为3~5 min时,Tn都显著多于Tf(P< 0.05,P< 0.01);图5B表示在测试期实验时间为2~5 min时,KM和ICR两种小鼠DI都显著大于0(P< 0.05,P< 0.01),两品系小鼠可辨别出物体位置的变化。
2.3 时序记忆实验结果分析
由于出现的时序不同,相对于熟悉期2,动物对先出现的熟悉期1物体更感兴趣。熟悉期1中的物体作为新物体,其探索时间以Tn表示,熟悉期2中的物体探索时间以Tf表示。由图6A可知,测试期实验KM小鼠3 min时和ICR小鼠1 min、4 min或5 min时Tn显著多于Tf(P< 0.05),由图6B可知,KM小鼠3 min或5 min时DI显著大于0(P< 0.05),ICR小鼠3~5 min时DI都大于0(P< 0.05,P< 0.01)。两种品系的小鼠对先后遇到的物体是有记忆的,都能够根据出现的前后对其进行辨别。

图3 熟悉期KM和ICR小鼠对两物体放置位置偏爱性的比较Fig.3 Comparison of time consumption of the KM and ICR mice for exploring both familiar objects during familiar phase in the novel object recognition task

注:A:测试期探索时间;B:相对辨别指数。与Tf比较,**P< 0.01;与0比较,## P< 0.01。图4 测试期KM和ICR小鼠的物体辨别记忆能力比较Note. A: Exploring time during test phase. B: Relative discrimination index. Compared with Tf,**P< 0.01. Compared with 0,##P< 0.01.Fig.4 Comparison of the object recognition ability between the KM and ICR mice during test phase
2.4 情景记忆实验结果分析
测试背景与物体之前所处实验背景不匹配或不一致的物体更能引起啮齿类动物的兴趣,与测试期实验箱背景不匹配物体可看作新物体,测试期探索时间即Tn,而与测试期实验箱背景匹配物体探索时间即Tf。由图7A可知,测试期KM小鼠3~5 min时和ICR小鼠4 min及5 min时Tn均显著多于Tf(P< 0.05,P< 0.01);由图7B可知,KM小鼠在2~5 min时DI显著大于0(P< 0.05,P< 0.01),ICR小鼠在4 min或5 min时DI都大于0(P< 0.01)。两种品系的小鼠在该种实验条件下,对物体熟悉期所处的实验箱背景是有记忆的,都能够根据物体与实验箱背景是否匹配对其进行辨别。

注:A:测试期探索时间;B:相对辨别指数。与Tf比较,* P< 0.05,**P< 0.01;与0比较,# P< 0.05,## P< 0.01。图5 测试期KM和ICR小鼠的物体位置辨别记忆能力比较Note. A: Exploring time during test phase. B: Relative discrimination index. Compared with Tf,*P< 0.05,**P< 0.01. Compared with 0,#P< 0.05,##P< 0.01.Fig.5 Comparison of the object location recognition ability between the KM and ICR mice during test phase
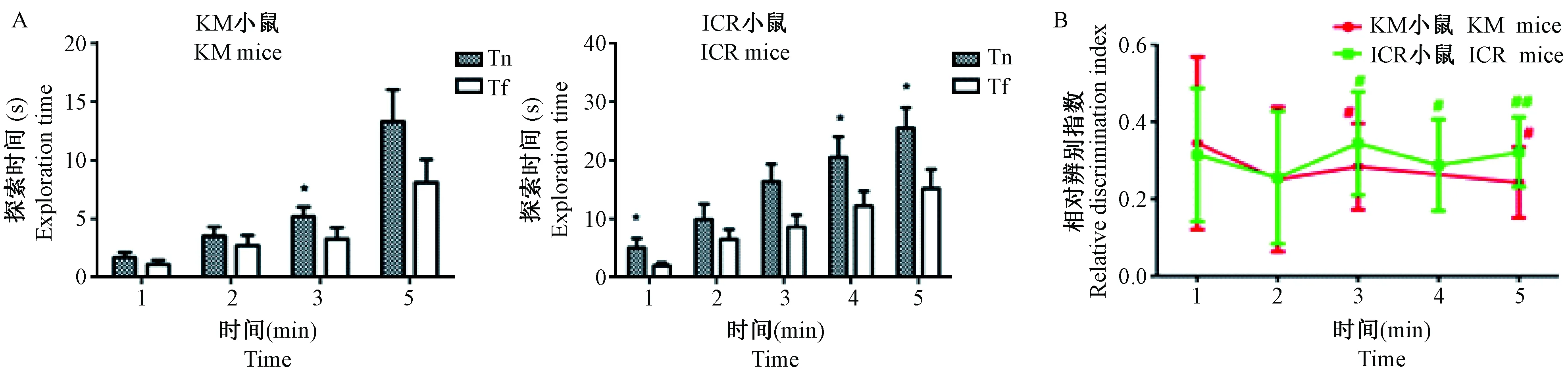
注:A:测试期探索时间;B:相对辨别指数。与Tf比较,* P< 0.05,**P< 0.01;与0比较,# P< 0.05,## P< 0.01。图6 测试期KM和ICR小鼠的时序记忆能力比较Note. A: Exploring time during test phase. B: Relative discrimination index. Compared with Tf,*P< 0.05,**P< 0.01. Compared with 0,#P< 0.05,##P< 0.01.Fig.6 Comparison of the object temporal order memory ability between the KM and ICR mice during test phase
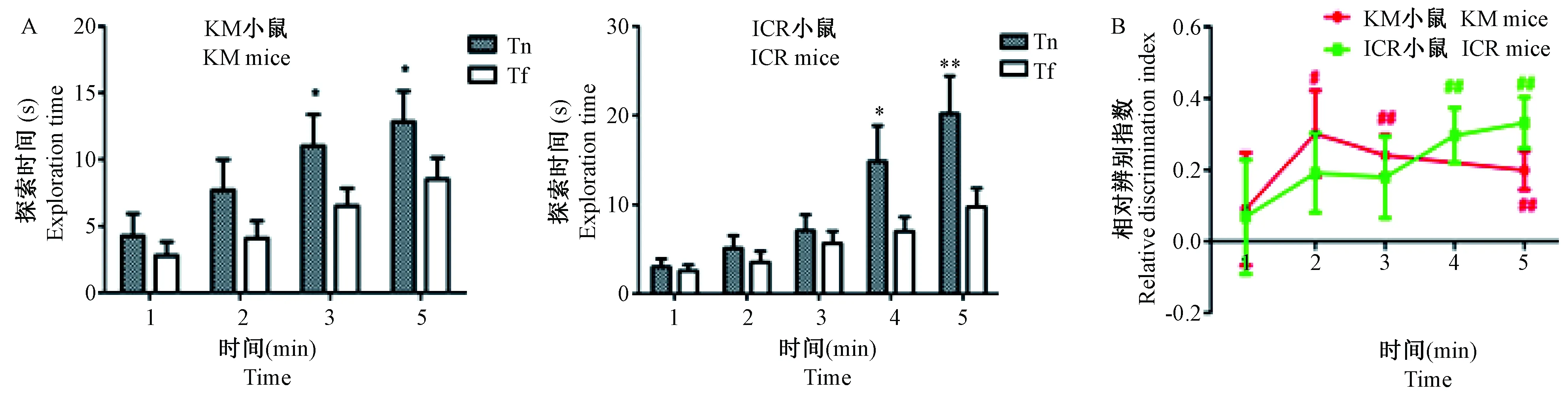
注:A:测试期探索时间;B:相对辨别指数。与Tf比较,* P< 0.05,** P< 0.01;与0比较,# P< 0.05,## P< 0.01。图7 测试期KM和ICR小鼠的情景记忆能力比较Note. A: Exploring time during test phase. B: Relative discrimination index. Compared with Tf,*P< 0.05,**P< 0.01. Compared with 0,#P< 0.05,##P< 0.01.Fig.7 Comparison of the object context recognition ability between the KM and ICR mice during test phase
2.5 不同时间间隔时的物体辨别记忆能力
由图8可知,新物体识别实验熟悉期与测试期的间隔(delay)10 min,ICR小鼠在测试期2~5 min时,DI显著大于0(P< 0.01);当delay=30 min,ICR小鼠在测试期3~5 min时,DI显著大于0(P< 0.05,P< 0.01);当delay=1 h,ICR小鼠在测试期4 min或5 min时,DI显著大于0(P< 0.05)。以上结果表明,ICR小鼠时间间隔≤ 1 h时,能够辨别出新奇物体和熟悉物体,当时间间隔延长到90 min或2 h时则难以辨别。

注:与0比较,# P< 0.05,## P< 0.01。图8 ICR小鼠不同时间间隔下的物体辨别记忆能力Note. Compared with 0,#P< 0.05,##P< 0.01.Fig.8 Object recognition and memory ability of the ICR mice at different time intervals
3 讨论
在物体认知实验中,需充分考虑动物对物体的识别能力和物体配对的平衡。物体材料要相同,同时避免气味触觉影响[5],另考虑对物体尺寸与动物的体长的关系[6-8]。除对物体本身的要求外,另需注意实验过程中物体的取拿及擦洗,如保持同种材料手套的佩戴,每只动物实验完成后,采用10%的酒精进行擦拭,每阶段实验完成后用75%酒精擦洗,后用清水冲洗干净,晾干备用等[9]。
研究表明“新奇偏爱性”存在时间有一定限度[10],动物对物体的探索主要集中在实验开始的几分钟内[11-12]。本实验中两种小鼠在测试期总时间为5 min,各时间段内的DI有所不同,但在3~5 min时间段时最为敏感,能够反应出动物的识别记忆能力。
由于存在自然遗忘现象,随着熟悉期和测试期时间间隔延长,动物对不同物体的识别难度增加。ICR小鼠在新物体识别实验中,自然遗忘开始在1 h~90 min之间。Bertaina-Anglade等发现大鼠在10 min~3 h内,小鼠在1 h时表现正常辨别记忆能力[13]。确定自然遗忘开始时间是进行物体认知实验的前提,一般选较长间隔用于筛选改善作用潜在药物,而选较短间隔检测记忆损伤作用[14]。
物体认知实验方法经调整后,可用于不同程度以及不同类型的学习记忆评价[15-16],并具有简单易操作、周期较短[17-18]、不需要额外的动机(奖励或惩罚)等优点。其中,物体位置识别实验考察动物的空间记忆能力,另三种为非空间记忆能力检测方法。除此之外,时序及情景记忆能力能够充分模拟人类日常生活的识别记忆。目前,物体认知实验中探索时间多采用人工计时,主观性和判断标准难统一。Matsumoto等采用3D跟踪系统进行分析[19],但其准确性有待考证。总之,利用计算机图像技术实现探索时间自动化分析,优化探索标准和评价指标,是物体认知实验未来发展方向,此种精细敏感的行为学实验方法也将获得更广泛的应用。
[1] Dere E, Huston JP, De Souza Silva MA. The pharmacology, neuroanatomy and neurogenetics of one-trial object recognition in rodents [J]. Neurosci Biobehav Rev, 2007, 31(5): 673-704.
[2] Mathiasen JR, DiCamillo A. Novel object recognition in the rat: a facile assay for cognitive function [J]. Curr Protoc Pharmacol, 2010, Chapter 5: Unit 5.59.
[3] Brancato A, Lavanco G, Cavallaro A, et al. The use of the Emotional-Object Recognition as an assay to assess learning and memory associated to an aversive stimulus in rodents [J]. J Neurosci Methods, 2016, 274: 106-115.
[4] Lueptow LM. Novel object recognition test for the investigation of learning and memory in mice [J]. J Vis Exp, 2017(126).
[5] Braida D, Donzelli A, Martucci R, et al. Mice discriminate between stationary and moving 2D shapes: application to the object recognition task to increase attention [J]. Behav Brain Res, 2013, 242: 95-101.
[6] Heyser CJ, Chemero A. Novel object exploration in mice: Not all objects are created equal [J]. Behav Processes, 2012, 89(3): 232-238.
[7] Jurado-Berbel P, Costa-Miserachs D, Torras-Garcia M, et al. Standard object recognition memory and “what” and “where” components: improvement by post-training epinephrine in highly habituated rats [J]. Behav Brain Res, 2010, 207(1): 44-50.
[8] 宋广青, 孙秀萍, 刘新民. 大鼠物体识别实验方法综述 [J]. 中国比较医学杂志, 2013, 23(7): 55-60.
[9] Vogel-Ciernia A, Wood MA. Examining object location and object recognition memory in mice [J]. Curr Protoc Neurosci, 2014, 69: 8.31.1-8.31.17.
[10] Ennaceur A. One-trial object recognition in rats and mice: methodological and theoretical issues [J]. Behav Brain Res, 2010, 215(2): 244-254.
[11] Akkerman S, Blokland A, Reneerkens O, et al. Object recognition testing: methodological considerations on exploration and discrimination measures [J]. Behav Brain Res, 2012, 232(2): 335-347.
[12] Dix SL, Aggleton JP. Extending the spontaneous preference test of recognition: evidence of object-location and object-context recognition [J]. Behav Brain Res, 1999, 99(2): 191-200.
[13] Bertaina-Anglade V, Enjuanes E, Morillon D, et al. The object recognition task in rats and mice: a simple and rapid model in safety pharmacology to detect amnesic properties of a new chemical entity [J]. J Pharmacol Toxicol Methods, 2006, 54(2): 99-105.
[14] Lyon L, Saksida LM, Bussey TJ. Spontaneous object recognition and its relevance to schizophrenia: a review of findings from pharmacological, genetic, lesion and developmental rodent models [J]. Psychopharmacology (Berl), 2012, 220(4): 647-672.
[15] Ishikawa H, Yamada K, Pavlides C, et al. Sleep deprivation impairs spontaneous object-place but not novel-object recognition in rats [J]. Neurosci Lett, 2014, 580: 114-118.
[16] Bello-Medina PC, Sánchez-Carrasco L, González-Ornelas NR, et al. Differential effects of spaced vs. massed training in long-term object-identity and object-location recognition memory [J]. Behav Brain Res, 2013, 250: 102-113.
[17] Lu C, Shi Z, Dong L, et al. Exploring the effect of ginsenoside Rh1 in a sleep deprivation-induced mouse memory impairment model [J]. Phytother Res, 2017, 31(5): 763-770.
[18] Wang K, Lu JM, Xing ZH, et al. Effect of 1.8 GHz radiofrequency electromagnetic radiation on novel object associative recognition memory in mice [J]. Sci Rep, 2017, 7: 44521.
[19] Matsumoto J, Uehara T, Urakawa S, et al. 3D video analysis of the novel object recognition test in rats [J]. Behav Brain Res, 2014, 272: 16-24.
猜你喜欢
导航定位学报(2022年5期)2022-10-13
小猕猴智力画刊(2022年3期)2022-03-28
昆明医科大学学报(2021年10期)2021-12-02
科学大众(2021年6期)2021-07-20
学苑创造·A版(2020年9期)2020-10-13
铁道建筑技术(2020年11期)2020-05-22
中学生数理化·高一版(2020年1期)2020-02-20
娃娃乐园·综合智能(2019年3期)2019-04-03
电子制作(2017年13期)2017-12-15
少儿科学周刊·儿童版(2015年2期)2015-07-07
