
Growth and nutrient dynamics of Betula alnoides seedlings under exponential fertilization
2018-03-27 12:10LinChenChunshengWangBernardDellZhigangZhaoJunjieGuoDapingXuJieZeng
Journal of Forestry Research 2018年1期
Lin Chen·Chunsheng Wang·Bernard Dell·Zhigang Zhao·Junjie Guo·Daping Xu·Jie Zeng
Introduction
Betula alnoides is a fast-growing valuable hardwood species indigenous to Southeast Asia and South China.Its wood is commonly used for making high quality furniture,floor boards,decorative veneer and materials for room decoration and its wood products are popular due to their affordable price.It is an environment-friendly tree species because abundant vegetation can grow beneath its canopy and a number of tree species can grow well with it(Zeng et al.2006).It is normally a dominant species in natural water conservation forests near villages in rural areas.Therefore,B.alnoides plays important roles not only in valuable timber production,but also in the establishment of ecologically oriented plantations.It has been used in largescale plantation development in tropical and warm subtropical areas in South China and Southeast Asia.However,seedling quality remains an important issue(Jiang et al.2003),and production of robust nursery stock is a key driver of its survival and early-growth performance after outplanting.
Reasonable balanced fertilization can improve seedling quality and is particularly essential to grow robust seedlings by promoting survival,early growth of plantations and resistance to diverse stresses such as water de ficit,low temperate and disease(Van den Driessche 1991;Salifu et al.2008;Heiskanen et al.2009;Duan et al.2013;Wang et al.2015).Nutrient loading can also improve growth performance of seedlings when out-planted on weed-prone and nutrient-stressed sites(Salifu and Timmer 2003).Thus,a conceptual model of nutrient loading proposed by Timmer(1996)has been successfully applied in many crops and tree species to distinguish between nutrient de ficiency,suf ficiency,luxury consumption(nutrient loading),as well as optimum and toxicity levels.Although B.alnoides has become a major timber species with large scale plantations in South China and Southeast Asia,only a few studies have been conducted on fertilization even for seedling production.Jiang et al.(2003)demonstrated that application of compound fertilizer(N,P and K)or urea(N)could promote seedling growth of B.alnoides,and our study on its seedling responses to macronutrient de ficiencies showed that severe nitrogen and potassium de ficiencies could inhibit growth of B.alnoides seedlings(Chen et al.2010a).Nonetheless,its nutrient requirements and fertilization have not yet been fully understood.
To determine the nitrogen requirement necessary for robust seedling production,we have carried out an exponential fertilization trial for B.alnoides seedlings.We previously assessed growth performance and leaf nutrient status of seedlings under exponential fertilization and used the critical level approach to analyze the leaf nutrient status of seedlings(Chen et al.2010b).However,further understanding of interactions among nitrogen,phosphorus and potassium,as well as the relationship between growth and nutrient supply,are needed to improve the fertilization regime for robust seedling production of this species.Vector analysis,based on seedling biomass and wholeplant nutrition,is an ef ficient method for this purpose(Haase and Rose 1995;Imo and Timmer 1997;Quoreshi and Timmer 2000).
Therefore,this study aims to determine the responses of B.alnoides seedlings to nutrient supply in terms of growth and nutrient dynamics,to identify the interactions among nitrogen,phosphorus and potassium during the whole experimental period through vector analysis,and to reveal the feasibility of nutrient loading for this species by a conceptual model in relation to nutrient supply,plant dry mass and nutrient status.The hypothesis tested was that reasonable fertilization could improve the growth and nutrient uptake of B.alnoides seedlings.The findings will improve understanding on nutrient demand of B.alnoides and provide evidence for the future production of technical guidelines for the nursery production of robust stock.
Materials and methods
Plant material and cultural conditions
Betula alnoides seedlings of approximately 4.7 cm height,produced from seeds from Jingxi County,Guangxi,were transplanted to 230 cm3plastic pots arranged in 24 cavity trays in a ventilated greenhouse at the Research Institute of Tropical Forestry,CAF,Guangzhou,China on October 21,2008,as described in our previous article(Chen et al.2010b).To prevent runoff of water or leaching of nutrients,white plastic bags were placed in the pots,which were then uniformly filled with a mixture of autoclaved peat,vermiculite,and perlite mixture(3:2:2 for v:v:v).The mean daytime temperature of the greenhouse ranged from 15 to 30°C,naturallightintensity ranged from 110 to 140 μmol m-2s-1and relative air humidity from 23 to 60%during the experimental period.
Experimental design and seedling management
The experiment was carried out as a randomized complete block design with four blocks under the control and six exponential fertilization treatments.The seven treatments were 0,50,100,200,300,400 and 600 mg N seedling-1,coded as E0,50E,100E,200E,300E,400E and 600E,respectively.This range of concentrations was selected because B.alnoides is a nutrient-loving species(Jiang et al.2003),and optimum amounts of nitrogen might exceed those for other broadleaf species,e.g.,25–100 mg N seedling-1for Quercus rubra(Salifu and Jacobs 2006)and 200 mg N seedling-1for Mytilaria laosensis(Chen et al.2012).Each plot contained 48 seedlings,and a total of 1344 seedlings were used in the experiment.The seedlings were supplied with a commercial water-soluble fertilizer mixture(N–P2O5–K2O:20:20:20,plus micronutrients,Plant Products Co.Ltd.,Brampton,Ontario,Canada).Fertilization treatments began 2 weeks after transplanting and were applied on a weekly basis for 12 weeks.At each time of fertilization,seedlings received the same concentration but different volumes as a soil drench for six exponential fertilization treatments.The concentration of applied fertilizer was 0.3%for the first 5 weeks after fertilization and then increased to 0.5,1,2,5 and 10%at weeks 6,7,8,9 and 10–12 after fertilization,respectively,while the volume varied from 0 to 14.97 mL seedling-1calculated by the concentration and amount of fertilizer(Table 1)at each time.The amount of fertilizer applied ateach time was calculated according to the following exponential function described by Timmer(1996):

Table 1 Weekly N addition schedule of Betula alnoides seedlings under seven fertilization treatments

where,NTis the applied amount of N(ranging from 50 to 600 mg N seedling-1)in the total number of fertilizer application(t=12 weeks),NSis the initial level of N in the whole plant(0.15 mg)determined at the beginning of the fertilization,and r is the relative addition rate needed weekly to increase NSto a final level NT+NS.Once r was determined for the fertilization period,the quantity of N to be added on a speci fic day(Nt)was calculated from the following equation:

where,Nt-1is the cumulative amount of N added up to and including the previous fertilization.The weekly application rates are shown in Table 1.
Before the experiment,we adopted the measure of Timmer and Armstrong(1989)to determine the maximum water-holding capacity of the peat mixture and used 70%of the capacity as a daily irrigation standard.Afterward,the trays were weighed every day and watered to reach the appropriate mass.The positions of trays were moved randomly by hand in each block to remove any edge effects every 2 weeks throughout the experiment.
Harvesting and measurements
Plant growth,nutrient composition(N,P and K)and biomass were assessed at 5,8,11,and 14 weeks after transplanting(or 3,6,9 and 12 weeks after the first nutrient addition).At each survey,20 seedlings per plot were randomly selected to measure height and root collar diameter.Seven random seedlings in each plot were generally sampled each time,except that only three seedlings were sampled in each plot of the 600E treatment at the last harvest because fewer seedlings were left due to nutrient toxicity.Plants were separated from the peat mixture by gently washing with distilled water.The whole plant was separated into shoot and root at the root collar,and ovendried at 65°C for 48 h for dry matter determination.Samples were combined and ground with a hammer mill for subsequent chemical analysis.The plant material was wet-digested in a block digester using a H2SO4–HClO4solution.The digest was analyzed for total N by diffusion distillation,P by the molybdenum blue method and K by atomic absorption spectroscopy(LY/T 1271-1999).
Statistical analyses and data interpretation
One-way ANOVA was conducted to examine the effects of different fertilization treatments on growth performances and nutrient conditions of B.alnoides seedlings using SPSS 11.5(SPSS,Chicago,IL,USA).Signi ficant means were further compared by Duncan’s multiple range tests at the 5%level.
Vector analysis was employed to evaluate the dynamics of nutrient conditions under different fertilization treatments.Nutrient concentration(c=a/m)is a product of nutrient uptake(a)and biomass accumulation(m),thus nutrient concentration change with time can be expressed by the function dc/dt=d(a/m)/dt(Imo and Timmer 1997).If the plant samples are taken at progressive intervals,then the initial status of the plants at t0(week 5 in this study)can be taken as the reference(normalized to 100)and can be compared with changes occurring at subsequent time intervals(t0to t1,t2,…).Plant responses were expressed relative to the reference and vector interpretation and possible diagnoses are given in Fig.1.
To quantify fertilization targets for growing B.alnoides seedlings,we used the conceptual model proposed by Timmer(1996)to distinguish nutrient de ficiency,suf ficiency,luxury consumption,suf ficiency and toxicity in plants.The principle behind this approach is that growth,N content and N concentration increase as N supply increases in the de ficiency range until luxury consumption,when N content and concentration continue to increase but no signi ficant change in total dry mass occurs.Nutrient suf ficiency is achieved when growth is the maximum,which is the transition between de ficiency and luxury consumption.Toxicity occurs when growth decreases,although N concentration increases due to accumulation effects(Salifu and Timmer 2003).This model can help quantify and de fine target rates for production of seedlings for field planting(Salifu and Jacobs 2006).
Results and discussion
Seedling growth dynamics
Root collar diameter,height and dry mass of B.alnoides seedlings in six fertilization treatments closely matched exponential functions over time(R2>0.95,P<0.01,Fig.2).At week 5 after transplanting,the difference was only signi ficant in the height among treatments(P<0.05,Fig.2b).As the nutrient supply increased,root collar diameter,dry mass and root and shoot ratio differed signi ficantly(P<0.05,Fig.2a,c,d),and largest root collar diameter,height and dry mass and smallest root and shoot ratio occurred in the 600E treatment at weeks 8 and 11 after transplanting(Fig.2).At the end of the experiment,root collar diameter,height and dry mass of six fertilization treatments were signi ficantly higher than those of the E0 treatment(P<0.05,Fig.2a–c),but according to growth indexes,the maximum values occurred in different treatments.For example,the maximum value was evident in 200E and 300E treatments for root collar diameter(Fig.2a);100E,200E and 300E treatments for seedling height(Fig.2b);and treatments from 100E to 600E for dry mass(Fig.2c).The root and shoot ratio declined signi ficantly with increasing nutrient application up to 200E,and no signi ficant difference occurred among treatments with higher fertilizer supply(Fig.2d).Therefore,as a whole,applications of 200 and 300 mg N seedling-1were adequate for growth of B.alnoides seedlings.
Nutrient uptake dynamics
The N,P and K contents of B.alnoides seedlings increased exponentially in the six fertilization treatments as time progressed(R2>0.95,P<0.01,Table 2),similar to that of seedling growth performance,inferring that exponential fertilization corresponded well to the increasing growth and nutrient uptake of B.alnoides seedlings.The N,P and K concentrations also increased over time except for the E0 treatment.This kind of positive correlation between nutrient concentration and nutrient supply under exponential fertilization was also detected in the study by Timmer et al.(1991)study on Picea mariana and by Imo and Timmer(1992)on Prosopis chilensis.A modi fied exponential fertilization is thus recommended to keep the nutrient concentration stable(Xu and Timmer 1998;Quoreshi and Timmer 2000).
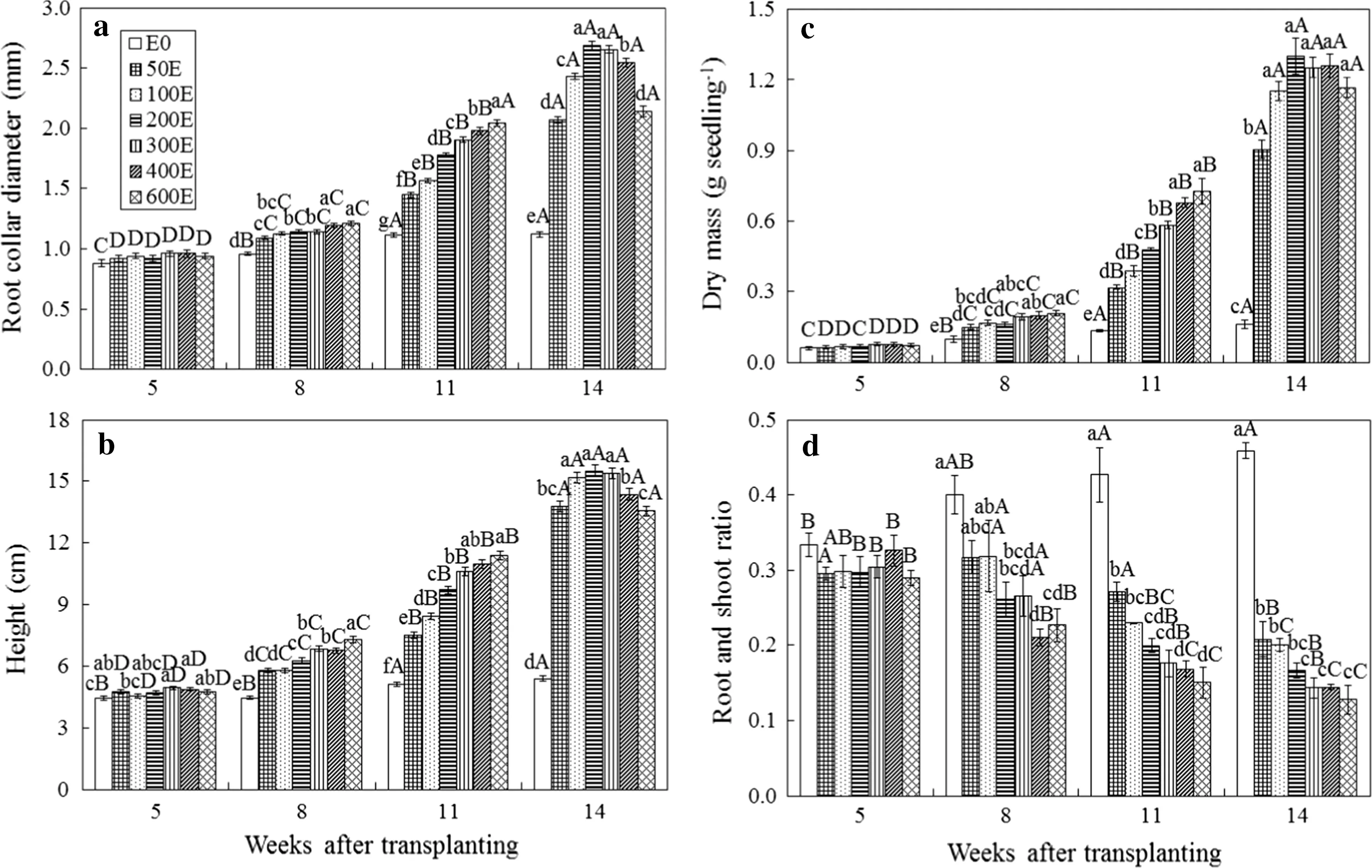
Fig.2 Growth variables of Betula alnoides seedlings at 5,8,11,14 weeks after transplanting during the greenhouse experiment.Paired treatments without the same lowercase and capital letters differed signi ficantly among treatments at each sampling time and in the same treatment over time according to Duncan’s multiple range tests at the 5%level.Error bars represent standard errors.E0,50E,100E,200E,300E,400E and 600E represent unfertilized treatment(control)and six exponential fertilization treatments as 50,100,200,300,400 and 600 mg N seedling-1
There were signi ficant differences in the N,P and K contents and concentrations among treatments at each survey and among surveysforthesametreatment(P<0.05,Table 2).By the end of the experiment,the N,P and K contents in seedlings had increased and remained stable at 300E,100E and 200E treatments,respectively(Table 2).These results showed that seedling requirements for these nutrient elements were quite different,and 300E,100E and 200E treatments were adequate for N,P and K demands of the seedlings,respectively.However,nutrient concentrations increased constantly with nutrient supply to maximum values in the 600E treatment(Table 2).Thus,application of 300 mg N seedling-1was suf ficient for nutrient uptake by B.alnoides seedlings.
Vector analysis
Dynamics of the nutrient status for each treatment was further characterized by vector analysis(Quoreshi and Timmer 2000).These progressions were expressed on a relative basis when the initial seedling status at week 5 after transplanting was normalized to 100 as the reference for comparison with samples collected at the subsequent 3 week intervals(Table 2;Fig.2)and were represented using vector nomograms(Fig.3).Phosphorus was the most responsive nutrient based on its largest vector length,followed by N and K(Fig.3).In the E0 plants,N and K were diluted(shift A,Fig.3a)associated with increased dry mass and nutrient contents but decreased nutrient concentrations,suggesting that the nutrient demand to sustain the high growth rate exceeded nutrient uptake(Imo and Timmer 1997).However,the dilutions of N and K induced initial suf ficiency of P(shift B,Fig.3a)characterized by increased dry mass and P content but no change in P concentration,and then luxury consumption of P(shift D,Fig.3a)associated with increase in P content and concentration but no signi ficant difference in dry mass as shown by the vector magnitudes.The dynamics of these nutrient elements were complex,which was due perhaps to antagonism between N and P(Chen et al.2010a).
Vector patterns signi fied de ficiencies in N and P(shift C,Fig.3b–g)under six exponential fertilization treatments,since dry mass,nutrient contents and concentrations increased,except that P in the 300E treatment exhibited suf ficiency(shift B,Fig.3e)at the end of the experiment.

Table 2 Nutrient conditions of Betula alnoides seedlings at 5,8,11,14 weeks after transplanting during the greenhouse experiment
These results demonstrated limitations of N and P in these seedlings because nutrient uptake rate was much less than the growth rate(Salifu and Jacobs 2006).Therefore,the best way to improve the nutrient uptake ef ficiency is an important issue for further studies by modifying exponential fertilization(Xu and Timmer 1998),adjusting the ratio of nutrient elements,inoculating with bene ficial mycorrhizal fungi(Gong et al.1999),and other strategies(Agathokleous et al.2016;Wang et al.2016).For K,initial dilution(shift A,Fig.3b)was evident in the 50E treatment,which was probably attributed to lower starting fertilizer rate and incomplete development of the root system(Imo and Timmer 1997),while initial suf ficiency of K(shift B,Fig.3c–g)was evident in higher exponential regimes.At the end of the experiment,the pattern changed to de ficiency of K(shift C,Fig.3c–g)in all exponential treatments,signifying K limitation was also evident in the seedlings.
Quantifying fertilizer regimes
The relationships among nutrient supply,dry mass,N content and N concentration of B.alnoides seedlings were divided into three phases to distinguish nutrient de ficiency,suf ficiency,luxury consumption,optimum and toxicity in the seedlings(Fig.4).As N supply increased from 0 to 100 mg seedling-1,dry mass,N contentand Nconcentration increased by more than 6,18 and 1 times,respectively(Table 2;Fig.2c),demonstrating N de ficiency in the seedlings.The N content and N concentration increased by 52 and 39%without a signi ficant increase in the dry mass as N supply increased from 100 to 400 mg seedling-1.When the N supply exceeded 400 mg seedling-1,N toxicity occurred in the 600E seedlings as N concentration increased by 8.2%,and a small number of seedlings withered and gradually died.Toxicity often decreases plant dry mass and nutrient content(Salifu and Timmer 2003).However,there were no signi ficant differences in the dry mass and N content between the 400E and 600E plants in this study,probably because the exclusion of dead plants would have overestimated the mean dry mass and N content.From the results,it could be determined that 100 and 400 mg N seedling-1were two crucial points,the former between de ficiency and luxury consumption,and the latter between luxury consumption and toxicity,de fining levels of suf ficiency and nutrient loading,respectively.
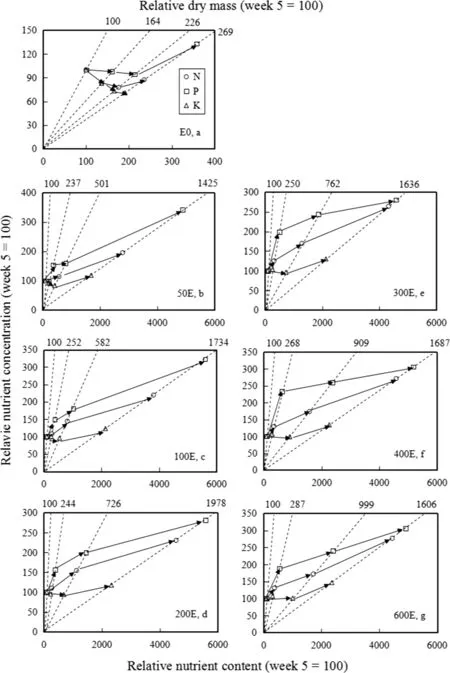
Fig.3 Progression of relative changes in dry mass,nutrient(N,P and K)contents and concentrations of Betula alnoides seedlings under E0,50E,100E,200E,300E,400E and 600E treatments(0,50,100,200,300,400,600 mg N seedling-1,respectively),sampled at weeks 5,8,11,and 14 after transplanting.The seedling status at week 5 of each treatment was normalized to 100.Vectors re flect response of individual treatments described in Fig.1

Fig.4 Responses of dry mass,N content,and N concentration of Betula alnoides seedlings to increasing N supply for one growing season(14 weeks)in the greenhouse.The vertical scale insert represents N concentration(%).Two vertical lines separate nutrient de ficiency,suf ficiency,luxury consumption,optimum and toxicity.Means followed by the same letter(a-c for biomass;u-z for N content;and f-k for N concentration)are not signi ficantly different at P<0.05 according to Duncan’s multiple range test for each variable.Fertilization followed exponential addition schedules.The lines for P and K were omitted since the seedling requirement for N(300 mg seedling-1)was higher than those of P(100 mg seedling-1)and K(200 mg seedling-1)at the end of the experiment(Table 2)
Although a broad range of nutrient supply(100–400 mg N seedling-1)was suitable for growing B.alnoides seedlings,400 mg N seedling-1can be recommended as the optimum target level for building seedling nutrient reserves byluxuryconsumption.Itispostulatedthatseedlingnutrient accumulation will improve early growth performance after outplanting,especiallyonsiteswithnutrientstressand fierce competition(Rytteretal.2003;Rikalaetal.2004;Heiskanen et al.2009;Basave Villalobos et al.2014).
Conclusions
Root collar diameter,height,dry mass and nutrient contents of B.alnoides seedlings in all nutrient addition treatments followed exponential functions over time.Maximum growth was achieved in 200E and 300E treatments,while treatments 300E,100E and 200E were adequate for nitrogen(N),phosphorus(P)and potassium(K)demands of the seedlings,respectively.Vector analysis showed that dilutions of N and K,and P suf ficiency or luxury consumption were evident in the E0 plants probably due to nutrient interaction between N and P.However,de ficiencies of N,P and K were observed in all exponential fertilization treatments at the end of the experiment,suggesting that these nutrients limited seedling growth because growth rate exceeded nutrient uptake rate.The 100E and 400E treatments were the two critical points distinguishing nutrient de ficiency,luxury consumption and toxicity based on a nutrient loading model and the 400E treatment was recommended to apply for nutrient loading of seedlings before outplanting.
AcknowledgementsThe authors were grateful to Mrs.Kai-Qin LIN and Mr.Er SHA for their assistances in the nursery managements,and to Mrs.Le-Su YANG and Bin YU for the nutrient determinations.
Agathokleous E,Saitanis CJ,Wang X,Watanabe M,Koike T(2016)A review study on past 40 years of research on effects of tropospheric O3on belowground structure,functioning,and processes of trees:a linkage with potential ecological implications.Water Air Soil Pollut 227(1):33
Basave Villalobos E,Cetina AlcaláVM,López López MA,Aldrete A,Del Valle Paniagua DH(2014)Nursery practices increase seedling performance on nutrient-poor soils in Swietenia humilis.iForest-Biogeosciences For 8(4):552
Chen L,Zeng J,Xu DP,Zhao ZG,Guo JJ(2010a)Macronutrient de ficiency symptoms in Betula alnoides seedlings.J Trop For Sci 22:403–413
Chen L,Zeng J,Xu DP,Zhao ZG,Guo JJ,Lin KQ,Sha E(2010b)Effects of exponential nitrogen loading on growth and foliar nutrient status of Betula alnoides seedlings.Sci Sil Sin 46(5):35–40(in Chinese)
Chen L,Zeng J,Jia HY,Zeng J,Guo WF,Cai DX(2012)Growth and nutrient uptake dynamics of Mytilaria laosensis seedlings under exponential and conventional fertilizations.Soil Sci Plant Nutr 58(5):618–626
Duan J,Xu C,Jacobs DF,Ma L,Wei H,Jiang L,Ren J(2013)Exponential nutrient loading shortens the cultural period of Larix olgensis seedlings.Scand J For Res 28(5):409–418
Gong M,Wang F,Chen Y,Chen Y(1999)Mycorrhizal dependency and inoculant effects on growth of Betula alnoides seedlings.For Res 13(1):8–14(in Chinese)
Haase DL,Rose R(1995)Vector analysis and its use for interpreting plant nutrient shifts in response to silvicultural treatments.For Sci 41(1):54–66
Heiskanen J,Lahti M,Luoranen J,Rikala R(2009)Nutrient loading has a transitory effect on the nitrogen status and growth of outplanted Norway spruce seedlings.Silva Fenn 43(2):249–260
Imo M,Timmer VR(1992)Nitrogen uptake of mesquite seedlings at conventional and exponential fertilization schedules.Soil Sci Soc Am J 56(3):927–934
Imo M,Timmer VR(1997)Vector diagnosis of nutrient dynamics in mesquite seedlings.For Sci 43(2):268–273
Jiang Y,Wang D,Qiu Q,Yang D,Xu L,Zhao Y,Zhang K,Li Y,Zhao H(2003)Fertilization experiment on young plants of seven broadleaved tree species indigenous to tropical areas.Yunnan For Sci Tech 103:11–16(in Chinese)
Quoreshi M,Timmer VR(2000)Growth,nutrient dynamics,and ectomycorrhizal development of container-grown Picea mariana seedlings in response to exponential nutrient loading.Can J For Res 30(2):191–201
Rikala R,Heiskanen J,Lahti M(2004)Autumn fertilization in the nursery affects growth of Picea abies container seedlings after transplanting.Scand J For Res 19(5):409–414
Rytter L,Ericsson T,Rytter RM(2003)Effects of demand-driven fertilization on nutrient use,root:plant ratio and field performance of Betula pendula and Picea abies.Scand J For Res 18(5):401–415
Salifu KF,Jacobs DF(2006)Characterizing fertility targets and multi-element interactions in nursery culture of Quercus rubra seedlings.Ann For Sci 63(3):231–237
Salifu KF,Timmer VR(2003)Optimizing nitrogen loading of Picea mariana seedlings during nursery culture.Can J For Res 33(7):1287–1294
Salifu KF,Jacobs DF,Birge ZKD(2008)Performance of nutrientloaded red oak and white oak seedlings on mine lands in southern Indiana.In:Dumroese RK,Riley LE(Technical coordinators)National proceedings:forest and conservation nursery associations-2007.Fort Collins(CO):USDA Forest Service,Rocky Mountain Research Station.Proceedings RMRSP-57,p 65–71
Timmer VR(1996)Exponential nutrient loading:a new fertilization technique to improve seedling performance on competitive sites.New For 13(1):279–299
Timmer VR,Armstrong G(1989)Growth and nutrition of containerized Pinus resinosa seedlings at varying moisture regimes.New For 3(2):171–180
Timmer VR,Armstrong G,Miller BD(1991)Steady-state nutrient preconditioning and early outplanting performance of containerized black spruce seedlings.Can J For Res 21(5):585–594
Van den Driessche R(1991)Effects of nutrients on stock performance in the forest.In:Van den Driessche R(ed)Mineral nutritional in conifer seedlings.CRC Press,Florida,pp 229–260
Wang J,Li G,Pinto JR,Liu J,Shi W,Liu Y(2015)Both nursery and field performance determine suitable nitrogen supply of nurserygrown,exponentially fertilized Chinesepine.SilvaFenn 49(3):1–13
Wang X,Agathokleous E,Qu L,Watanabe M,Koike T(2016)Effects of CO2and O3on the interaction between root of woody plants and ectomycorrhizae.J Agric Meteo 72(2):95–105
Xu XJ,Timmer VR(1998)Biomass and nutrient dynamics of Chinese fir seedlings under conventional and exponential fertilization regimes.Plant Soil 203(2):313–322
Zeng J,Guo WF,Zhao ZG,Weng QJ,Yin GT,Zheng HS(2006)Domestication of Betula alnoides in China:current status and perspectives.For Res 19(3):379–384(in Chinese)
 Journal of Forestry Research2018年1期
Journal of Forestry Research2018年1期
- Journal of Forestry Research的其它文章
- Vascular bundle connection between seed stalk and seed coat of Caragana arborescens
- Mini-cutting technique for Khaya anthotheca:selection of suitable IBA concentration and nutrient solution for its vegetative propagation
- Reconstructing the size of individual trees using log data from cut-to-length harvesters in Pinus radiata plantations:a case study in NSW,Australia
- Phenotypic variation in Phoebe bournei populations preserved in the primary distribution area
- Effects of soil drought stress on photosynthetic gas exchange traits and chlorophyll fluorescence in Forsythia suspensa
- Flavonoid content and radical scavenging activity in fruits of Chinese dwarf cherry(Cerasus humilis)genotypes