
干旱胁迫条件下贵阳市引种5种柑橘砧木的抗旱性比较
2018-03-27 01:36杨雪莲
江苏农业科学 2018年4期
杨雪莲
(贵州大学,贵州贵阳 550025)
柑橘(Citrusreticulata)属芸香科柑橘属柑橘种植物,是我国南方的重要果树之一,但柑橘大多种植于丘陵、山地或海涂,季节性干旱往往给柑橘的生长发育和产量品质造成极为不利的影响[1],因此,筛选或培育抗旱性强而又高产的柑橘种质资源尤为重要。砧木是嫁接果树的基础,对接穗品种的生长发育及果实产量和品质等均有重要影响。我国柑橘砧木资源丰富,在广西也有较多柑橘砧木地方品种和变异类型,对当地的气候和土壤有良好的适应性。全球气候变化与局部干旱化将导致越来越多的干旱、半干旱地区受到更为严重的干旱胁迫影响[2]。在不同干旱条件下植物叶片内的超氧化物歧化酶(SOD)、过氧化物酶(POD)、游离脯氨酸、可溶性糖、丙二醛、电导率等生理生化指标的变化,可以用于研究这些因子在干旱胁迫时发生变化的规律。干旱胁迫处理在蓝莓、李、桃、梨和杏方面已有比较广泛的应用[3-4]。目前,我国关于干旱胁迫在柑橘方面的相关研究还很少。
在柑橘品种抗旱性研究上,聂华堂等对柑橘抗旱性与生理生化指标的变化关系曾有过报道,认为它们在干旱胁迫时可溶性蛋白质含量的累积及叶片中POD活性的增加率与柑橘品种的抗旱力强弱呈正相关[5]。甘海峰等在柑橘砧木的干旱胁迫的研究中指出,干旱胁迫对山东枳壳、宁明橘、阳朔金宝酸橘、滑皮金橘、桂枳1号5个供试品种叶片中抗氧化酶系统SOD、POD活性以及对蛋白质、MDA含量均产生了影响。SOD、POD活性均有不同程度的升高,可溶性蛋白质含量均有不同程度的降低,且干旱胁迫显著提高了5个柑橘品种叶片中MDA的含量[2]。砧木的抗旱性对柑橘树体抗旱力强弱有十分重要的影响,因此,对不同柑橘砧木抗旱性的研究,对促进我国柑橘发展有重大意义。
1 材料与方法
1.1 材料及处理
以厚皮枳、皱皮枳、枳橙、77-1、锦橙2年实生幼苗为试验材料。处理组于2015年3月1日进行充分浇水后让其做避雨干旱处理,对照组放置于温室外进行正常日常管理。随后每隔5 d分别对干旱胁迫和对照组取其叶片进行生理生化指标测定1次。
1.2 测定指标及方法
丙二醛(MDA)、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、脯氨酸含量、可溶性糖含量、蛋白质含量的测定参考李合生的《植物生理生化实验原理和技术》[6]。
1.3 柑橘砧木抗旱性比较

1.4 数据分析
使用Excel表格进行数据统计和图表分析。
2 结果与分析
2.1 天气状况
试验于2015年3月在贵州大学农学院开展,对照在试验阶段的生长环境见表1。从表1可以看出,贵阳市花溪区春季多雨,温度较低,温差相对较大。在此天气状况下对5个柑橘砧木材料进行干旱胁迫。3月基本上每天都有小雨,雨水充沛,对置于室外的对照,即使不浇水,靠雨水也能够维持正常的生理活动。试验期间风力均为微风。
2.2 可溶性蛋白含量的动态变化
可溶性蛋白是植物生命活动的体现,不同植物可溶性蛋白的含量有所不同。植物体内的可溶性蛋白大多是参与各类代谢的酶类,在植物受到干旱胁迫时,可溶性蛋白含量会发生一定的变化,可溶性蛋白含量越高抗旱性越强,测定其含量变化是了解植物抗旱性的重要指标之一[7]。
从图1-A可以看出,77-1、锦橙、皱皮枳的可溶性蛋白含量都在5~10 d下降,10~20 d呈现上升趋势;其中,厚皮枳可溶性蛋白含量一直处于上升状态;枳橙是在5~10 d呈现缓慢上升后下降,以锦橙、77-1可溶性蛋白含量上升最为明显。
从图1-B可以看出,对照组的不同品种蛋白质含量波动不大。品种间比较厚皮枳可溶性蛋白含量最低,在干旱胁迫阶段呈现逐渐上升的趋势;皱皮枳蛋白质含量相对较高,且呈现先上升后下降再上升变化趋势;77-1、锦橙可溶性蛋白变化趋势相同,均呈现先下降后上升趋势,但锦橙可溶性蛋白含量高于77-1。图1-A、图1-B比较可知,在大棚内的植株对干旱胁迫表现较为突出,可溶性蛋白随时间的延长波动较大,表明春季遇到干旱对柑橘砧木生长有较大影响。
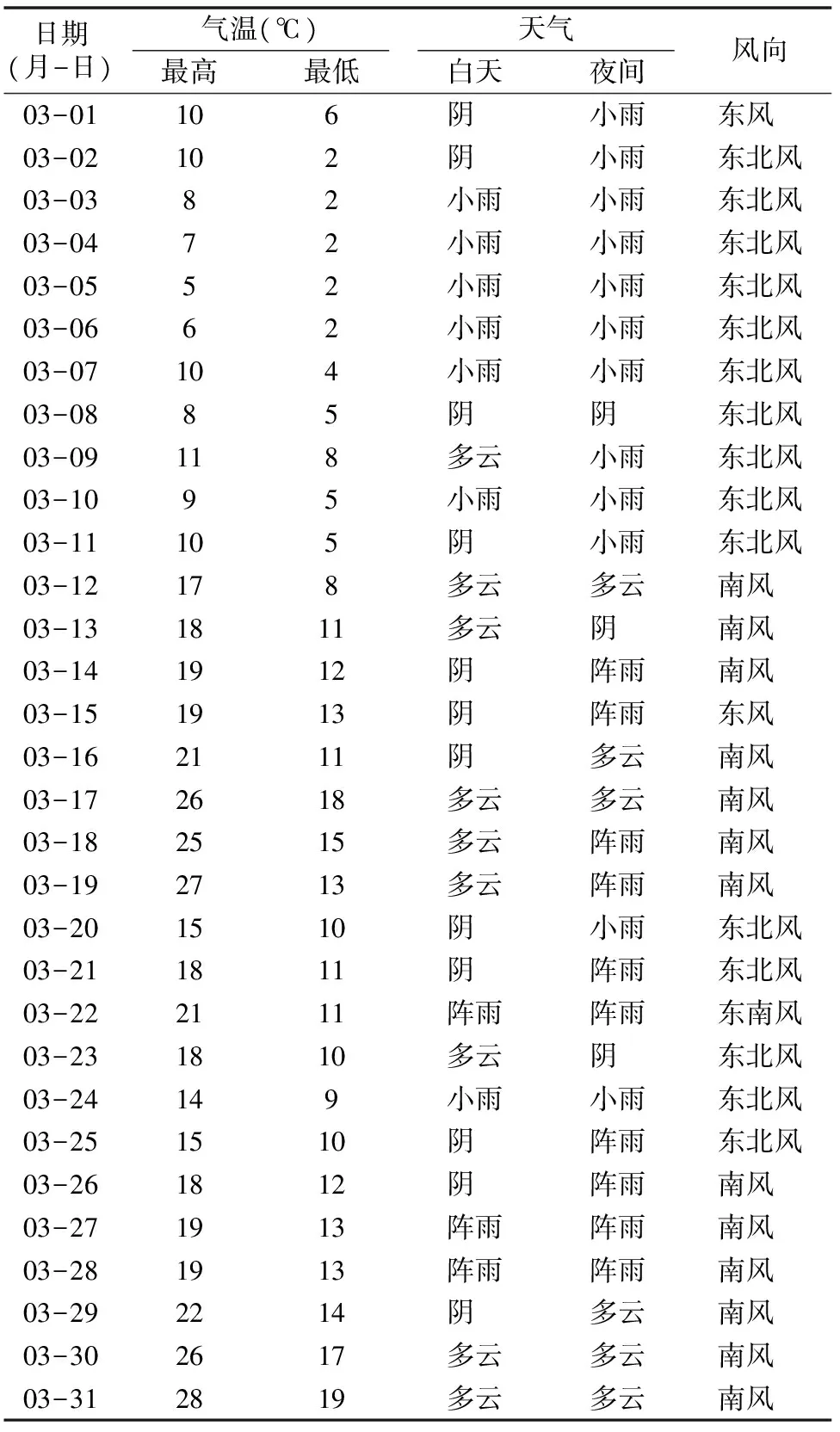
表1 2015年3月贵州市花溪区天气状况
2.3 可溶性糖含量的动态变化
可溶性糖含量不仅是植物体内重要的能量来源,还是植物体内重要的渗透调节物质[8]。从图2-A可以看出,厚皮枳、皱皮枳、枳橙、77-1、锦橙中的可溶性糖含量都是随着干旱胁迫时间的增加呈现先下降再上升的趋势,但其中以皱皮枳、枳橙可溶性糖含量上升趋势最为明显而且大于其他几个品种。从图2-B可以看出,在自然条件下厚皮枳、皱皮枳、枳橙、77-1、锦橙中的可溶性糖含量虽然都在缓慢的上升,但上升趋势都不大。从图2-A、图2-B可以看出,经过温室内干旱胁迫处理的可溶性糖含量随着干旱胁迫时间的增加明显高于在温室外自然条件下的可溶性糖含量。
2.4 过氧化物酶(POD)活性的动态变化
POD可以清除掉植物体内过多的H2O2等过氧化物,并且是细胞内防御酶系统中自由基的清除剂。不同品种POD活性变化会随着干旱胁迫时间的增加而有所不同[9]。
从图3-A可以看出,不同柑橘砧木品种的POD活性都呈现出稳定上升趋势,其中以皱皮枳、枳橙上升趋势最为明显,POD活性明显高于其他3个品种。从图3-B可以看出,5个品种中POD活性的变化基本维持不变。从图3-A、图3-B 综合可以看出, 随着干旱胁迫天数的增加, 干旱胁迫处理不同品种中的POD活性都会高于自然条件下的品种。



2.5 超氧化物歧化酶(SOD)活性的动态变化
SOD是植物体内重要的抗氧化保护酶,SOD活性能直接影响着活性氧的清除,而细胞对活性氧的清除能力越强,越能避免植物体内过度积累的活性氧对细胞膜和细胞功能的伤害[9]。
从图4-A可以看出,不同品种SOD活性都是随着干旱胁迫时间的增加呈现先上升后下降的趋势,其中皱皮枳、枳橙的SOD活性明显高于其他3个品种。从图4-B可以看出,随着干旱胁迫天数的增加,不同品种SOD活性没有明显变化。

2.6 丙二醛(MDA)含量的动态变化
MDA是膜脂过氧化的主要产物之一,植物体内MDA含量的多少可以直接反映植物受害程度,随着干旱胁迫时间的增加丙二醛含量也会有所增加,但丙二醛含量越多植物的抗旱性会越弱[10-11]。
从图5-A可以看出,随着干旱胁迫时间的增加,厚皮枳、皱皮枳、枳橙、77-1、锦橙MDA含量有不同程度的上升,其中厚皮枳、77-1上升趋势最为明显,随着干旱胁迫时间的增加厚皮枳、77-1、锦橙的MDA含量都高于皱皮枳、枳橙。从图5-B可以看出,在自然条件下几个柑橘砧木品种MDA含量都是基本维持不变。从图5-A、图5-B综合可以看出,随着干旱胁迫时间的增加,干旱胁迫处理中MDA含量都高于自然条件的对照。

2.7 脯氨酸含量的动态变化
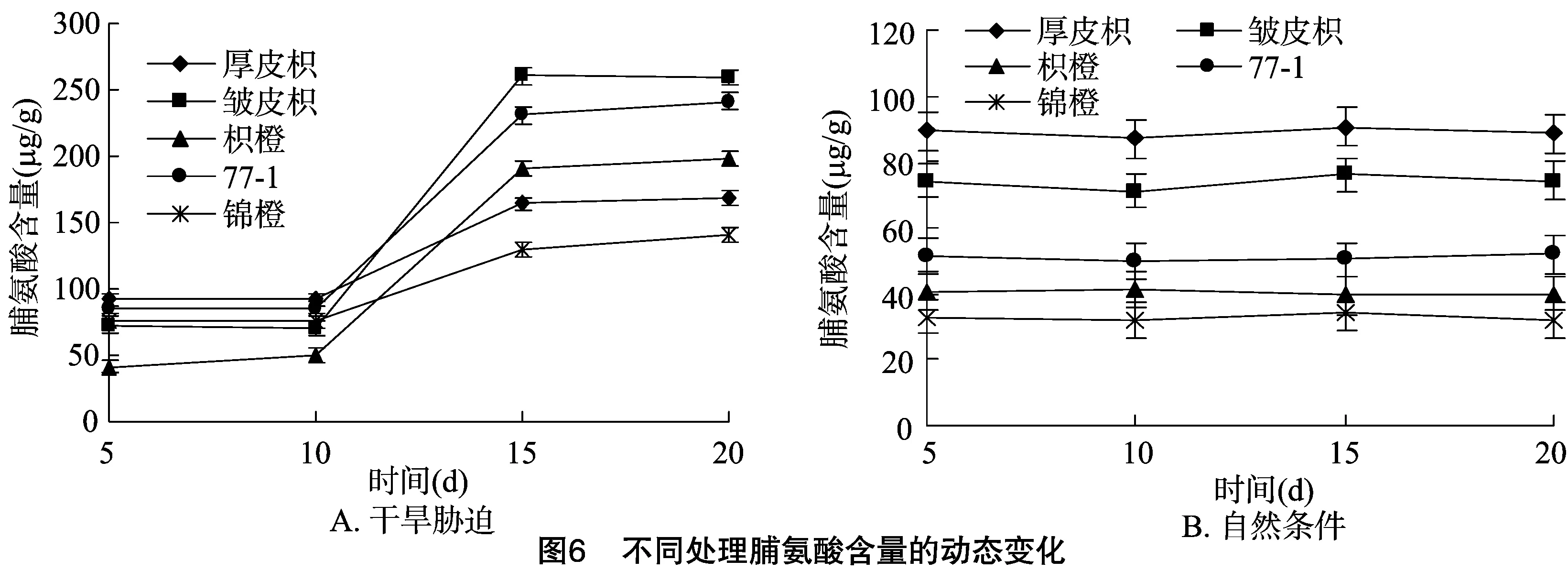
干旱胁迫下,植物细胞内能快速合成一些小分子渗透调节物质,以达到保持细胞膨压,维持细胞正常生理功能的目的。脯氨酸是较为普遍的渗透调节物质之一,在干旱胁迫期间不同品种柑橘砧木的脯氨酸含量都有所增加[12-13]。
从图6-A可以看出,随着干旱胁迫时间的增加,厚皮枳、皱皮枳、枳橙、77-1、锦橙的脯氨酸含量都有所增加,不同品种在5~10 d脯氨酸含量都基本维持不变,10 d以后不同品种的脯氨酸含量都有明显上升,其中以皱皮枳、枳橙上升最为明显。从图6-B可以看出,随着干旱胁迫时间的增加,在自然条件下的对照不同品种的脯氨酸含量基本维持不变。
2.8 柑橘砧木抗旱性综合比较
以不同柑橘砧木在干旱胁迫下测定各指标最终进行模糊统计中隶属函数分析加权平均值作为柑橘砧木抗旱性综合比较标准。
从表2可以看出,皱皮枳的平均隶属值最大,表明抗旱性最强;77-1的平均隶属值最小,表明抗旱性最弱。分析结果,不同柑橘砧木材料抗旱性依次表现为:皱皮枳>厚皮枳>枳橙>锦橙>77-1。

表2 柑橘砧木抗旱性指标隶属函数值及综合比较
为进一步研究5个柑橘砧木材料抗旱性指标间的协同变异程度,对上述材料的可溶性蛋白质含量、可溶性糖含量、SOD活性、POD活性、丙二醛、脯氨酸含量进行相关性分析,从表3可以看出,平均隶属值与可溶性糖、SOD活性、丙二醛之间都呈显著水平。从表4可知,平均隶属值与可溶性糖SOD活性、丙二醛的决定系数都在0.5以上,表明可以用平均隶属值的大小来比较柑橘砧木材料抗旱性的强弱[14]。

表3 不同柑橘砧木材料生理指标间的相关性分析
注:A1—可溶性蛋白质;A2—可溶性糖;A3—超氧化物歧化酶;A4—过氧化物酶;A5—丙二醛;A6—脯氨酸;A7—平均隶属值;“**”为1%水平显著相关;“*”为5%水平显著相关。表4同。

表4 不同柑橘砧木材料生理指标间的相关性分析
3 讨论与结论
干旱胁迫处理会引起植物体内多种生理生化指标发生不同程度的变化,本试验结果表明,在干旱胁迫下柑橘砧木的可溶性蛋白质、可溶性糖、SOD、POD、脯氨酸、丙二醛含量都有不同程度的影响。
为了有效提高植物渗透调节能力,植物会在干旱胁迫下积累渗透调节物质,以增强植物的抗旱性。植物中参与渗透调节的有机物包括可溶性蛋白质、可溶性糖、脯氨酸等,而可溶性糖含量、可溶性蛋白含量、脯氨酸含量的增加有利于植物抗旱性的提升[15]。本试验中不同柑橘砧木材料的可溶性糖含量都呈现先缓慢下降后再上升的趋势,其中皱皮枳、枳橙上升趋势最明显,皱皮枳含量高于枳橙,表明皱皮枳抗旱性要强于其他几个品种。可溶性蛋白含量在干旱胁迫处理下总体呈现上升趋势,可以推断出柑橘砧木在受到干旱胁迫时以提高可溶性蛋白质、可溶性糖含量来提高其抗旱力。脯氨酸含量在干旱胁迫过程也随着干旱胁迫时间增加而提高,从而提高了柑橘砧木的渗透调节能力;其中以皱皮枳和枳橙的提高最为明显,皱皮枳又明显高于枳橙,表明皱皮枳的抗旱能力最高。
SOD、POD是柑橘砧木体内的保护酶,主要作用是清除柑橘砧木体内的活性氧、自由基和过氧化物。柑橘砧木在受到干旱胁迫等逆境时体内的活性氧和自由基等物质会增加,从而对植物体内的细胞膜和细胞功能造成伤害,保护性酶在保持植物正常发育的过程中起到关键作用。前人研究结果,保护性酶与植物的抗氧化能力成正比,是评价植物抗逆性的重要标志。本试验中SOD、POD活性会随着干旱胁迫时间的延长呈现上升的趋势,表明柑橘砧木在受到干旱胁迫时会提高体内的SOD、POD活性,从而增强柑橘砧木清除活性氧、自由基等伤害植物细胞膜和细胞功能物质的能力。
MDA与蛋白质氨基酸残基反应生成的产物会降低细胞膜的稳定性,导致膜的渗漏,增大细胞膜的选择透性,使电解质外渗,破坏叶绿体膜、线粒体膜等亚细胞器膜的功能和结构,导致生理功能的紊乱,MDA含量增加会加重细胞膜的伤害。本试验中柑橘砧木在干旱胁迫处理下不同品种MDA含量都呈现逐渐上升趋势,其中以厚皮枳、77-1、锦橙的含量上升最快,而皱皮枳MDA含量略低于枳橙的MDA量。
本试验以厚皮枳、皱皮枳、枳橙、77-1、锦橙为试验材料,在干旱胁迫下分别在5、10、15、20 d时测定其可溶性蛋白质含量、可溶性糖含量、超氧化物歧化酶活性、过氧化物酶活性、丙二醛含量、脯氨酸含量,对其含量采用模糊统计分析中隶属函数分析其隶属函数平均值作为柑橘砧木抗旱性综合比较标准。通过比较得出不同柑橘砧木材料的抗旱性依次表现为皱皮枳>厚皮枳>枳橙>锦橙>77-1。
[1]陈仕俏,赵文红,白卫东. 我国柑橘的发展现状与展望[J]. 农产品加工·学刊,2008(3):21-22.
[2]甘海峰,傅翠娜,雷新南,等. 干旱胁迫对不同柑橘砧木叶片抗氧化系统的影响[J]. 安徽农业科学,2010,38(35):19983-19986.
[3]曾玮玮. 蓝莓对干旱胁迫的生理响应研究[D]. 金华:浙江师范大学,2012.
[4]胡学华. 李、桃、杏和梨四种经济树木对干旱胁迫的生理响应及其抗旱性的研究[D]. 雅安:四川农业大学,2005.
[5]聂华堂,陈竹生,计 玉. 水分胁迫下柑橘的生理变化与抗旱性关系[J]. 中国农业科学,1991,24(4):14-18.
[6]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[7]蒲光兰,袁大刚,胡学华,等. 土壤干旱胁迫对3个杏树品种生理生化特性的影响[J]. 浙江林学院学报,2005,22(4):375-379.
[8]廖景容,郑中阳,张卫星,等. 玉米不同杂交组合抗旱性、丰产性的研究[J]. 安徽农业科学,2005,33(1):7-10.
[9]徐启贺. 苹果砧木对干旱胁迫的生理响应及抗旱性评价[D]. 北京:中国农业科学院,2010.
[10]史玉炜,王燕凌,李文兵,等. 水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响[J]. 新疆农业大学学报,2007,30(2):5-8.
[11]陆云梅. 水杨酸对干旱胁迫下柑橘生理生化特性的影响[D]. 武汉:华中农业大学,2007.
[12]罗 音,孙明高. 干旱胁迫对5树种叶片中脯氨酸含量的影响[J]. 山东林业科技,1999(4):1-4.
[13]向碧霞,成明昊,季晓林,等. 苹果砧木资源的抗旱性研究[J]. 西南大学学报(自然科学版),1995,17(5):381-385.
[14]张 静,赵亮明,邹志荣. 不同苹果砧木组培苗抗旱性的比较研究[J]. 果树学报,2013,30(1):88-93.
[15]周 玲,王乃江,张丽楠. PEG胁迫对文冠果种子萌发和幼苗生理特性的影响[J]. 西北植物学报,2012,32(11):2293-2298.
猜你喜欢
中老年保健(2022年3期)2022-11-21
落叶果树(2021年6期)2021-02-12
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
现代园艺(2017年21期)2018-01-03
浙江农业学报(2017年1期)2017-05-17
湖南农业(2017年1期)2017-03-20
湖南林业科技(2017年1期)2017-02-06
西南农业学报(2016年4期)2016-05-17
山西农经(2015年7期)2015-07-10
