
不同水分条件下小麦回交导入系群体旗叶持绿性与千粒重的遗传相关分析
2018-03-20 00:53辛筱筱栗孟飞程宏波陈菁菁柴守玺杨德龙
干旱地区农业研究 2018年1期
辛筱筱,栗孟飞,刘 媛,程宏波,常 磊,陈菁菁,柴守玺,杨德龙
(1.甘肃省干旱生境作物学重点实验室,甘肃农业大学生命科学技术学院, 甘肃 兰州 730070;2.甘肃农业大学农学院, 甘肃 兰州 730070)
小麦(TriticumaestivumL.)是我国四大粮食作物之一,常年播种面积约2.67×107hm2,占粮食作物播种总面积的27.0%左右,占粮食作物总产量的22.0%左右[1]。随着农业用水资源的日益匮乏,干旱已成为小麦减产的最主要的环境影响因子,每年我国因不同程度干旱导致的小麦减产在30%以上,其危害相当于其他自然灾害的总和[2]。研究表明,小麦产量形成是源-流-库协同互作的复杂生理生化过程,主要取决于功能叶片的持绿性[3]。尤其是,小麦生育后期(抽穗期至灌浆期)是其产量形成的关键时期,同时也是对水分响应极为敏感的时期,该时期旗叶已成为小麦最为主要的光合器官[3-4]。在此条件下,小麦旗叶保持较高的叶绿素含量和一定的绿叶面积往往具有较高的产量潜势,表现出较强的抗旱性[4],这种性状有利于植株延缓叶片衰老,改善蒸腾效率和延长光合作用时间[5],从而有效地促进干物质积累、转运和籽粒灌浆,提高粒重和产量[6]。因此,解析干旱胁迫条件下小麦生育后期旗叶持绿性与粒重关系及其遗传特征对提高小麦抗旱遗传改良效率具有重要意义。
小麦持绿性是一个复杂的生理代谢过程[7]。现已证明,叶绿素合成主要受谷氨酰tRNA还原酶、胆色素原合酶、尿叶琳原Ⅲ合酶、NADPH原叶绿素酸醋氧化还原酶、叶绿素合酶、叶绿素酸醋a氧化酶等关键酶及其基因表达调控[8];且其表型易受基因型、环境、基因型×环境互作等因子的显著影响[9-11]。借助分子数量遗传学手段发现,小麦持绿性和千粒重属于多基因控制的复杂数量性状,遗传力低,易受水分环境影响[9,12-13]。前人利用初级作图群体,如重组近交系群体(recombinant inbred line,RIL)[11-12]、加倍单倍体群体(doubled haploid line,DH)[13]等,对小麦持绿性和千粒重进行数量性状位点(quantitative trait locus,QTL)定位发现,控制这些性状的QTL在小麦整个基因组上均有分布,其遗传受加性、上位性及其与水分环境互作效应控制,且不同的环境和遗传背景数量遗传基础有显著差异。因此,利用不同背景小麦材料,设置不同的水分处理,有利于挖掘更多控制这些性状的抗旱遗传信息。
小麦回交导入系(introgression lines,ILs)群体的建立为小麦基因从初级定位向精细定位发展奠定了良好的材料基础。通过多次回交建立的小麦ILs群体表现高比率的轮回亲本基因型,同时保留少量供体亲本染色体片段(基因),有效排除了不同遗传背景对导入基因的干扰,是检测目标性状QTL/基因,进行基因精细定位与克隆的理想遗传材料。施伟等[14]利用晋麦47和长6878分别作为供体亲本,鲁麦14作为轮回亲本,分别构建了两套ILs群体,并对其进行了抗旱性相关性状QTL定位分析。陈稳良等[15]以晋麦47为轮回亲本,鲁麦14为供体亲本,构建ILs群体,通过分子标记技术检测,对不同水分条件下小麦抗旱相关重要性状和生理性状进行了QTL定位。Ibrahim等[16]利用2套小麦ILs群体(Triso×Syn084和Devon×Syn084)对小麦抗旱相关性状进行了精细定位。以上研究为揭示小麦抗旱性相关复杂数量性状的遗传特性和抗旱相关性状精细定位奠定了良好的材料基础和理论依据。因此,本研究以西峰20×晋麦47杂交创建的ILs群体作为供试材料,研究干旱胁迫和灌溉条件下不同发育时期小麦旗叶叶绿素含量和千粒重相关性及其数量遗传特性,并对ILs群体的遗传变异进行评价,旨在为小麦抗旱性研究和遗传改良提供理论基础。
1 材料与方法
1.1 供试材料
以强抗旱性的冬小麦西峰20为供体亲本,以抗旱高产品种晋麦47为轮回亲本,创建的ILs(BC3F4)群体160个株系为供试材料,群体两亲本在抗旱性和千粒重上遗传差异较大。其中,供体亲本西峰20是以西峰18作为母本,CA8055作为父本,通过有性杂交、分离世代的单株选择,育成的具有抗旱、抗寒、抗病性强的新品种。轮回亲本晋麦47是以12057为母本,(522×K37-20)为父本,经杂交,采用后裔育种选育而成,高产,抗寒耐冻性好,分蘖较强,灌浆速度快,现为国家黄淮旱地、山西省旱地、陕西省旱地区试的对照品种。
试验于2013年10月—2014年6月在甘肃省兰州市榆中县小麦试验点(35°51′N,104°07′E,平均海拔1 900 m,平均气温6.6℃,年降雨量450 mm,年蒸发量1 450 mm,无霜期140 d)进行。小麦播前基肥施用量为纯N 180 kg·hm-2,P2O5120 kg·hm-2,K2O 75 kg·hm-2,在整个生育期内均不再施肥。田间试验水分管理分为雨养,即干旱胁迫(drought stress,DS)和灌溉(well watered,WW)两种处理,播前均统一灌底墒水900 m3·hm-2。灌溉处理按正常的田间水分管理进行,分别在越冬前、翌年拔节期和开花期补充灌水,每次灌水量均为600 m3·hm-2。雨养的试验材料仅越冬前灌水750 m3·hm-2,其余生育期依靠自然降水,全生育期(播种至收获)降水量为103 mm。以上试验为随机区组设计,3次重复,6行区,均为稀条播,行长1 m,行距0.2 m,每行点播小麦60粒。
1.2 试验方法
1.2.1 目标性状的测定 在小麦的抽穗期、开花期和灌浆期,利用便携式叶绿素测定仪(SPAD-502Plus,日本)测定不同水分条件下ILs群体亲本和各株系的旗叶叶绿素含量(ChlC),分别记为ChlCh,ChlCf和ChlCg。在小麦收获脱粒后,风干种子,测定各株系及其亲本的千粒重(TGW)。以上表型测定均5次重复,求平均值,计算干旱胁迫系数(drought stress coefficient,DSC),即干旱胁迫下表型值与灌溉条件下表型值的比值(DS/WW),代表材料的抗旱性[17]。

2 结果与分析
2.1 小麦ILs群体与其亲本叶绿素含量和千粒重的表型分析
在两种水分条件下,小麦ILs群体两亲本不同发育阶段旗叶ChlC和TGW表型值均呈显著或极显著差异(表1)。供体亲本西峰20不同发育时期旗叶ChlC表型值显著高于轮回亲本晋麦47,而TGW表型值反之。ILs群体各性状的表型值均介于两亲本之间,群体内各株系变异广泛,存在超亲分离现象,不同发育时期旗叶ChlC的变异系数在9.89%~16.71%(DS)和11.4%~16.54%(WW)之间;TGW的变异系数分别为8.39%(DS)和8.61%(WW)。表明,小麦ILs 群体双亲对所考察性状有贡献的等位基因在后代群体中得到广泛分离,呈现出多基因控制的数量性状特点,且目标性状表型多偏于轮回亲本晋麦47,体现出导入系群体的遗传特性。同时,小麦ILs 群体所有目标性状偏度和峰度系数的绝对值普遍小于1,表型符合正态分布,表明这些性状系典型的多基因控制的复杂数量性状。
小麦ILs群体及其双亲WW条件下的目标性状表型值显著高于DS条件下(表1),各目标性状的旱胁迫系数在0.79~0.91之间(图1)。其中,西峰20的旗叶ChlC旱胁迫系数随着生育期的推移稳定增大,千粒重的旱胁迫系数较高(0.90)。晋麦47的旗叶ChlC旱胁迫系数随着生育期的推移而下降,千粒重的旱胁迫系数较低(0.87)。小麦ILs群体的旗叶ChlC旱胁迫系数在开花期最高(0.91),千粒重的旱胁迫系数介于双亲之间(0.89)。由此看出,在个体和群体水平上反映出小麦旗叶ChlC和TGW的表型变异易受水分环境的影响。
2.2 小麦ILs群体叶绿素和千粒重的相关分析
从表2看出,在不同水分环境条件下,小麦ILs群体不同发育时期旗叶ChlC间呈不同程度的正相关(r=0.256~0.505**,DS;0.312*~0.429**,WW)。而ChlC与TGW间呈现极显著正相关(r=0.500**~0.629**,DS;0.488**~0.622**,WW),其中灌浆期的旗叶ChlC与TGW表现出较高的相关性(r=0.629**,DS;0.622**,WW),且DS条件下的相关系数普遍高于WW。说明在DS条件下,小麦灌浆期维持较高的叶绿素含量对千粒重具有重要作用。

表1 不同水分条件下小麦ILs及其亲本的性状表型值 Table 1 Phenotypes of the ILs and its parents under different water conditions
注:ChlCh, ChlCf和ChlCg分别表示抽穗期、开花期和灌浆期旗叶叶绿素含量;TGM表示千粒重,DS和WW分别表示干旱胁迫和灌溉处理。*P<0.05; **P<0.01.下同
Note: ChlCh, ChlCf and ChlCg mean chlorophyll content of wheat flag leaves at heading, flowering, and grain-filling stages, respectively; TGW means thousand-grain weight; DS and WW mean drought-stress and well-watered treatment, respectively. *P<0.05; **P<0.01. The same below.
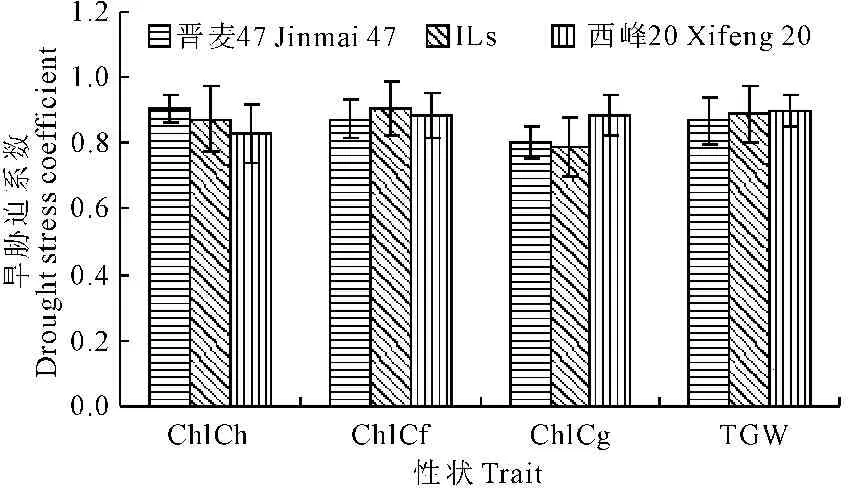
图1 小麦ILs群体与其亲本叶绿素含量和 千粒重的旱胁迫系数
Fig.1 Drought stress coefficients of chlorophyll content of flag leaf and 1000-grain weight of wheat ILs and its parents
2.3 小麦ILs群体旗叶叶绿素和千粒重的通径分析
由表3看出,在不同水分条件下,小麦ILs群体的开花期和灌浆期旗叶ChlC对TGW均有极显著的正向直接作用,且灌浆期正向直接作用较大(0.582**,DS; 0.312**,WW),而拔节期的直接作用较小(-0.074,DS;0.087,WW)。在DS条件下,开花期旗叶ChlC对TGW的间接总效应呈正向作用,而拔节期和灌浆期为负向间接作用;在WW条件下,灌浆期ChlC对TGW的间接总效应呈正向作用,而拔节期和开花期的为负向间接作用。由此可见,小麦不同发育时期旗叶叶绿素对千粒重的作用具有复杂性。对小麦ILs群体不同发育时期旗叶ChlC对TGW的总效应(直接作用与间接作用之和)

表2 不同水分条件下小麦ILs群体各性状之间的相关系数 Table 2 Correlation coefficients among traits of wheat ILs under different water conditions
注:**, 相关性在0.01水平上显著;*相关性在0.05水平上显著。
Note: **, correlation is significant at the 0.01 level; *, correlation is significant at the 0.05 level.
进行综合分析发现,DS条件下总效应分别为-0.231、0.464和0.429;WW条件下分别为-0.137、0.130和0.425。说明在不同水分条件下开花期和灌浆期旗叶叶绿素对千粒重有促进作用,且在干旱胁迫下的作用更为显著。
2.4 小麦ILs群体旗叶叶绿素和千粒重遗传力及表型多样性分析
由表4可以看出,在不同水分条件下,小麦ILs群体在不同发育时期ChlC遗传力在0.29~0.62(DS)和0.20~0.43(WW)之间,表型多样性指数在0.74~0.78(DS)和0.71~0.76(WW)之间,相比较开花期的ChlC具有较高的遗传力和表型多样性指数。TGW的遗传力为0.52(DS)和0.50(WW),表型多样性指数为0.83(DS)和0.82(WW)。各目标性状整体表现出表型多样性指数较高,遗传力低的数量性状的特点,且DS条件下的各性状遗传力和表型多样性指数高于WW条件下的,表明干旱胁迫在一定程度上丰富了各目标性状表型多样性。

表3 不同水分条件下小麦ILs群体不同发育时期叶绿素含量对千粒重的通径分析 Table 3 Path analysis of flag leaf chlorophyll contents at different growth stages to thousand-grain weight of wheat ILs under different water conditions

表4 不同水分条件下小麦ILs群体各性状的 遗传力和多样性指数 Table 4 Heritability and diversity index of traits in wheat ILs under different water conditions
3 讨 论
小麦持绿性和千粒重是小麦重要的生理和农艺性状,是产量形成的重要影响因子[13,19-20]。小麦功能叶片持绿性,即较高的叶绿素含量与光合速率与强度,干物质的积累与转运,籽粒灌浆和产量的形成密切相关[4-6,21-22]。然而,小麦持绿性和千粒重对水分环境响应极为敏感,干旱胁迫可以加速叶绿素降解,促进叶片衰老,削弱光合作用能力,影响籽粒灌浆,从而导致千粒重和产量显著降低,并且具有显著基因型差异和基因型×水分环境互作差异[9-11]。本研究发现,在干旱胁迫条件下,小麦ILs群体及其亲本旗叶叶绿素含量和千粒重较正常灌溉显著下降,但供体亲本西峰20不同发育阶段旗叶叶绿素含量显著高于轮回亲本晋麦47;群体表型介于双亲之间,更偏向轮回亲本晋麦47,且各目标性状表型旱胁迫系数有显著差异,说明小麦ILs群体所考察的目标性状表型对水分环境敏感,存在显著的基因型和发育阶段差异。同时发现,小麦ILs群体不同发育时期旗叶叶绿素含量与千粒重间呈现极显著正相关,其中灌浆期旗叶叶绿素含量与千粒重表现出较高的相关性(r=0.629**,DS;0.622**,WW),且干旱胁迫条件下的相关系数普遍高于灌溉的。通径分析进一步表明,小麦ILs群体旗叶叶绿素对千粒重均具有显著的正向直接作用,且灌浆期正向直接作用较大(0.582**,DS;0.312**,WW),干旱胁迫条件下的总效应大于灌溉的。说明在干旱胁迫条件下,小麦灌浆期维持较高的叶绿素含量对千粒重贡献尤为重要。从而在小麦群体水平上进一步证实了小麦不同基因型品种间比较得出的,在干旱胁迫条件下保持小麦生育后期旗叶较高的叶绿素含量和较多绿叶面积有利于延长光合作用时间和干物质的积累转运,促进籽粒灌浆,增加粒重和产量[6,10,23]的观点。
小麦持绿性和千粒重是典型的微效多基因控制的复杂数量性状[9,12-13,20-21]。前人利用初级作图群体DH群体[20]和RIL群体[9,12,21]对小麦持绿性和千粒重遗传剖析发现,控制这些性状的QTL在小麦整个基因组上均有分布,其遗传受加性、上位性、加性与水分环境互作和上位性与水分环境互作效应控制,不同的水分环境和发育阶段所检测到的QTL的数目、定位区间,以及各遗传因子的效应显著不同。说明控制这些数量性状的基因可能以一定的时空方式表达,在不同环境和不同的发育阶段下具有不同的表达模式[9,12,24-25]。因此,加速开展不同水分环境、不同遗传背景小麦材料后代群体的持绿性和千粒重数量遗传研究,有利于发掘该性状更为丰富的抗旱遗传信息。本研究利用1套ILs群体对不同水分条件下该群体不同发育时期的持绿性和千粒重表型变异发现,各目标性状的表型值介于双亲之间,群体内株系表型变异广泛,存在超亲分离现象,多样性指数高,遗传力低,易受水分环境的影响。表明该群体双亲对所考察性状有贡献的等位基因在其后代群体中得到广泛分离,呈现出多基因控制的数量性状特点,且目标性状表型多偏向于轮回亲本晋麦47,体现出导入系群体的遗传特性。这些研究结果将为进一步解析干旱调控小麦持绿性和千粒重表型变异的遗传基础,及其QTL精细定位奠定良好的材料和理论基础,对小麦抗旱遗传改良具有重要意义。
致谢:感谢中国农科院作物科学研究所景蕊莲研究员为本试验提供实验材料。
[1] 邢素丽,刘孟朝,彭青伟.我国小麦资源与综合生产能力研究[J].干旱地区农业研究,2006,24(2):169-172.
[2] 张 娟,谢惠民,张正斌,等.小麦抗旱节水生理遗传育种研究进展[J].干旱地区农业研究,2005.23(3):231-238.
[3] Biswal AK, Kohli A. Cereal flag leaf adaptations for grain yield under drought: knowledge status and gaps[J]. Molecular Breeding, 2013,31(4):749-766.
[4] Zhang K, Zhang Y, Chen G, et al. Genetic analysis of grain yield and leaf chlorophyll content in common wheat[J]. Cereal Research Communication, 2009,37(4):499-511.
[5] Borrell A K, Hammer G L, van Oosterom E. Stay-green: a consequence of balance between supply and demand for nitrogen during grain filling?[J]. Annals of Applied Biology, 2001,138:91-95.
[6] 李永攀,罗培高,任正隆.小麦持绿性及其与产量关系研究[J].西南农业学报,2008,21(5):1221-1225.
[7] Rampino P, Spano G, Pataleo S, et al. Molecular analysis of a durum wheat ‘stay green’ mutant: Expression pattern of photosynthesis-related genes[J]. Journal of Cereal Science, 2006,43(2):160-168.
[8] Tanaka A, Tanaka R. Chlorophyll metabolism[J]. Current Opinion in Plant Biology, 2006,9(3):248-255.
[9] Yang D L, Li M F, Liu Y, et al. Identification of quantitative trait loci and water environmental interactions for developmental behaviors of leaf greenness in wheat[J]. Frontiers in Plant Science, 2016,7(7):273.
[10] Hafsi M, Hadji A, Guendouz A, et al. Relationship between flag leaf senescence and grain yield in durum wheat grown under drought conditions[J]. Journal of Agronomy, 2013,12(2):67-77.
[11] Naruoka Y, Sherman J D, Lanning S P, et al. Genetic analysis of green leaf duration in spring wheat[J]. Crop Science, 2012,52(1):99-109.
[12] 胡亮亮,叶亚琼,吕婷婷,等.不同水分环境下小麦粒重QTL定位及遗传分析[J].草业学报,2015,24(8):118-129.
[13] Yang D L, Jing R L, Chang X P, et al. Quantitative trait loci mapping for chlorophyll fluorescence and associated traits in wheat(TriticumaestivumL.)[J]. Journal of Integrative Plant Biology, 2007,49(5):646-654.
[14] 施 伟,昌小平,景蕊莲.不同水分条件下小麦生理性状与产量的灰色关联度分析[J].麦类作物学报,2012,32(4):653-659.
[15] 陈稳良,景蕊莲,刘惠民,等.晋麦47背景回交导入系的遗传选择与性状分析[J].麦类作物学报,2009,29(2):206-211.
[16] Ibrahim S E, Schubert A, Pillen K, et al. Comparison of QTLs for drought tolerance traits between two advanced backcross populations of spring wheat[J]. International Journal of Molecular Sciences, 2012,2(3):237-243.
[17] Wu X S, Wang Z H, Jing R L, et al. Genetic dissection of the developmental behaviours of plant height in wheat under diverse water regimes[J]. Journal of Experimental Botany, 2010,6l(11):2923-2937.
[18] Shannon C E, Weaver W. The Mathematical Theory of Communication[M]. Urbana, Chicago, USA: The University of lllinois, 1949:3-24.
[19] Xue Q, Zhu Z, Musick J T, et al. Physiological mechanisms contributing to the increased water-use efficiency in winter wheat under deficit irrigation[J]. Journal of Plant Physiology, 2006,163(2):154-164.
[20] Yang D L, Jing R L, Chang X P, et al. Identification of quantitative trait loci and environmental interactions for accumulation and remobilization of water-soluble carbohydrates in wheat (TriticumaestivumL.) stems[J]. Genetics, 2007,176:571-584.
[21] Verma V, Foulkes M J, Worland A J, et al. Mapping quantitative trait loci for leaf senescene as a yield determinant in winter wheat under optical and drought-stressed environments[J]. Euphytica, 2004,135:255-263.
[22] 侯贤清,贾志宽,韩清芳,等.轮耕对宁南旱区冬小麦花后旗叶光合性能及产量的影响[J].中国农业科学,2011,44(15):3108-3117.
[23] Gong Y H, Zhang J, Gao J F, et al. Slow export of photoassimilate from stay-green leaves during late grain-filling stage in hybrid winter wheat (TriticumaestivumL.)[J]. Journal of Agronomy & Crop Science, 2005,191(4):292-299.
[24] Sun X Y, Wu K, ZhaoY, et al. QTL analysis of kernel shape and weight using recombinant inbred lines in wheat[J]. Euphytica, 2009,165:615-624.
[25] Wang R X, Hai L, Zhang X Y, et al. QTL mapping for grain-fillling rate and yield-related traits in RILs of the Chinese winter wheat population Heshangmai×Yu8679[J]. Theoretical and Applied Genetics, 2009,118:313-325.
猜你喜欢
作物学报(2022年9期)2022-07-18
西南农业学报(2022年5期)2022-06-06
干旱地区农业研究(2021年6期)2021-12-08
河北果树(2021年4期)2021-12-02
今日农业(2021年17期)2021-11-26
中国糖料(2021年3期)2021-07-13
天津医科大学学报(2021年1期)2021-01-26
医药前沿(2020年20期)2020-11-10
浙江农业科学(2019年11期)2019-11-09
河北农业科学(2019年6期)2019-03-21
