
紫茎泽兰对小葱生长的影响及其经济阈值
2018-03-01 08:52曹坳程颜冬冬朱文达李林欧阳灿彬张国云
山西农业大学学报(自然科学版) 2018年2期
曹坳程,颜冬冬,朱文达,李林,欧阳灿彬,张国云
(1.中国农业科学院 植物保护研究所,北京 100193; 2.湖北省农业科学院 植保土肥所,湖北 武汉 430064; 3.德宏州农业环境保护监测工作站, 云南 芒市 678400)
紫茎泽兰(EupatoriumadenophorumSpreng)属于菊科、泽兰属的多年生草本植物,原产于南美洲,作为观赏性植物被引种到欧洲,之后又被引种到澳洲和亚洲。该草远距离扩散主要通过人为引种和交通运输方式,近距离传播主要为种子的随风传播[1]。目前除原产地外,紫茎泽兰已广泛分布在全球热带和亚热带的30多个国家和地区[2]。紫茎泽兰约于20世纪40年代由中缅和中越边境传入我国与其接壤的云南省临沧地区,之后迅速蔓延[3],现已在西南地区云南、四川、贵州、广西等地广泛分布,并逐步随风向东向北传播蔓延[4,5]。由于其发达的根系和巨大的种子库,一旦定植很难将其彻底清除。紫茎泽兰可以入侵农田、草地、经济林地、森林。入侵棉田、花生田,影响棉花和花生的生长发育,显著降低作物的产量[6,7]。紫茎泽兰入侵后,很快会发展成为优势种,甚至会成为单一优势种群,并分泌化感物质,抑制周围其他植物的生长[8]。紫茎泽兰的适应性极强,在年平均温度高于10 ℃,相对湿度不低于68%的气候环境下均适宜生长[9]。随着全球气候变化和温度的上升,我国长江流域及以北区域,如湖北、陕西、河南等省区也可能是其潜在的适生区域[10,11]。小葱(Alliumascalonicum)为百合科葱属草本植物,是一种广泛使用的调味品,南方多产,长江流域多地均有种植。紫茎泽兰不断向北扩散,势必会蔓延到长江流域的小葱产区,影响小葱的生长。本研究通过田间试验采用系列添加的方法,观察了不同密度紫茎泽兰对小葱生长和产量的影响,分析了紫茎泽兰在小葱田对光照和水肥的竞争和消耗,通过模型拟合出不同密度紫茎泽兰与小葱产量损失率之间的数学关系,计算小葱田的经济危害允许水平和紫茎泽兰的经济阈值,为紫茎泽兰的精准防控提供数据支撑。
1 材料与方法
1.1 材料
小葱,又名四季葱,葱属百合科,多年生草本植物,是一种常用调料,南方多产,湖北各地均有种植,选用的小葱为当地市售的小葱品种。
1.2 试验地情况及试验设置
试验点设置在湖北省农业科学院植保土肥所日光温室内。土壤类型为粘壤土,pH7.2、有机质含量2.7%。小葱育苗后移栽,株行距10 cm×15 cm, 每兜3~5株,所有处理小区均采用常规的栽培管理措施。试验分别设置8个不同密度的紫茎泽兰处理,按照0、1、2、5、10、20、40、60株·m-2的密度,试验开始时在各小区人工接种紫茎泽兰的幼苗,每处理设置4次重复,小区之间采用随机区组排列,整个生育期内人工拔除其它干扰杂草。
1.3 调查取样
在小葱生长期(8月23日)调查所有处理小区小葱的长势,在每个小区选取4个样点,每点调查10株小葱的株高、分蘖数、分株数和紫茎泽兰株高。在小葱生长期(9月12日),天气晴朗时采用数字式照度计,选取田间光照强度变化较小的时段(10:00-12:00)分别测量距离地面100 cm、小葱顶部和距离地面15 cm处的光照强度(LIH),每样点重复测量4次,取平均值,计算每个采样点的透光率(LPR):LPR/%=LIH×100/LIT。式中LIH为小葱顶部和距离地面15 cm高度光照强度;LIT为植株顶部光照强度。在小葱采收期,将所有小区的小葱全部收获,测定小区产量。小葱采收后收集每个处理小区紫茎泽兰样品,分别测定紫茎泽兰叶片和茎干样品中全氮、全磷、全钾和水分含量。采用凯氏定氮法测定样品全氮含量,通过干灰化法处理后采用电感耦合等离子光谱仪测定样品的全磷和全钾含量,采用烘干法测定样品中水分含量[12]。
1.4 统计与分析
所有数据均为4次重复的平均值,多重比较和数据拟合均采用SPSS 软件进行数据分析。显著性检验采用重复测量单因素方差分析(ANOVA),不同处理间显著性分析采用邓肯氏新复极差法,采用Pearson相关系数法进行相关性分析。对小葱产量损失率和紫茎泽兰不同密度之间进行拟合分析,拟合模型采用直线(y=ax+b)、指数(y=beax)、二次曲线(y=ax2+bx+c)、幂函数(y=bxa)、对数(y=alnx+b)5种拟合方式根据拟合度和显著性水平筛选出最优的拟合方程。依据小葱产量损失率与紫茎泽兰不同密度的最优模型及小葱生产所允许的产量损失水平确定经济危害允许水平以及紫茎泽兰的经济阈值。经济危害允许水平(EIL)的影响因素包括小葱产量、价格及杂草防除费用,计算方法为:EIL/%=C/(Y×P×E),式中C代表紫茎泽兰防除成本(药剂成本和用工),Y代表小葱的预期产量,P代表小葱市场价格,E为紫茎泽兰的防除效果。
2 结果与分析
2.1 不同密度紫茎泽兰对小葱生长和产量的影响
在小葱生长期,随着紫茎泽兰密度的增加,小葱每兜分蘖数和分株数逐渐减少。当密度为60株·m-2时,小葱每兜分蘖数和分株数比无紫茎泽兰对照分别减少了93.8%和84.9%。当密度为0~60 株·m-2时,小葱株高随着紫茎泽兰密度的增加而降低,小葱的鲜重产量也随紫茎泽兰密度的增加逐渐降低。密度达到60株·m-2时,小葱产量仅为每兜5.3 g,产量损失率达到93.3%(表1)。通过相关性分析发现,紫茎泽兰密度与小葱的株高、分蘖数、分株数、鲜重产量均呈显著负相关(表2)。
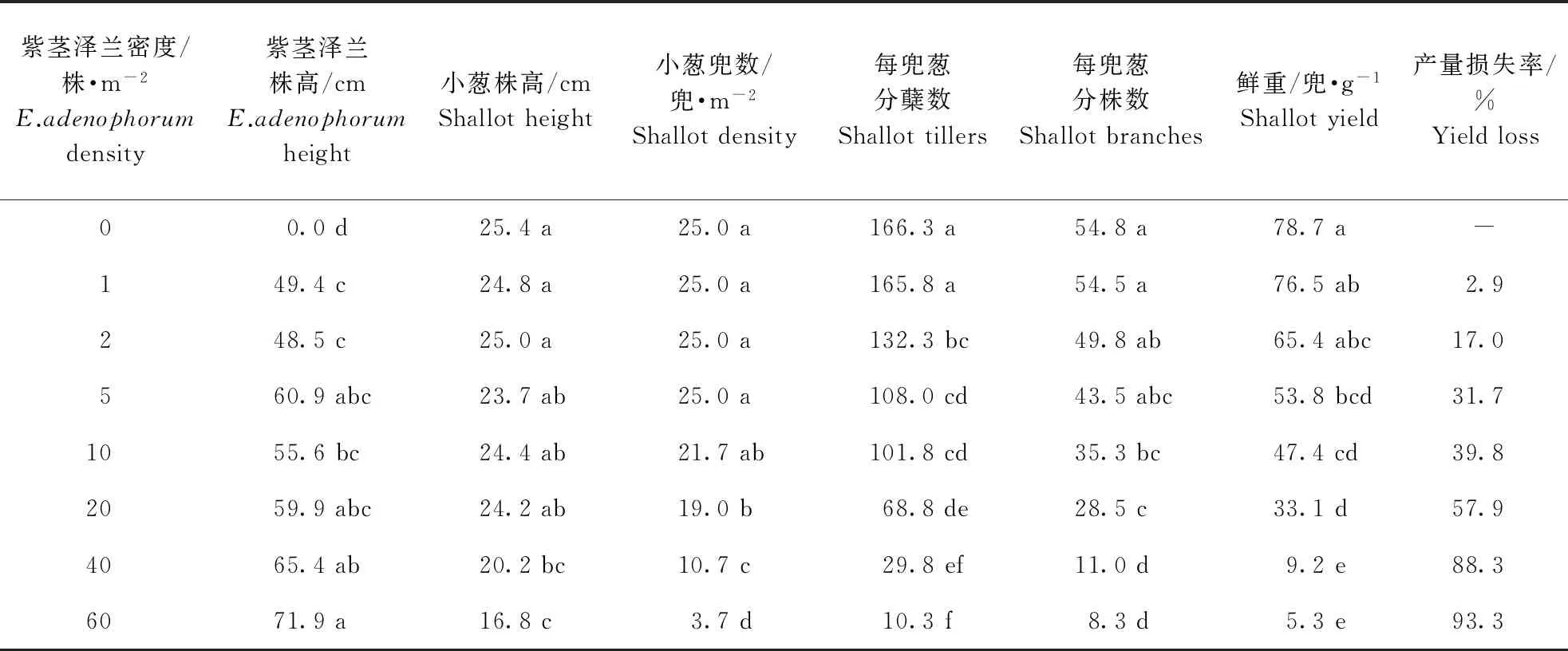
表1 紫茎泽兰对小葱生长及产量的影响Table 1 Effect of E.adenophorum density on shallot growth and yield
注:同列数据后相同字母代表5%水平差异不显著。下同。
Note: Data within a column followed by the same letter are not significantly different atP=0.05 level.The same below.

表2 紫茎泽兰密度与小葱植株经济性状、产量的相关性分析Table 2 Correlation analysis among E.adenophorum density, shallot economic characteristics and yield
注:表中*和**分别代表相关性分析在5%和1%水平差异显著。
Note: Data in the table followed by*and**stand for correlation analysis are significantly different at levels ofP=0.05和P=0.01, respectively.
2.2 紫茎泽兰对水肥的积累和田间光照强度的影响
紫茎泽兰对土壤中养分具有很强的吸收能力,能够极大消耗土壤肥力。随着紫茎泽兰密度的增加,其叶片或茎秆对氮、磷、钾以及水分的累积量均显著增加(表3)。紫茎泽兰对田间的透光率也有显著影响。田间的透光率随着紫茎泽兰密度的增加逐渐降低,当紫茎泽兰发生密度达到40、60 株·m-2时,小葱顶部的透光率仅为6.9%和1.8%(表4)。

表3 不同密度紫茎泽兰植株地上部分氮磷钾及水分含量Table 3 N, P, K and water content of E.adenophorum
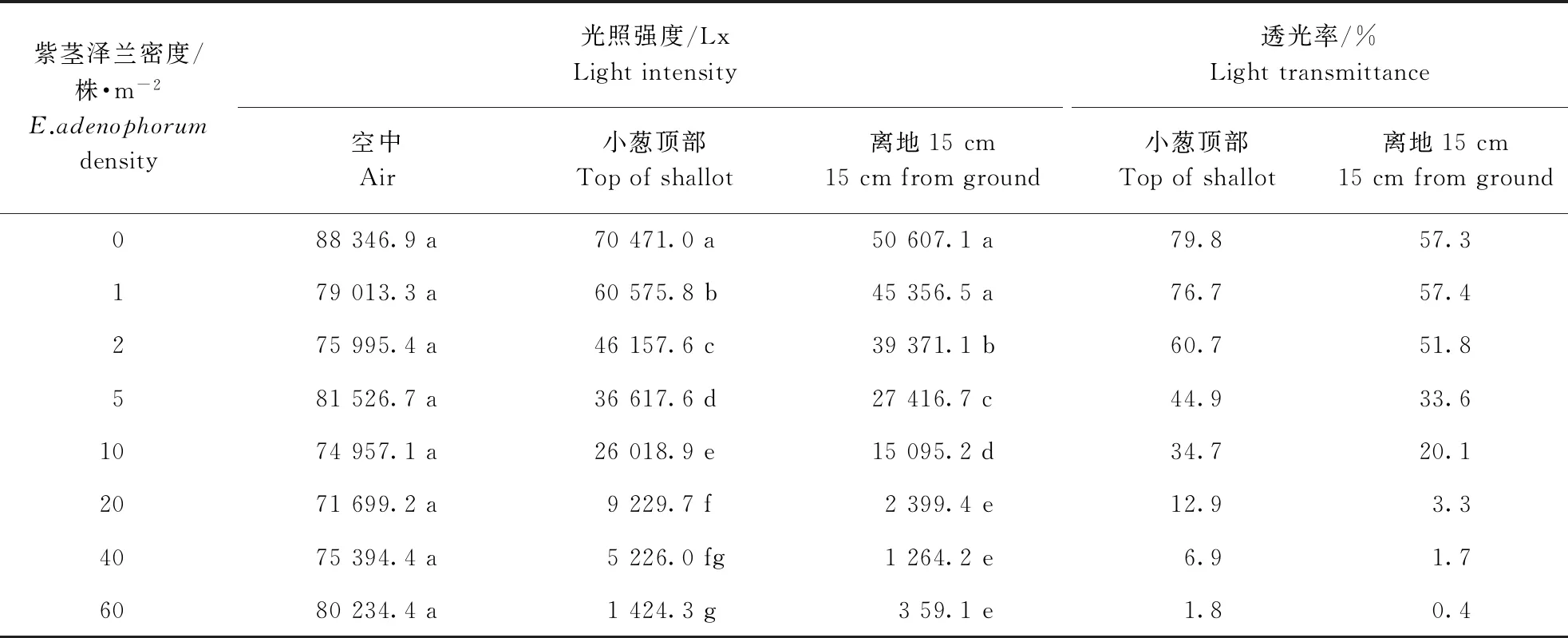
表4 紫茎泽兰处理对小葱田光照强度的影响Table 4 Effect of different density of E.adenophorum on field light intensity
2.3 小葱产量损失率与紫茎泽兰密度的关系
通过曲线拟合对小葱产量损失率和紫茎泽兰密度的关系进行分析。结果表明,幂函数模型拟合能够较好地反应小葱产量损失率和紫茎泽兰密度之间的关系(图1)。曲线拟合的决定系数R2值幂函数最大为0.981,指数函数R2值最小为0.227(表5)。综合分析曲线拟合的决定系数、显著性结果以及曲线与实测值之间的拟合度,发现幂函数曲线模型可以较好地反应小葱产量损失率和紫茎泽兰密度之间的关系,根据拟合的结果选择幂函数模型y=2.823x1.069计算紫茎泽兰的经济阈值。
2.4 紫茎泽兰对小葱经济危害允许水平及经济阈值
小葱的预期产量约为15 000 kg·hm-2,湖北地区小葱的平均价格约为5 元·kg-1。目前田间杂草的防除主要通过人工除草和药剂防控,人工除草一般需要3次,费用约为2 000元·hm-2。采用除草剂进行防除时除药剂费用外还需要施药人工费用。采用41%草甘膦异丙胺盐水剂的防治费用合计为225 元·hm-2。显然,不同除草措施的作物经济危害允许存在一定差异,采用人工除草和41%草甘膦异丙胺盐水剂防除紫茎泽兰时,小葱田的经济危害允许水平分别为3.33%、0.32%。根据经济危害允许水平,由拟合的小葱产量损失率与紫茎泽兰密度的幂函数模型y=2.823x1.069计算出的紫茎泽兰密度即为相应除草措施的经济阈值。人工除草的经济阈值为1.17株·m-2,即当紫茎泽兰发生密度达到1.17 株·m-2以上时,就必须进行人工除草的方法来控制紫茎泽兰的发生;而41%草甘膦异丙胺盐水剂的经济阈值较低,仅为0.13 株·m-2(表6)。
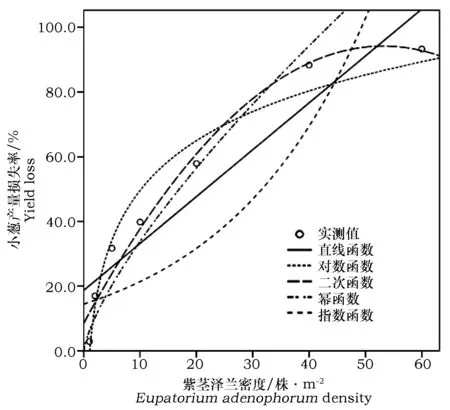
图1 小葱产量损失率与紫茎泽兰密度的曲线拟合Fig.1 Curve fit between yield loss of shallot and E.adenophorum density
Table5 Regression analysis between yield loss of shallot andE.adenophorumdensity
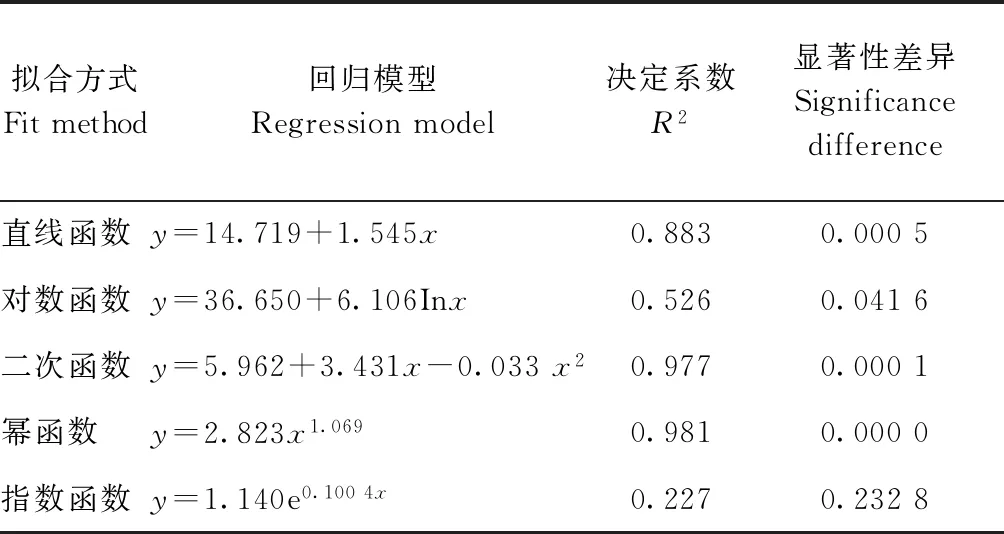
拟合方式Fitmethod回归模型Regressionmodel决定系数R2显著性差异Significancedifference直线函数y=14.719+1.545x0.8830.0005对数函数y=36.650+6.106Inx0.5260.0416二次函数y=5.962+3.431x-0.033x20.9770.0001幂函数y=2.823x1.0690.9810.0000指数函数y=1.140e0.1004x0.2270.2328
表6小葱田不同除草措施的经济危害允许水平和经济阈值
Table6 Economic injury level and economic threshold of weed control measures in shallot fields
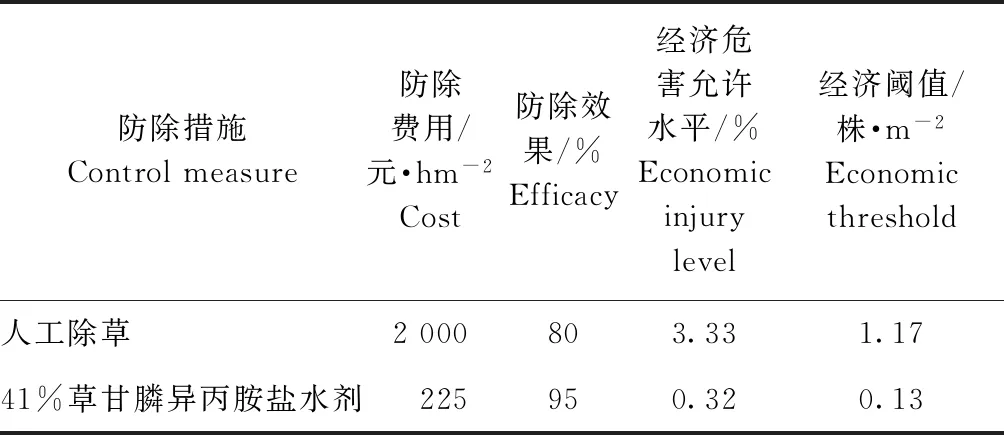
防除措施Controlmeasure防除费用/元·hm-2Cost防除效果/%Efficacy经济危害允许水平/%Economicinjurylevel经济阈值/株·m-2Economicthreshold人工除草2000803.331.1741%草甘膦异丙胺盐水剂225950.320.13
3 结论与讨论
紫茎泽兰入侵农田和林地与农作物和林木竞争水肥、阳光和空间,致使作物和林木的减产,造成严重的经济损失[13]。紫茎泽兰一旦入侵,便很快成为群落中的优势种,甚至发展成单一优势种,并对本地生态系统造成一定危害,导致生物多样性丧失[14]。有研究发现空气湿度和温度是影响紫茎泽兰发生的2个主要生态环境因子,在年平均温度大于10 ℃,相对湿度不低于68%,绝对最低温度-11.5 ℃以上,最高气温不超过35 ℃,最冷月平均温度6 ℃以上的气候环境条件下均能生长[9],随着全球气温的上升,我国长江流域及以北的地区,如湖北、山西、陕西、河南都是紫茎泽兰的潜在入侵区域[11]。紫茎泽兰新枝的萌发是从每年5月开始,至9月为生长旺期,其中高温高湿的7、8月生长最快[15,16],在此条件下紫茎泽兰分枝多、株型高大,较强的水肥光利用效率[17],如入侵农田生态系统,将会对农作物的生长带来一定影响。本研究通过田间试验采用系列添加的方法,分析了不同密度紫茎泽兰对小葱生长和产量的影响。结果表明小葱分蘖数随着紫茎泽兰密度的增加呈减少趋势。当田间紫茎泽兰密度较大时,紫茎泽兰生长繁茂,巨大生物量使得小葱中下部位无法接受阳光照射,导致小葱生长空间形成郁闭环境,不能正常进行光合作用,阻碍小葱的生长。小葱的分蘖数直接影响着产量的高低,紫茎泽兰密度的增加显著降低了小葱的分蘖数,是导致小葱产量显著降低的主要原因。有研究表明紫茎泽兰入侵后,土壤中的速效氮磷钾会显著降低,从而使得土壤肥力大幅下降[18],影响土壤生态系统的组成[19~21],导致土壤严重退化,因此紫茎泽兰对田间养分和水分的累积和消耗也可能是导致小葱减产的重要原因。
作物与杂草间的竞争是导致作物产量降低的重要因素,国内外有关杂草与作物竞争的研究已有较多报道,大多采用数学模型来描述杂草对作物造成的危害[22]。本研究结果显示,湖北地区小葱产量损失率与紫茎泽兰密度之间的关系符合幂函数模型y=2.823x1.069。小葱田采用人工除草和41%草甘膦异丙胺盐水剂防除紫茎泽兰经济危害允许水平分别为3.33%、0.32%,经济阈值则分别为1.17、0.13株·m-2。通过化学方法防治紫茎泽兰仍然是主要方法之一,具有效果好、成本低的优点[23,24]。人工借助机械进行防除也是十分有效的防除方法之一。此外生物替代控制的方法近年来也被广泛推广应用,不仅能够有效控制紫茎泽兰的危害,而且能够取得较大的经济和生态效益[25~27]。早期针对大蒜、茄子、花生和棉花田紫茎泽兰的经济阈值研究中,提出了双曲线函数模型和幂函数模型来评价紫茎泽兰对作物的影响[6,7,28]。方焱等在研究中通过对基础数据的整理分析,采用软件建立模型来预测紫茎泽兰对作物造成的经济损失的风险[29]。Cousens[30]在早期研究中构建了作物产量损失和杂草密度的矩形双曲线模型,模型中的主要特征参数都能反映一定实际的生物学意义,且具有较高的准确性和广泛的实用性。不同防控措施的经济阈值除直接与作物产量、产品价格、防除效果和防除成本有关外,还受许多其他因素的影响,如不同地区、环境气候因子、杂草群体的发生规律等,因此在对紫茎泽兰的实际防治中,具体经济阈值的选择还需结合生态学的理论,研究作物与杂草的竞争关系,制定出适合本地区的生态经济阈值,为杂草的科学防控提供理论依据。
[1] Auld BA,Martin PM.The autoecology ofEupatoriumadenophorumSperng in Australia[J].Weed Research,1975,15(1):27-31.
[2] 刘伦辉,谢寿昌,张建华.紫茎泽兰在我国的分布、危害与防除途径的探讨[J].生态学报,1985,5(1):1-6.
[3] Zhu L,Sun O,Sang W,et al.Predicting the spatial distribution of an invasive plant species (Eupatoriumadenophorum) in China[J].Landscape Ecology,2007,22(8): 1143-1154.
[4] 向业勋.紫茎泽兰的分布、危害及防除意见[J].杂草学报,1991 (4):10-11.
[5] Lu Z J,Ma K P.Spread of the exotic croftonweed (Eupatoriumadenophorum) across southwest China along roads and streams[J].Weed Science,2006,54(6): 1068-1072.
[6] 朱文达,颜冬冬,曹坳程,等.紫茎泽兰对花生生长的影响及其经济阈值[J].中国油料作物学报,2012,34(5):518-522.
[7] 曹坳程,朱文达,颜冬冬,等.紫茎泽兰对棉花生长的影响及经济阈值[J].植物保护学报,2013,40(2):171-176.
[8] 万方浩,刘万学,郭建英,等.外来植物紫茎泽兰的入侵机理与控制策略研究进展[J].中国科学: 生命科学,2011,41(1): 13-21.
[9] 强胜.世界性恶性杂草-紫茎泽兰研究的历史及现状[J].武汉植物学研究,1998,16(4):366-372.
[10] Papes M,Peterson A T.紫茎泽兰EupatoriumadenophorumSpreng.在中国入侵分布预测[J].武汉植物学研究,2003,21(2):137-142.
[11] 鲁萍,桑卫国,马克平.外来入侵种紫茎泽兰研究进展与展望[J].植物生态学报,2005,29(6):1029-1037.
[12] 鲍士旦.土壤农化分析(第3版)[M].北京:中国农业出版社,2000:263-270.
[13] 张国良.生物也疯狂—外来物种入侵的危害[J].科学中国人,2002(10):52-53.
[14] 丁晖,徐海根,刘志磊.外来入侵植物紫茎泽兰对植物多样性的影响[J].生态与农村环境学报,2007,23(2):29-32.
[15] 刘伦辉,刘文耀,郑征,等.紫茎泽兰个体生物及生态学特性研究[J].生态学报,1989,9(1):66-70.
[16] 周世敏,欧国腾,李吉松,等.紫茎泽兰生物生态学特性调查[J].杂草科学,2010(2):38-40.
[17] 何澍然,罗文,赵平,等.外来植物与本地植物光合作用特征比较研究[J].西南农业学报,2015,28(6):2479-2483.
[18] 赵国晶,马云萍.云南省紫茎泽兰的分布与危害的调查研究[J].杂草学报,1989(2):37-40.
[19] 梁作盼,李立青, 万方浩,等.土壤微生物对紫茎泽兰生长与竞争的反馈:不同灭菌方法的比较[J].中国生态农业学报,2016,24(9): 1223-1230.
[20] 朱珣之,李 强,李扬苹,等.紫茎泽兰入侵对土壤细菌的群落组成和多样性的影响[J].生物多样性, 2015,23 (5): 665-672.
[21] 邓丹丹,刘棋,蒋智林,等.紫茎泽兰与不同植物群落土壤养分及酶活性差异[J].生态环境学报,2015,24(9): 1466-1471.
[22] 刘德立,Lovett J V,Johnson I R.杂草对作物产贵损失的经验模型[J].植物保护学报,1991,18(4):371-376.
[23] 曹坳程,芮昌辉,郭美霞,等.紫茎泽兰的化学防除及生态修复效果[J].安全与环境学报,2006,6(3):80-85.
[24] 陈才俊,廖国会,秦立新,等.紫茎泽兰化学防除效果的药剂筛选试验[J].贵州农业科学,2009,37(10):101-103.
[25] 朱文达,何燕红,李林.大豆竞争替代紫茎泽兰的种植模式研究[J].中国油料作物学报,2015,37(2):214-219.
[26] 朱文达,杨新笋,胡洪涛,等.甘薯替代控制紫茎泽兰的研究[J].湖北农业科学,2015,54(14):3448-3450,3453.
[27] 李林,曹坳程,喻大昭,等.油菜对紫茎泽兰的替代控制效果[J].中国油料作物学报,2016,38(4):513-517.
[28] 朱文达,何燕红,曹坳程.紫茎泽兰对茄子产量的影响及其经济阈值[J].植物保护学报,2011,38(6):563-568.
[29] 方焱,秦萌,李志红,等.紫茎泽兰对我国花生产业造成的潜在经济损失评估[J].中国农业大学学报,2015,20(6):146-151.
[30] Cousens R.A simple model relating yield loss to weed density[J].Annals of Applied Biology,1985,107(2): 239-252.
猜你喜欢
热带农业工程(2022年3期)2022-08-16
科教新报(2022年22期)2022-07-02
山东畜牧兽医(2020年11期)2020-11-24
中成药(2019年12期)2020-01-04
高中生学习·阅读与写作(2019年2期)2019-09-10
中成药(2018年12期)2018-12-29
中成药(2018年6期)2018-07-11
中国化妆品(2017年12期)2017-06-27
绿色科技(2013年12期)2013-08-28
杂草学报(2012年1期)2012-11-06
