
河南4种栎属树木叶脉序特征研究
2018-01-23 00:42:16陈丹丹张益菻张硕新
西北林学院学报 2018年1期
陈丹丹,郝 凡,张益菻,张 通,张硕新,2*
(1.西北农林科技大学 林学院,陕西 杨陵 712100;2.陕西秦岭森林生态系统 国家野外科学观测研究站,陕西 宁陕 711699)
叶脉在叶片中的分布和排列式样就是叶片脉序,也称为叶脉网络结构。植物在不断适应环境的过程中,其叶脉序特征也对环境条件产生适应性[1],且植物叶脉序特征稳定[2]。叶的宏观结构特征是众多学者进行活植物分类和叶化石鉴定的重要依据[3],近年来叶的很多稳定性状如整体脉型、一级脉、二级脉、三级脉、边缘末级脉、网眼和叶缘齿端结构等被越来越广泛地应用在现代植物分类的研究中[4],叶脉性状在种的鉴定方面体现出重要的意义[5-7]。
栎属(Quercus)20余种落叶栎构成了我国温带和暖温带地区落叶阔叶林和针阔混交林的主要组成部分[8]。栎属植物为适应不同的生存环境,叶脉序特征也产生相应的变异。河南宝天曼自然保护区地处伏牛山南麓,是北亚热带向南暖温带过渡区域,也是中原地区保存最为完好的综合性森林生态系统,植物群落类型复杂,具有丰富的生物多样性[9-10]。短柄枹栎(Q.serratavar.brevipetiolata)、麻栎(Q.acutissima)、锐齿槲栎(Q.alienavar.acuteserrata)和栓皮栎(Q.variabilis)均是河南落叶阔叶林的重要成分。国内对栎属植物叶脉序的研究鲜有报道,因此对河南宝天曼自然保护区内短柄枹栎、麻栎、锐齿槲栎和栓皮栎的叶脉序特征进行提取和分析,探究其叶脉序特征差异,具有一定的理论和实践价值。
1 材料与方法
在宝天曼自然保护区内采摘短柄枹栎、麻栎、锐齿槲栎和栓皮栎的叶片。各树种均选取阳坡正常生长的健康树木8株,每株摘取阳面中部3~5个小枝,每株的小枝上采摘完全展开的成熟叶片共20片,使用彩色扫描仪(BenQ 5560C)逐一对新鲜叶片进行扫描,获取灰度图像。
由于各树种叶片大小不一,叶面积过大会对数据的精确性产生一定的影响,故统一对叶片图像进行截取。以栓皮栎为例,通过分析发现,所获取的8个叶脉序特征指标在完整叶片图像与截取的主脉长度1/3中部图像间均无明显差异,即P>0.05。故截取叶片中部主脉长度的1/3的图像,使用Leaf extraction and analysis framework graphical user interface(简称LEAF GUI)叶脉序分析软件获取叶脉网络结构特征数据。
短柄枹栎、麻栎、锐齿槲栎和栓皮栎叶片图像样本共计115个。8个叶脉序特征分别为叶脉平均宽度、网眼密度(单位面积网眼数)、网眼面积(叶脉所围成的闭合区间面积)、叶脉间距离(闭合环形结构的最大当量直径)、叶脉密度(单位面积叶脉长度)、叶脉边缘平均二维面积(叶脉所围成的闭合区间中叶脉的二维面积)、叶脉边缘平均三维面积(叶脉围成的近似圆柱体的表面积)和闭合环形结构平均体积(叶脉所围成的近似圆柱体的体积)。
通过主成分分析、单因素方差分析和多重比较的方法,对叶脉序特征数据进行分析。所有分析和作图均使用SPSS Statistics V20.0软件和Microsoft Excel 2010完成。
2 结果与分析
2.1 叶脉序特征指标的筛选
对短柄枹栎、麻栎、锐齿槲栎和栓皮栎的叶脉序特征指标进行主成分分析(PCA)(表1)。从各主成分的方差贡献率可以看出,第1主成分和第2主成分的累积贡献率达到87.351%,即第1主成分和第2主成分可以解释原8个变量的87.351%变异,所反映的信息已经比较全面,故提取前2个主成分。
以叶脉序特征主成分为横坐标,各主成分对应的特征值为纵坐标得到叶脉序特征主成分分析提取图(图1)。主成分1和主成分2之间连线的斜率比较大,从第3个主成分开始,其对应的特征值都很小,这对解释原变量的贡献也很小,可以被忽略。图1 与表1的结果相一致,说明提取2个主成分较为合理。
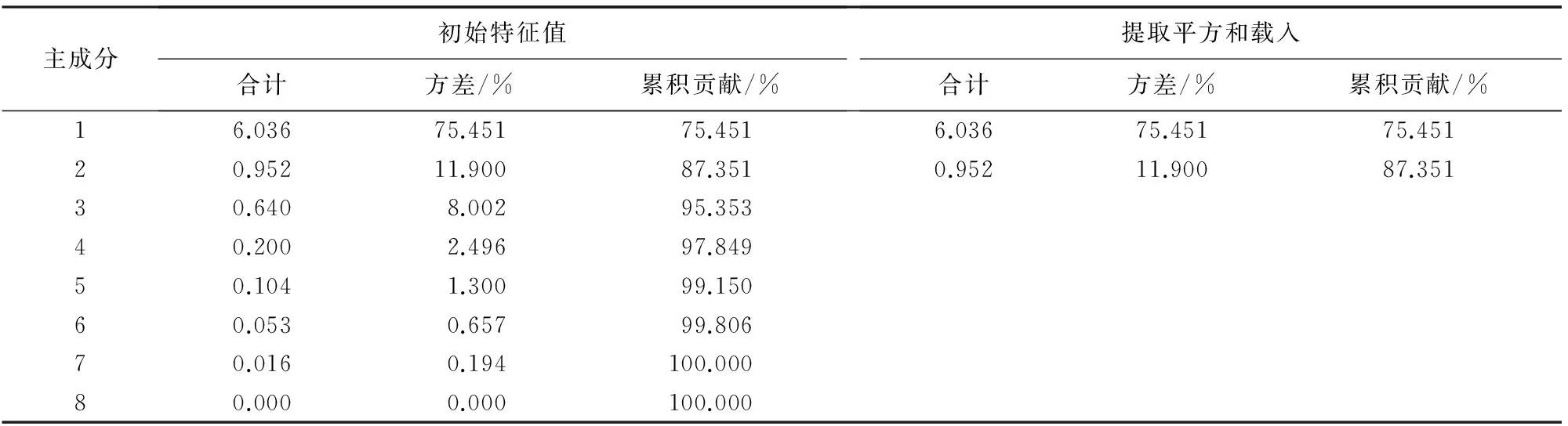
表1 短柄枹栎、麻栎、锐齿槲栎和栓皮栎叶脉序特征主成分的特征值和累积贡献
主成分与各叶脉序特征指标相关程度见表2。第1主成分与叶脉边缘平均三维面积、叶脉边缘平均二维面积、闭合环形结构平均体积和叶脉密度的相关性较高;第2主成分与叶脉间距离相关性较高。
经过对原8个叶脉序特征进行主成分分析,筛选出最适合进行研究的叶脉序特征共5个,分别为叶脉边缘平均三维面积、叶脉边缘平均二维面积、闭合环形结构平均体积、叶脉密度和叶脉间距离。
2.2 叶脉序特征差异比较
短柄枹栎、麻栎、锐齿槲栎和栓皮栎的5个重要叶脉序特征方差分析和多重比较结果见图2。短柄枹栎的叶脉边缘平均三维面积、叶脉边缘平均二维面积、叶脉间距离和叶脉密度与麻栎、锐齿槲栎、栓皮栎均存在显著差异(P<0.05),栓皮栎的这4个特征与麻栎、锐齿槲栎也存在显著差异(P<0.05),麻栎与锐齿槲栎之间的差异不显著(P>0.05)。在闭合环形结构平均体积上,短柄枹栎、麻栎、锐齿槲栎和栓皮栎两两之间均存在显著差异(P<0.05)。

图1 短柄枹栎、麻栎、锐齿槲栎和栓皮栎叶脉序特征的主成分提取

叶脉序特征主成分12叶脉边缘平均三维面积0.973-0.102叶脉边缘平均二维面积0.973-0.102闭合环形结构平均体积0.965-0.128叶脉密度-0.9370.166网眼面积0.8610.410叶脉平均宽度0.856-0.265网眼密度-0.7340.136叶脉间距离0.5680.794
从柱形图中可以看出,短柄枹栎的叶脉边缘平均三维面积、叶脉边缘平均二维面积、闭合环形结构平均体积和叶脉间距离最大,栓皮栎次之,锐齿槲栎与麻栎的叶脉边缘平均三维面积、叶脉边缘平均二维面积相差不多,但锐齿槲栎的闭合环形结构平均体积和叶脉间距离稍>麻栎。叶脉密度最大的是麻栎和锐齿槲栎,两者无明显差别,栓皮栎次之,短柄枹栎最小。
3 结论与讨论
相比短柄枹栎而言,麻栎、锐齿槲栎和栓皮栎可能通过增加叶脉密度而更适应寒冷和干旱的环境条件。叶脉边缘三维面积和二维面积小、叶脉间距离以及闭合环形结构体积小、叶脉密度和网眼密度大的特点可能使麻栎和锐齿槲栎的叶脉系统运输能力相对更强。叶脉密度和网眼密度大,增加了麻栎、锐齿槲栎和栓皮栎叶脉系统的机械支持能力和抵御外部损伤能力。
注:不同小写字母表示不同树种之间在0.05水平存在显著差异。
图2短柄枹栎、麻栎、锐齿槲栎和栓皮栎叶脉序特征比较
Fig.2 Comparisons of five venation characteristics among four oak species
叶脉承担着运输植物养分和水分的重要作用,其结构特征具有种的特异性,同时对环境具有适应性[11]。植物随着环境中生态因子的变化而发生形态结构的变化,其中叶的变化是最明显和快速的[12]。叶脉边缘平均三维面积、叶脉边缘平均二维面积、闭合环形结构平均体积、叶脉间距离和叶脉密度等特征在不同的栎属植物中呈现出差异性,是物种与环境相互作用而产生的结果。
李晓鹏[13]通过研究发现,北方枣品种通过增加叶脉密度而提高自身的抗旱性和抗寒性。在干旱胁迫下,植物通过增加末端叶脉密度来响应水分的变化[14]。落叶栎在中国的地理分布[15]显示,麻栎、锐齿槲栎和栓皮栎的分布北界明显比短柄枹栎分布北界纬度更高,麻栎、锐齿槲栎和栓皮栎的分布西界也明显比短柄枹栎分布西界更靠近四川西部,由此推测麻栎、锐齿槲栎和栓皮栎的抗寒性和抗旱性>短柄枹栎,研究结果显示麻栎、锐齿槲栎和栓皮栎的叶脉密度明显>短柄枹栎。这与李晓鹏[13]的研究结果相似。
李乐[1]等研究发现,不论是末端叶脉还是较粗叶脉,其密度越大,叶脉系统的物质运输能力则越强。不同物种中,高叶脉密度则能支持高的光合速率[16]。栎属树种在南北样带温暖带区内,由北往南,其叶脉密度增加,较高的叶脉密度保证了叶片水分和养分的运输,促进叶片生理活动正常进行[17]。叶脉间距离越小,则叶脉到气孔的距离就短,允许通过的水分和碳通量就越高[18]。叶脉密度和叶脉间距离等性状对水分、养分和光合产物在叶脉系统中的运输产生重要的影响[19]。分析结果显示麻栎和锐齿槲栎的叶脉间距离明显短于栓皮栎和短柄枹栎,麻栎和锐齿槲栎的叶脉密度也相对更高,而麻栎具有养分奢侈吸收、养分损失少和养分再吸收能力强的特性[20],生长迅速,适应能力强[21],35年生锐齿槲栎林地上部分净生产力与年龄呈正相关,年龄递增则叶的分配比递增[22],由此推测麻栎和锐齿槲栎的叶脉系统具有较强的运输能力。
叶脉系统的运输能力与闭合环形结构的特性有着密切的关系,当闭合环形结构的特点有利于水分、养分及光合产物的流动和运输时,则叶脉系统的运输能力更强。相比短柄枹栎和栓皮栎,麻栎和锐齿槲栎的叶脉间距离短、叶脉密度大,其叶脉系统的运输能力更强,说明其闭合环形结构特点有利于物质的流动。研究中发现,麻栎和锐齿槲栎的叶脉边缘平均三维面积和平均二维面积、叶脉间距离以及闭合环形结构平均体积明显<栓皮栎和短柄枹栎,说明麻栎和锐齿槲栎的网眼密度更大,且通过单因素方差分析发现,麻栎和锐齿槲栎的网眼密度明显>短柄枹栎(P<0.05),但栓皮栎的网眼密度与锐齿槲栎处于同一水平。由此推测,叶脉边缘三维面积和二维面积小、叶脉间距离短、闭合环形结构体积小的特点更有利于提高叶脉系统的运输能力。
叶脉密度和网眼密度等性状还与叶脉系统的机械支持能力和抵御虫害等干扰能力有着密切的关系,叶脉密度大更有利于提高机械支持能力,叶脉密度和网眼密度大则更有利于抵御虫害[1]。李东胜[17]等认为栎属植物多为高大乔木,叶片在群落冠层中更加突出,降水和风吹等外部影响更为明显,叶脉密度增加可以增大叶片的强韧度,从而减少或避免外部冲击造成的机械损伤。麻栎、锐齿槲栎和栓皮栎的叶脉密度和网眼密度都明显>短柄枹栎,由此推测麻栎、锐齿槲栎和栓皮栎的叶脉系统机械支持能力和叶片抵御外部损伤能力更强。
[1] 李乐,曾辉,郭大立.叶脉网络功能性状及其生态学意义[J].植物生态学报,2013,37(7):691-698.
LI L,ZENG H,GUO D L.Leaf venation functional traits and their ecological significance [J].Chinese Journal of Plant Ecology,2013,37(7):691-698.(in Chinese)
[2] ROTH-NEBELSICK A,UHL D,MOSBRUGGER V,etal.Evolution and function of leaf venation architecture:a review[J].Annals of Botany,2001,87(5):553-566.
[3] 胡蓉,周春景.4种柑橘属植物叶脉序比较研究[J].内江师范学院学报,2011,26(2):34-37.
[4] 刘利霞,侯元同,李法曾.中国拳参属(蓼科)植物叶脉序式样的研究[J].西北植物学报,2006,26(8):1554-1561.
LIU L X,HOU Y T,LI F Z.Leaf-venation patterns ofBistorta(Polygonaceae) in China [J].Acta Botanica Boreali-Occidentalia Sinica,2006,26 (8):1554-1561.(in Chinese)
[5] 刘颖,孙稚颖,李法曾.十字花科芸薹族植物叶脉序的研究[J].西北植物学报,2006,26(3):544-550.
LIU Y,SUN Z Y,LI F Z.Venation of the Tribe Brassiceae in China [J].Acta Botanica Boreali-Occidentalia Sinica,2006,26 (3):544-550.(in Chinese)
[6] 许炳强,夏念和,王少平,等.中国木犀属植物叶脉形态及其分类学意义[J].广西植物,2007,27(5):697-705.
XU B Q,XIA N H,WANG S P,etal.Leaf venation ofOsmanthus(Oleaceae) from China and its taxonomic significance [J].Guihaia,2007,27(5):697-705.(in Chinese)
[7] ELLIS B.Manual of leaf architecture[M].Ithaca:Cornell University Press,2009:1-5.
[8] 张金香,王海霞,杨鸿飞.栎树利用价值及资源培育[J].河北林业科技,2014(3):76-77.
[9] 史作民,刘世荣,程瑞梅,等.宝天曼落叶阔叶林种间联结性研究[J].林业科学,2001,37(2):29-35.
SHI Z M,LIU S R,CHENG R M,etal.Interspecific association of plant populations in deciduous broad-leaved forest in Baotianman[J].Scientia Silvae Sinicae,2001,37(2):9-35.(in Chinese)
[10] 闫东锋,王亚南,杨喜田,等.宝天曼栎类天然次生林主要树木种群结构及空间分布格局[J] .西北林学院学报,2012,27(3):155-159.
YAN D F,WANG Y N,YANG X T,etal.Structure and spatial distribution patterns of the main woody plant populations of oak natural secondary forests in Baotianman nature reserve,Henan Province[J].Journal of Northwest Forestry University,2012,27(3):155-159.(in Chinese)
[11] 张钟,李潇雅.枣叶脉网络结构特征研究[J].陕西林业科技,2014(4):8-10.
[12] 王金照,张文辉.不同生境下栓皮栎叶形态解剖的研究[J] .西北林学院学报,2004,19(2):44-46.
WANG J Z,ZHANG W H.The research on form dissecting ofQuercusvariabilisleaf in different habitats[J].Journal of Northwest Forestry University,2004,19(2):44-46.(in Chinese)
[13] 李晓鹏.枣叶脉序及其功能性状研究[D].陕西杨陵:西北农林科技大学,2016.
[14] 李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(Supp.1):118-127.
LI F L,BAO W K.Response of the morphological and anatomical structure of the plant leaf to environmental change [J].Chinese Bulletin of Botany,2005,22(Supp.1):118-127.(in Chinese)
[15] 王良民,任宪威,刘一樵.我国落叶栎的地理分布[J].北京林学院学报,1985(2):57-69.
[16] 宋丽清,胡春梅,侯喜林,等.高粱、紫苏叶脉密度与光合特性的关系[J].植物学报,2015,50(1):100-106.
[17] 李东胜,史作民,冯秋红,等.中国东部南北样带暖温带区栎属树种叶片形态性状对气候条件的响应[J].植物生态学报,2013,37(9):793-802.
LI D S,SHI Z M,FENG Q H,etal.Response of leaf morphometric traits ofQuercusspecies to climate in the temperate zone of the north-south transect of eastern China [J].Chinese Journal of Plant Ecology,2013,37(9):793-802.(in Chinese)
[18] BRODRIBB T J,FEILD T S,JORDAN G J.Leaf maximum photosynthetic rate and venation are linked by hydraulics[J].Plant Physiology,2007,144(4):1890-1898.
[19] SACK L,HOLBROOK N M.Leaf hydraulics[J].Annual Review of Plant Biology,2006,57:361-381.
[20] 唐罗忠,刘志龙,虞木奎,等.两种立地条件下麻栎人工林地上部分养分的积累和分配[J].植物生态学报,2010,34(6):661-670.
TANG L Z,LIU Z L,YU M K,etal.Nutrient accumulation and allocation of aboveground parts inQuercusacutissimaplantations under two site conditions in Anhui,China [J].Chinese Journal of Plant Ecology,2010,34(6):661-670.(in Chinese)
[21] 魏国余,覃德文,孙灿岳,等.麻栎人工林生长规律模拟与研究[J].西北林学院学报,2014,29(4):145-150.
WEI G Y,QIN D W,SUN C Y,etal.Simulation and research on the growth regularity ofQuercusacutissimaplantation[J].Journal of Northwest Forestry University,2014,29(4):145-150.(in Chinese)
[22] 刘玉萃,吴明作,郭宗民,等.宝天曼自然保护区栓皮栎林生物量和净生产力研究[J].应用生态学报,1998,9(6):11-16.
LIU Y C,WU M Z,GUO Z M,etal.Biomass and net productivity ofQuercusvariabilisforest in Baotianman natural reserve [J].Chinese Journal of Applied Ecology,1998,9(6):11-16.(in Chinese)
猜你喜欢
广东蚕业(2022年4期)2022-11-23 13:31:55
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
核农学报(2020年6期)2020-06-30 06:15:38
南方农业·下旬(2020年1期)2020-06-01 07:47:32
现代园艺(2019年24期)2019-01-07 01:19:36
现代农业科技(2009年21期)2018-09-03 00:00:00
植物研究(2018年4期)2018-07-24 01:06:24
农村百事通(2017年18期)2017-02-08 11:23:54
云南中医学院学报(2012年4期)2012-07-31 18:06:20
