
麻栎和栓皮栎叶脉序与环境因子关系研究
2018-01-23 00:42:48张益菻陈丹丹张硕新
西北林学院学报 2018年1期
张益菻,陈丹丹,郝 凡,张 通,张硕新,2*
(1.西北农林科技大学 林学院,陕西 杨陵 712100;2.陕西秦岭森林生态系统国家野外科学观测研究站,陕西 宁陕 710600)
环境变化对于森林植物的影响复杂多样,而植物对环境的适应能力和调控能力可以通过叶片性状反映[1]。植物的叶片是植物最主要的生命活动场所,所以用叶片作为研究对象更能反映植物对气候变化的响应和适应机制[2-3]。叶脉网络结构(叶片脉序,leaf venation)是叶脉系统(leaf vein system)重要的形态结构指标,它表征了叶脉系统在叶片里的分布和排列样式[4-5]。不同植物类群具有不同类型的叶脉网络结构,所以叶脉序在植物分类上一直起着重要作用[6]。随着研究的不断深入,叶脉网络功能性状与叶片水分关系、叶片光合能力、叶片碳构建以及其与环境之间的关系的研究得到广泛关注[7-9]。
麻栎(Quercusacutissima)和栓皮栎(Q.variabilis)作为我国重要的森林树种,关于其生理生长相关研究较为常见,但是对其叶脉与环境因子的相关研究鲜见报道。鉴于此,拟对其叶脉结构与环境的相关性进行研究,以便揭示其叶脉结构对环境变化的适应特征。
1 材料与方法
1.1 研究区概况
麻栎叶片采集点为辽宁省、河南省、山东省、江苏省和陕西省;栓皮栎叶片采集点为辽宁省、北京市、河南省、山东省、江苏省、山西省和陕西省。在中国气象科学数据共享服务系统的中国自然资源数据库中采集得到每个采样地的气候数据,包括年平均降水量和年平均温度等(表1)。

表1 采样地位置和气候特点Table 1 Sampling location and climatic characteristics
1.2 样品采集和数据分析
在采样地栎属的中心分布区,选取阳坡林分,在30 m×30 m的样地内,随机选取8棵健康树木(林龄40 a左右),在每棵树上选取阳面中部3~5个小枝,采摘完全伸展的健康叶片20片,放在塑封袋保存。并且登记好采样的时间、样本树的基本情况、采样地的海拔高度和地理位置等,采样时尽量选择环境条件相对一致的区域。
叶脉图像采集:采用背光扫描法采集叶脉信息,将麻栎和栓皮栎的叶片剪掉叶柄后在高分辨率扫描仪下进行背光扫描。扫描出来的叶片图像选取叶脉序清晰的图像5~10张(重复),通过Photo shop软件将图片转换成JEPG格式。用叶脉序信息转换软件Leaf Extraction and Analysis Framework Graphical User Interface(LEAF GUI)进行叶脉序特征分析(图1)。通过设定比例尺、图像剪裁、图像阈值化,调整图像的全局阈值、自适应阈值,获取全叶图像。再通过二进制图像清理和数据运算分析得到面积统计、网眼统计和静脉统计等相关数据,并选取14个叶脉序特征:叶面积、叶周长、环形结构(单位面积内网眼数)、叶脉宽度(叶脉厚度大小)、网眼数、网眼面积(叶脉所围成的闭合区间面积)、节点数、叶脉间距离(每个闭合环形结构的最大当量直径的平均值)、叶脉密度(单位面积内叶脉总长度)、叶脉总长度、叶脉总面积、叶脉边缘的平均二维面积(叶脉所围成的闭合区间中叶脉的二维面积)、叶脉边缘的平均三维面积(叶脉围成的近似圆柱体的表面积)和闭合环状结构平均体积(叶脉所围成的近似圆柱体的体积)。
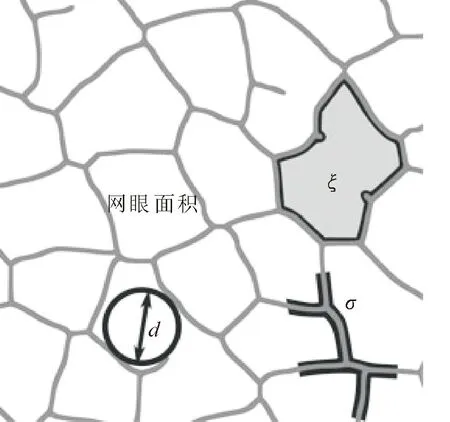
注:叶脉密度(σ);叶脉间距离(d);环形结构(ξ)[4]。
对14个叶脉序指标使用SPSS19.0软件(Statistical Product and Service Solutions)进行主成分分析,得到解释的总方差(表2)。从各主成分的方差贡献率可以看出,第1主成分特征值为7.603,它所解释的方差占总方差的54.31%,第2主成分所解释的方差占总方差的22.931%,第3主成分所解释的方差占总方差的15.444%。通过各主成分的方差累计贡献率可以看出,前3个主成分可以解释原14个变量的92.685%变异,已经能比较全面得反映所有信息。
其中,第1主成分与叶周长、叶脉宽度和叶脉密度相关性较高;第2主成分与网眼数、网眼面积、环形结构和叶脉间距离相关性较高;第3主成分与节点数、叶脉总长度和叶面积相关性较高(表3)。因此对麻栎和栓皮栎的14个叶脉序特征因子经过主成分提取,用SPSS19.0软件对这10个叶脉序指标分别与气象因子进行相关性分析。得到与气象因子相关的8个指标包括叶面积、叶周长、网眼面积、网眼数、环形结构、叶脉间距离、节点数和叶脉密度。在Origin8.5软件中对这8个特征指标与环境因子进行相关性分析作图。

表2 解释的总方差Table 2 Total variance explained

表3 麻栎和栓皮栎叶脉序特征的主成分分析Table 3 Principal components analysis based on leaf venation characteristic data of Q.acutissima and Q.variabilis
2 结果与分析
2.1 麻栎和栓皮栎叶脉序与年平均温度的关系
2.1.1 叶片形态指标与年平均温度的关系 叶片大小在不同树种和不同生境之间差异很大,它影响着植物对光的截取和对碳的获取能力,在自然条件下表现出较大的可塑性。本试验发现麻栎和栓皮栎叶片形态与气候因子之间有较强的相关性。其中,麻栎叶面积(r2=0.882 3,P=0.009)、叶周长(r2=0.778 9,P=0.031)与年平均温度呈正相关;栓皮栎叶面积(r2=0.739 67,P=0.011)、叶周长(r2=0.733,P=0.014)与年平均温度呈正相关。麻栎的叶面积随着年平均温度的变化比栓皮栎更明显,而栓皮栎的叶周长随着年平均温度的变化比麻栎更敏感(图2)。
2.1.2 叶脉结构与年平均温度的关系 麻栎和栓皮栎功能性状特征是在长期演化过程中适应环境的结果,它强调与生态系统的关系,表现为植物在自然选择压力下的生态策略。[10]有研究发现,叶片的变化与气候因素密切相关[11]。本试验通过对麻栎和栓皮栎的叶脉结构与年均温度的相关性分析发现:麻栎(r2=0.897 91,P=0.019)和栓皮栎(r2=0.598 66,P=0.032)的叶脉间距离都与年均温度呈正相关性,并且二者的叶脉间距离对年均温度变化的敏感性相近(图3)。

注:误差线即标准差,下同。

图3 麻栎和栓皮栎叶脉间距离与年平均温度的关系Fig.3 Relationships between Q.acutissima and Q.variabilis leaf traits of distance between veins and the mean annual temperature
2.2 麻栎和栓皮栎叶脉序与年平均降水量的关系
有研究发现植物的叶片大小与降水量直接相关,与温度的相关性次之[12]。本试验发现麻栎和栓皮栎的部分叶脉特征指标也与年平均降水量密切相关:麻栎(r2=0.847 69,P=0.02)和栓皮栎(r2=0.679 21,P=0.045)的网眼数与年平均降水量呈负相关,且栓皮栎的网眼数随着年平均降水量变化的更为明显(图4);麻栎和栓皮栎的节点数与年平均降水量呈负相关(图5);麻栎和栓皮栎的叶脉密度也与年平均降水量呈负相关,且麻栎的叶脉密度随着年平均降水量变化更为敏感(图6);栓皮栎的网眼面积与年平均降水量呈正相关,单位面积网眼数却与年平均降水量呈负相关(图7)。

图4 麻栎和栓皮栎网眼数与年平均降水量间的关系Fig.4 Relationships between Q.acutissima and Q.variabilis leaf traits of areole number and the mean annual precipitation
3 结论与讨论
本研究结果表明麻栎和栓皮栎叶脉结构随着气候变化的总体趋势相同。麻栎叶面积、叶周长和叶脉间距离与年平均温度呈正相关,网眼数、节点数和叶脉密度与年平均降水量呈负相关;栓皮栎叶面积、叶周长和叶脉间距离与年平均温度呈正相关性,网眼面积与年平均降水量呈正相关性,网眼数、环形结构、节点数和叶脉密度与年平均降水量呈负相关性。麻栎的叶面积随着年平均温度的变化比栓皮栎更明显,而栓皮栎的叶周长随着年平均温度的变化比麻栎更敏感。二者的叶脉结构随着生存环境的变化展现出较高的生态适应性。

图5 麻栎和栓皮栎节点数与年平均降水量间的关系Fig.5 Relationships between Q.acutissima and Q.variabilis leaf traits of number of nodes and the mean annual precipitation

图6 麻栎和栓皮栎叶脉密度与年平均降水量间的关系Fig.6 Relationships between Q.acutissima and Q.variabilis leaf traits of vein length per area and the mean annual precipitation

图7 栓皮栎网眼面积和单位面积网眼数与年平均降水量间的关系Fig.7 Relationships between Q.variabilis leaf traits of areole area and number of areoles per leaf area and the mean annual precipitation
麻栎和栓皮栎的叶片大小即叶面积和叶周长都随着年平均温度的升高而增大,这与T.Navarro[19]以及D.J.Peppe[12]等进行的许多大尺度的研究结果相一致。这可能是因为在低温条件下小叶片的蒸腾成本更低[13],处于这种低代谢状态下有利于树木各器官寿命的延长[14],并且在温度较低的条件下植物不能满足更大叶片的养分和传输需求,所以随着温度的降低麻栎和栓皮栎的叶片大小也随之减小。一些控制试验研究也表明增加温度可以显著增加叶面积[15]。麻栎和栓皮栎的叶脉间距离随着年均温度的降低而减小,可能是由于在低温这种不利条件下植物需要减小叶脉间距离来增加叶脉对于叶片的支撑作用,也就是要增加额外的物理支撑结构投资[16]。但是本试验发现栓皮栎的网眼面积和单位面积的网眼数与年平均降水量均具有相关性,但是麻栎网眼面积和单位面积的网眼数却与年平均降水量不具有显著的相关性。这可能是因为麻栎在自然条件下受其他环境因素的综合作用而栓皮栎却受温度和降水的影响更为显著导致的[17-18]。
同时,本研究发现麻栎和栓皮栎的叶脉密度、节点数和网眼数随着年平均降水量的增加而减小,这可能因为在水分供应充足条件下,植物叶片会形成更多的叶脉来进行蒸腾作用,提高叶片代谢和水分循环利用效率。这与L.Sack[5]等比较了全球不同生态系统796个物种的叶脉网络功能性状所发现的结论一致,表明麻栎和栓皮栎的叶脉序在宏观尺度变化中符合自然选择压力下形成的生态适应策略。
树木的叶脉结构与大的气候环境整体适应性相关,但是这种外界生态环境影响通常是综合作用的,在局部小的环境条件(例如局部土壤特征、风速和一些生物因素)也对植物的生理构造产生相关影响,后续将对叶脉结构与局部立地环境的相关关系作进一步研究。本试验通过对叶片功能性状的科学量化,有助于更深入地研究其与气候变化的关系,下一步可以探究幼龄林的叶片脉络形成与成熟林的叶片脉络功能性状进行综合比对,以期完善对于叶脉网络性状的纵向研究。
[1] 李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学报,2005,22(Supp.1):118-127.
LI F L,BAO W K,Responses of the morphological and anatomical structure of the plant leaf to environmental change[J].Chanese Bulletin of Botany,2005,22(Supp.1):118-127.(in Chinese)
[2] ROYER D L,MILLER I M,PEPPE D J,etal.Leaf economic traits from fossils support a weedy habit for early angiosperms[J].American Journal of Botany,2010,97(3):438-45.
[3] SCOFFONI C,RAWLS M,MCKOWN A,etal.Decline of leaf hydraulic conductance with dehydration:relationship to leaf size and venation architecture[J].Plant Physiology,2011,156(2):832-843.
[4] BLONDER B,VIOLLE C,BENTLEY L P,etal.Venation networks and the origin of the leaf economics spectrum[J].Ecology Letters,2011,14(2):91-100.
[5] SACK L,SCOFFONI C.Leaf venation:structure,function,development,evolution,ecology and applications in the past,present and future[J].New Phytologist,2013,198(4):983-1000.
[6] 李乐,曾辉,郭大立.叶脉网络功能性状及其生态学意义[J].植物生态学报,2013,37(7):691-698.
LI L,ZENG H,GUO D L.Leaf venation functional traits and their ecological significance[J].Chinese Journal of Plant Ecology,2013,37(7):691-698.(in Chinese)
[7] BEERLING D J,FRANKS P J.Plant science:the hidden cost of transpiration[J].Nature,2010,464(464):495-496.
[8] ZHANG S B,GUAN Z J,SUN M,etal.Evolutionary association of stomatal traits with leaf vein density inPaphiopedilum,Orchidaceae[J].Plos One,2012,7(6):40080.
[9] SACK L,SCOFFONI C,MCKOWN A D,etal.Developmentally based scaling of leaf venation architecture explains global ecological patterns[J].Nature Communications,2012,3(837):199-202.
[10] 李晓鹏.枣叶脉序及其功能性状研究[D].杨陵:西北农林科技大学,2016.
[11] 孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能[J].植物生态学报,2007,31(1):150-165.
MENG T T,NI J,WANG G H.Plant functional traits,environments and ecosystem functioning[J].Journal of Plant Ecology,2007,31(1):150-165.(in Chinese)
[12] PEPPE D J,ROYER D L,CARIGLINO B,etal.Sensitivity of leaf size and shape to climate:global patterns and paleoclimatic applications[J].New Phytologist,2011,190:724-739.
[13] GIVNISH T J,BURKHARDT E L,HAPPEL R E.Carnivory in the bromeliad brocchinia reducta,with a cost/benefit model for the general restriction of carnivorous plants to sunny,moist,nutrient-poor habitats[J].The American Naturalist,1984,124(4):479-497.
[14] REICH P B,WALTERS M B,ELLSWORTH D S.Leaf life-span in relation to leaf,plant,and stand characteristics among diverse ecosystems[J].Ecological Monographs,1992,62(3):365-392.
[15] HUDSON J M G,HENRY G H R,CORNWELL W K.Taller and larger:shifts in arctic tundra leaf traits after 16 years of experimental warming[J].Global Change Biology,2011,17(2):1013-1021.
[16] WESTOBY M,FALSTER D S,MOLES A T,etal.Plant ecological strategies:some leading dimensions of variation between species[J].Annual Review of Ecology & Systematics,2003,33(1):125-159.
[17] 王金照,张文辉.不同生境下栓皮栎叶形态解剖的研究[J].西北林学院学报,2004,19(2):44-46.
WANG J Z,ZHANG W H.The research on form dissecting ofQuercusvariabilisleaf in different habitats[J].Journal of Northwest Forestry University,2004,19(2):44-46.(in Chinese)
[18] 周建云,林军,何景峰.栓皮栎研究进展与未来展望.西北林学院学报,2010,25(3):43-49.
ZHOU J Y,LIN J,HE J F.Review and perspective onQuercusvariabilisresearch[J].Journal of Northwest Forestry University,2010,25(3):43-49.(in Chinese)
[19] NAVARRO T,ELOUALIDI J,TALEB M S,etal.Leaf patterns,leaf size and ecologically related traits in high Mediterranean mountain on the moroccan high atlas[J].Plant Ecology,2010,210:275-290.
猜你喜欢
广东蚕业(2022年4期)2022-11-23 13:31:55
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
南方农业·下旬(2020年1期)2020-06-01 07:47:32
文萃报·周五版(2019年44期)2019-09-10 22:55:44
现代园艺(2019年24期)2019-01-07 01:19:36
现代农业科技(2009年21期)2018-09-03 00:00:00
农村百事通(2017年18期)2017-02-08 11:23:54
江苏丝绸(2014年3期)2014-08-15 00:49:06
杂文选刊(2014年5期)2014-04-24 03:59:18
