
氯化镉对本生烟草幼苗生长及蛋白质表达水平的影响
2018-01-18 09:07陆玉建石东里吴涛
湖北农业科学 2018年21期
陆玉建 石东里 吴涛

摘要:为探究氯化镉(CdCl2)对本生烟草(Nicotiana benthamiana)幼苗生长及蛋白质表达水平的影响,选用不同浓度的CdCl2处理本生烟草幼苗,测量其根长和茎长,提取本生烟草幼苗总蛋白质,利用SDS-PAGE进行检测。试验结果表明,低浓度CdCl2对本生烟草幼苗的生长具有促进作用;高浓度CdCl2则抑制了本生烟草幼苗的生长。当CdCl2浓度达到0.5 mmol/L时,幼苗的生长开始受到抑制;CdCl2浓度大于1.5 mmol/L时,幼苗大量变黄枯萎。SDS-PAGE顯示,低浓度CdCl2可提高植物总蛋白质的含量,而随着CdCl2浓度的提高,本生烟草总蛋白质的含量明显下降。
关键词:CdCl2;本生烟草(Nicotiana benthamiana);幼苗;总蛋白质;SDS-PAGE
中图分类号:S572 文献标识码:A
文章编号:0439-8114(2018)21-0087-04
DOI:10.14088/j.cnki.issn0439-8114.2018.21.021 开放科学(资源服务)标识码(OSID):
Effects of CdCl2 Treatment on the Growth and Protein Expression Level of
Nicotiana benthamiana Seedlings
LU Yu-jian,SHI Dong-li,WU Tao
(College of Biological and Environmental Engineering,Binzhou University/Shandong Provincial Engineering and Technology Research Center for Wild Plant Resources Development and Application of Yellow River Delta,Binzhou 256603,Shandong,China)
Abstract: In order to investigate the effects of CdCl2 treatment on the growth and protein expression of Nicotiana benthamiana,the seedlings were treated with CdCl2. The growth status of the seedlings was observed and the length of the root and stem was recorded. The protein of Nicotiana benthamiana were extracted by SDS extraction,and isolated by SDS-PAGE. A large number of experimental data showed that low concentration of CdCl2 could promote the growth of the seedlings,while the high concentration of CdCl2 inhibited the growth of the seedlings. When the concentration of CdCl2 reached 0.5 mmol/L,the growth of seedlings began to be inhibited. If the concentration of CdCl2 was greater than 1.5 mmol/L,the seedlings turned yellow and withered. SDS-PAGE showed that low concentration of CdCl2 could increase the total protein content of the plant,but with the increase of CdCl2 concentration,the total protein content of Nicotiana benthamiana significantly decreased.
Key words: CdCl2; Nicotiana benthamiana; seedling; total protein; SDS-PAGE
伴随着经济发展,重金属对土壤的污染日益严重,已成为亟待解决的世界性难题。重金属元素镉(Cd)对生物体具有较强毒性,迁移性强且易积累[1,2]。Cd在土壤中主要是以化合物氯化镉(CdCl2)的形式存在,当浓度超过一定值时,生物体的生长受到抑制,表现出了一系列的CdCl2毒害效应[3,4]。土壤Cd污染对植物的形态结构、生长发育、细胞结构以及植物体寿命有着重要的影响[5,6]。Cd能阻碍植物体叶绿素合成,影响植物对CO2的吸收和O2的排放能力,使其生物合成代谢能力下降[7-9]。当土壤中Cd的含量不断升高时,植物气孔的开放受到抑制,从而影响植物的蒸腾作用以及光合作用,造成植物体内的营养成分失调[10,11]。此外,Cd还可使植物中某些酶的活性丧失,破坏生物膜结构或引起细胞的氧化应激,导致活性氧自由基(ROS)增加,攻击生物体中细胞内蛋白质、脂质等生物大分子物质[12-15]。Cd在生物体中不易分解和转化,是一种极具危害性的土壤污染重金属,能够通过食物链逐级往上传递,并不断富集[5,10,11]。Cd可在人体内积累,对神经系统、代谢系统、骨骼系统以及消化系统等造成严重的伤害,具有极强的致癌、致畸作用,严重危害人类的健康[14]。近年来,重金属Cd污染的治理已成为国内外学者研究的热点问题。土壤中的Cd主要是以Cd2+和螯合物的形式进入植物根系细胞,利用H+/阳离子反向运输器、P1B-ATP酶、ABC转运蛋白等转运到液泡中,经共质体运输进入根茎中的木质层、韧皮层以及中柱内,通过蒸腾作用将其运输到植物体其他的组织内[16-18]。
目前,有关重金属Cd对植物体的毒害机制尚不十分清楚。为此,本试验以模式植物本生烟草(Nicotiana benthamiana)为材料,采用不同浓度的CdCl2处理其幼苗,通过分析幼苗生长及蛋白质表达情况,研究CdCl2对植物体的毒害效应,以期为今后揭示相关的生理机制提供依据。
1 材料与方法
1.1 材料
本生烟草种子,由山东省黄河三角洲野生植物资源开发利用工程技术研究中心提供。
1.2 方法
1.2.1 本生烟草种子的消毒及培养 取本生烟草种子,用0.1%的升汞消毒10 min,无菌水洗涤3~4次,接种于MS基本培养基中进行培养。本生烟草的生长条件为16 h光照/8 h黑暗,湿度保持在60%~70%,温度控制在25 ℃左右,光照度为2 000 lx。
1.2.2 本生烟草幼苗的CdCl2处理 取培养10 d的本生烟草幼苗,转接到含不同浓度CdCl2(0、0.125、0.250、0.500、0.750、1.000、1.500和2.000 mmol/L)的MS基本培养基中,置于光照培养箱中进行垂直培养。
1.2.3 本生烟草幼苗根长和茎长的测量 每天观察幼苗生长状况,测量并记录本生烟草幼苗在不同浓度CdCl2处理时根和茎的长度,连续统计14 d。试验3次重复。
1.2.4 提取本生烟草幼苗的总蛋白质 取本生烟草幼苗,加入1 mL SDS提取液1(5%蔗糖,5% β-巯基乙醇,4% SDS)进行研磨,静置30 min。4 ℃,12 000 r/min离心15 min。取上清,加入8倍体积的丙酮, -20 ℃静置1 h以上。4 ℃,12 000 r/min离心15 min,留沉淀进行干燥。加入50 μL SDS提取液2(5%蔗糖,5% β-巯基乙醇,4% SDS,1% NP-40)重悬沉淀,煮沸3 min。4 ℃,12 000 r/min离心5 min,-80 ℃保存备用。
1.2.5 SDS-PAGE分离蛋白质 制备分离胶(T=10%)和浓缩胶(T=5%)。将蛋白样品上样。电泳初始电压为100 V,样品进入分离胶时,将电压调整为180 V。电泳结束后,考马斯亮蓝(R-250)染色。加入脱色液进行脱色,直到出现清晰的蛋白质条带。观察并拍照记录。
1.2.6 数据分析 采用Excel 2003和SPSS 19.0软件对所得数据进行统计分析,通过配对样本t检验进行显著性检测。
2 结果与分析
2.1 CdCl2处理对本生烟草幼苗生长状态的影响
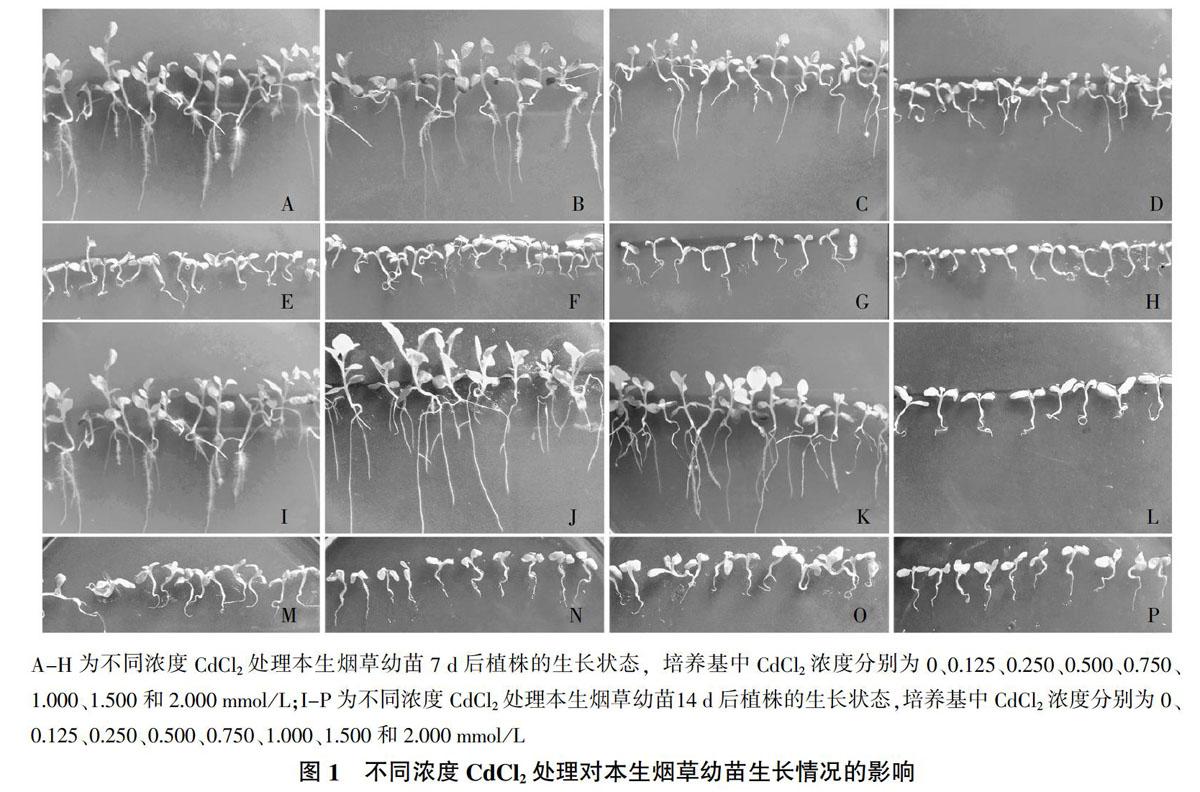
用不同浓度CdCl2处理本生烟草幼苗7 d后,观察本生烟草幼苗的生长情况。试验结果显示,与对照组相比(图1A),当培养基中CdCl2浓度为0.125~0.250 mmol/L时,幼苗的生长基本正常(图1B、图1C);当培养基中CdCl2浓度为0.500 mmol/L时,幼苗的生长开始受到抑制,生长速度下降(图1D);当CdCl2浓度达到0.750~1.000 mmol/L时,幼苗生长迟缓,出现叶片枯黄或植株枯萎现象(图1E、图1F);当CdCl2浓度超过1.000 mmol/L,叶片枯黄和植株枯萎更为明显,尤其当CdCl2浓度达到2.000 mmol/L,大部分本生烟草幼苗枯萎死亡,植株生长受到明显抑制(图1G、图1H)。
当CdCl2处理本生烟草幼苗14 d后,0.250 mmol/L CdCl2处理幼苗的对照组(图1I)和生长情况(图1K)基本相似;但0.125 mmol/L CdCl2处理的幼苗植株生长要优于对照组,表现出一定的促进作用(图1J);当培养基中CdCl2浓度达到0.500 mmol/L时,幼苗停止生长,大部分植株叶片枯黄(图1L);当培养基中CdCl2浓度为0.750~1.000 mmol/L时,幼苗植株个体小,大部分叶片枯黄和植株枯萎(图1M、图1N);当CdCl2浓度大于1.000 mmol/L时,叶片枯黄和植株枯萎现象更为明显,随着CdCl2浓度继续增加,本生烟草幼苗全部枯萎死亡(图1O、图1P)。
2.2 CdCl2处理对本生烟草幼苗茎和根长度的影响
对CdCl2处理7 d后的本生烟草幼苗的茎长进行数据分析,结果如图2A所示。当培养基中CdCl2浓度为0.125~0.250 mmol/L时,幼苗茎的长度在0.7~1.0 cm,和对照组区别不大。其中CdCl2浓度为0.125 mmol/L时,茎的生長速度稍快,接近1.0 cm。当CdCl2的浓度大于0.5 mmol/L时,茎的长度明显递减,和对照组差异显著。尤其当CdCl2浓度达到2.000 mmol/L时,和对照组差异极显著。当CdCl2处理14 d时(图2B),0、0.125、0.250 mmol/L CdCl2浓度下本生烟草幼苗茎继续生长,长度相近,生长速度逐渐变慢;当CdCl2浓度超过0.500 mmol/L时,其茎的伸长基本停滞,长度在0.4~0.6 cm,和对照组之间存在极显著差异。通过分析试验数据可知,低浓度CdCl2对本生烟草幼苗茎的生长有一定的促进作用,但随着CdCl2浓度的不断增加,茎的长度越来越短,生长受到了明显的抑制。
对CdCl2处理7 d后的本生烟草幼苗的根长进行数据分析,结果如图2C所示。0.125 mmol/L CdCl2处理的本生烟草幼苗主根的长度最长,接近1.2 cm,生长速度最快;0.250 mmol/L浓度下的本生烟草幼苗的主根长度和对照组相似,长度在0.9~1.0 cm;当CdCl2浓度大于0.500 mmol/L时,其主根长度明显变短,和对照组之间存在极显著的差异。当CdCl2处理14 d时(图2D),在0、0.125、0.250 mmol/L CdCl2浓度下,本生烟草幼苗根系继续生长。其中,当培养基中CdCl2浓度为0.125 mmol/L时,幼苗主根生长速度较快,长度接近2.5 cm,和对照组之间差异极显著。当CdCl2浓度大于0.500 mmol/L时,其根系的生长已基本停滞,长度在0.4~0.6 cm,和对照组之间差异极显著。上述结果表明,低浓度CdCl2可促进本生烟草幼苗根的生长,伴随着CdCl2浓度的不断增加,幼苗主根的长度越来越短,生长受到了抑制。
2.3 CdCl2处理对本生烟草幼苗蛋白质含量的影响
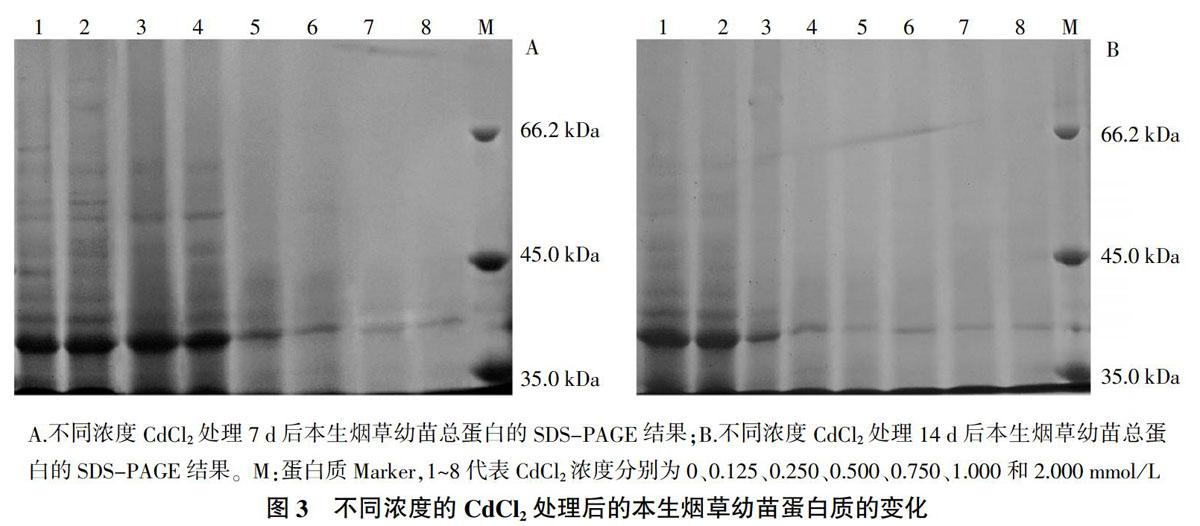
不同浓度CdCl2处理本生烟草幼苗7 d和14 d后,利用SDS法提取幼苗植株总蛋白质,通过SDS-PAGE分离蛋白质,分析蛋白质含量的变化,结果如图3所示。当CdCl2处理本生烟草幼苗7 d时,与对照组相比,0.125~0.500 mmol/L CdCl2处理的本生烟草幼苗其蛋白质合成基本正常;当CdCl2浓度大于0.500 mmol/L时,蛋白质合成受到明显抑制,含量有所降低。当CdCl2处理本生烟草幼苗14 d后,与对照组相比,0.125 mmol/L CdCl2处理下的幼苗其蛋白质含量变化不明显;0.250~0.500 mmol/L CdCl2处理的本生烟草幼苗蛋白质含量明显降低;CdCl2浓度超过0.500 mmol/L时,蛋白质含量继续减少,本生烟草幼苗蛋白质合成受到显著抑制。
3 小结与讨论
土壤中的多种重金属对于植物体具有毒害效应。CdCl2作为镉的化合物存在于自然界中,危害着植物的生长,是一种土壤中比较常见的重金属污染物,对农业生产以及生态平衡造成了不可估计的损失和影响[19,20]。为了研究CdCl2对植物生长发育的影响,试验中,利用不同浓度的CdCl2处理本生烟草幼苗,以观察统计其根、茎的生长状况,并检测蛋白质含量的变化。结果显示低浓度的CdCl2对本生烟草幼苗的生长有促进作用,而高浓度的CdCl2则抑制其生长。当CdCl2处理本生烟草幼苗7 d时,对照组和低浓度(0.125~0.250 mmol/L)CdCl2处理的幼苗生长状态没有明显区别;但高浓度(0.500~2.000 mmol/L)CdCl2处理的幼苗生长比较缓慢,植株矮小甚至出现了叶片枯黄的现象,和对照组差异比较明显,尤其当CdCl2浓度大于0.750 mmol/L时,幼苗的生长受到了极大抑制,随着CdCl2处理时间的延长,抑制作用愈发明显。当CdCl2处理本生烟草幼苗14 d时,随着CdCl2浓度(0.500~2.000 mmol/L)的增加,叶片枯黄和植株枯萎的幼苗数量不断增加,存活率降低,CdCl2对本生烟草幼苗的毒害效应愈加明显,其抑制作用愈加显著。而低浓度(0.125~0.250 mmol/L)CdCl2处理的本生烟草幼苗则继续生长,尤其是在0.125 mmol/L的浓度下其根系生长较快,长度最长,表明低浓度CdCl2对本生烟草生长具有明显的促进效应。
通过SDS-PAGE分离不同浓度CdCl2处理时本生烟草幼苗蛋白质并分析其含量变化,试验数据显示,随着CdCl2浓度的升高,其蛋白质条带数目由多到少,颜色也由深变浅。而CdCl2处理幼苗时间不同,其蛋白质合成以及含量也不同。随着CdCl2处理时间的延长,蛋白质条带数目由多到少,颜色也由深变浅。但0.125 mmol/L CdCl2处理的幼苗其蛋白质含量和对照组没有明显区别。SDS-PAGE结果进一步证实,不同处理时间、不同浓度CdCl2对本生烟草幼苗生长发育有着不同的毒害效应。试验初步分析了CdCl2处理对本生烟草幼苗生长及蛋白质表达水平影响,研究结果可以为今后揭示镉胁迫的生理机制以及污染土壤的植物修复提供参考。
参考文献:
[1] 陈艳芳.苋菜品种重金属Cd、Pb累积特性与根系重金属关键转运蛋白基因的表达及生长季节的关系[D].广州:暨南大学,2015.
[2] 时 萌,王芙蓉,王棚涛.植物响应重金属Cd胁迫的耐性机理研究进展[J].生命科学,2016(4):504-512.
[3] 王松良.芸苔属蔬菜重金属累积特性及抗Cd基因的差异表达与克隆[D].福州:福建农林大学,2004.
[4] 苏向楠.NO对Cd胁迫下紫花苜蓿幼苗氧化损伤与Cd积累的调控作用及其机制[D].兰州:兰州交通大学,2015.
[5] 张 玲.拟南芥水杨酸突变体对Cd胁迫的应答[D].沈阳:沈阳师范大学,2012.
[6] 姚诗音.超富集植物青葙对土壤镉的修复性能及强化措施研究[D].广西桂林:桂林理工大学,2017.
[7] 徐汉卿.商陆重金属代谢相关基因响应锰、Cd胁迫的表达分析[D].北京:中国农业科学院,2013.
[8] 韩承华.Cd、Pb、Cu、Zn对3种水生叶菜生长的影响以及Se缓解效果的研究[D].江苏扬州:扬州大学,2017.
[9] 罗 娅,黄翔鹄,李长玲.Hg-Cd联合胁迫对卵叶喜盐草光合色素和丙二醛含量及抗氧化酶活性的影响[J].广东海洋大学学报,2017,37(4):34-40.
[10] 李洪敬.氯化镉对霞多丽(Vitis vinifera)幼苗生理影响及Cd的形态分布[D].重庆:西南大学,2011.
[11] CHIEN H F,KAO C H. Accumulation of ammonium in rice leaves in response to excess cadmium[J].Plant Sci,2000,156(1):111-115.
[12] 陳忠翔.重金属镉胁迫细胞氧化毒理分子机制研究[D].上海:东华大学,2014.
[13] 李春喜,张志娟,张黛静,等.Cu2+、Cd2+胁迫对小麦幼根SOD活性及其基因表达的影响[J].中国农业科技导报,2011,13(4):92-98.
[14] 于祎飞.土壤Cd污染及农艺调控对苹果树体Cd积累影响的研究[D].河北保定:河北农业大学,2011.
[15] 宋 婕,王鹤潼,崔伟娜,等.Cd胁迫诱导拟南芥幼苗DNA损伤分析[J].农业环境科学学报,2017,36(4):635-642.
[16] 薛 亮,刘建锋,史胜青,等.植物响应重金属胁迫的蛋白质组学研究进展[J].草业学报,2013,22(4):300-311.
[17] DALCORSO G,FARIANTI S,MAISTRI S,et al. How plants cope with cadmium:staking all on metabolism and gene expression[J].Joumal of Integrative Plant Biology,2008,50(10):1268-1280.
[18] 岳 昊,杨友才.镉对不同品种烟草生长及生理生化指标的影响[J].山东农业科学,2015(4):76-79.
[19] 陈冬月.EDTA强化盐生植物修复Pb、Cd与次生盐渍化复合污染土壤[D].重庆:西南大学,2017.
[20] 赵淑玲,王 瀚,王让军,等.Cd2+对花椰菜种子的萌发及幼苗生理生化的影响[J].种子,2018,37(1):100-102.
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06
今日农业(2021年10期)2021-11-27
矿产综合利用(2020年1期)2020-07-24
中成药(2018年8期)2018-08-29
浙江中西医结合杂志(2017年2期)2017-01-12
当代化工研究(2016年9期)2016-03-20
当代化工研究(2016年6期)2016-03-20
中国资源综合利用(2016年3期)2016-01-22
中国火炬(2015年12期)2015-07-31
中国火炬(2013年3期)2013-07-24
