
氮素形态及水分胁迫对水稻根系水分吸收的影响机制
2018-01-18 09:07杨秀霞燕辉商庆银周春火郭世伟
湖北农业科学 2018年21期
关键词:水稻
杨秀霞 燕辉 商庆银 周春火 郭世伟
摘要:采用室內营养液培养及PEG6000模拟水分胁迫的方法,研究不同形态氮素营养(铵态氮和硝态氮)和水分条件对4种基因型水稻汕优63、扬稻6号、86优8、武运粳7号根系生理特性和水分吸收途径的影响。结果表明,在2种水分条件下,供铵营养水稻整株生物量均显著高于硝营养水稻,在水分胁迫条件下表现出较强的抗旱性。水分胁迫条件下供铵态氮营养水稻的根系具有较高的根系活力、吸收面积和水分吸收能力。在非水分胁迫条件下,供铵态氮和硝态氮营养水稻根系水分吸收和运输途径相同;水分胁迫后,硝营养水稻的水分运输途径以运输阻力较大的质外体途径为主,尤其在粳稻品种中表现显著。就不同基因型水稻品种而言,籼稻品种在供铵态氮营养时地上部干重显著高于粳稻品种,且籼稻品种在水分胁迫条件下供铵态氮营养时根系总吸收面积、活跃吸收面积和根系活力也显著高于粳稻品种,表现出较强的水分吸收能力。
关键词:水分胁迫;氮素形态;水稻(Oryza sativa L.);水分吸收
中图分类号:S511 文献标识码:A
文章编号:0439-8114(2018)21-0051-06
DOI:10.14088/j.cnki.issn0439-8114.2018.21.012 开放科学(资源服务)标识码(OSID):
Effects of Nitrogen Forms and Water Stress on the Water Absorption of Root Systems Among the Different Rice Cultivars
YANG Xiu-xia1a,2,YAN Hui1a,SHANG Qing-yin1b,ZHOU Chun-huo1a,GUO Shi-wei2
(1a.College of Land Resources and Environmental Sciences;1b.College of Agronomy, Jiangxi Agricultural University,Nanchang 330045,China;
2.College of Resources and Environmental Sciences,Nanjing Agricultural University,Nanjing 210095,China)
Abstract: In order to study the effects of different nitrogen forms and water stress on physiological characteristics and water absorption pathway of root system,hydroponic experiment was conducted in a greenhouse. Water stress was simulated by treatment with polyethylene glycol(PEG6000). Four rice(Oryza sativa L.) genotypic varieties (Shanyou 63,Yangdao 6,86you 8 and Wuyunjing 7) were utilized in the experiments. The results showed that regardless of water conditions,rice biomass was higher in the plants supplied with ammonium nutrition than that in nitrate nutrition. Under water stress condition, root activity, absorption area and water uptake ability were higher in the plants supplied with ammonium nutrition than that in nitrate nutrition. Regardless of water status, water uptake and transport were mainly through cell-cell passway in the plants supplied with ammonium nutrition; as for the plants supplied with nitrate nutrition, they were mainly through cell-cell pathway under non-water stress condition and throught apoplastic pathway under water stress condition. Compared with non-water stress condition, water stress significantly stimulated root growth in both N forms, and the stimulation was higher in nitrate nutrition. As to indica rice,water stress had no negative effects on shoot biomass and tillers in the plants supplied with ammonium nutrition, while significantly decreased them in nitrate nutrition. As to japonica rice, water stress significantly decreased shoot biomass and tillers in both N forms.
Key words: water stress; nitrogen form; rice (Oryza sativa L.); water uptake
水稻(Oryza sativa L.)作为农业中的耗水大户,每年用水量约占农业用水量的70%[1],而其中大量的灌溉水在地表蒸发或者渗漏流失,水资源浪费现象十分严重。随着中国水资源的日益紧缺,开展水稻抗旱生理研究受到越来越多的重视[2]。通过对水稻水分代谢的研究,证明除了生态用水浪费严重外,水稻的生理水分利用效率也十分低下。究其原因,主要是由于水稻叶片气孔开放程度很高[3],根系及木质部的水流导度(Hydraulic conductance)又很低造成的[4]。因此,即使是在田间灌水条件下,也会出现叶片因缺水而萎蔫的现象。所以理解和探讨水稻地上部需水与地下部供水的关系,对调节水稻的水分代谢具有重要意义。
根系作为水稻吸收水分的重要器官,其活力受氮素营养和水分供应状况的显著影响。周毅等[5]研究表明,供铵态氮营养有利于水分胁迫下水稻根系的生长,从而减轻水分胁迫对水稻生长的抑制作用。吴芳等[6]研究发现,供铵态氮营养水稻根系干重显著低于供硝态氮处理,但单位根表面积的水分吸收与根系伤流液量比供硝态氮处理高。这些研究为分析不同氮素形态与水稻根系抗旱能力之间的关系奠定了基础,但由于植物水分生理及其與其他生理过程的复杂联系,对于不同氮素形态改变植物水分利用状况的生理机制还有待深入研究。一般认为,植物根系对水分的吸收主要通过3种途径:质外体途径、共质体途径和跨细胞途径,其中后两种途径又统称为细胞-细胞途径[7-9]。据报道,细胞-细胞途径的水分运输有70%~90%通过水通道蛋白(原生质膜和液泡膜上的专一性水分运输蛋白)进行。然而哪种吸收途径对植物抗旱性起主要作用是国际上一直争论的话题。宋娜等[10]研究表明,供铵态氮和硝态氮营养水稻根系水分吸收差异的原因可能在于根系水通道蛋白活性的不同。但上述研究主要集中在对单一品种的分析,而不同氮素形态对水稻根系水分吸收的影响是否存在品种间差异仍缺乏了解。
本试验以4种不同基因型水稻汕优63、扬稻6号、86优8和武运粳7号为供试材料,采用营养液添加PEG(聚乙二醇6000)模拟水分胁迫、HgCl2抑制水通道蛋白活性的方法,研究不同供氮形态下水稻根系生理特性和水分吸收,探讨不同氮素形态营养对水稻水分吸收、利用及生长状况的影响,以期为水稻苗期合理施肥、提高水分利用率、发展水稻节水栽培技术提供理论依据。
1 材料与方法
1.1 供试品种
供试水稻品种为汕优63(杂交籼稻)、扬稻6号(常规籼稻)、86优8(杂交粳稻)、武运粳7号(常规粳稻)。
1.2 试验设计
供氮形态为铵态氮[NH4+-N,以(NH4)2SO4形式供应]、硝态氮[NO3--N,以Ca(NO3)2形式供应],供氮浓度均为40 mg/L。营养液采用国际水稻所(IRRI)的常规营养液配方[11],并略作改进。此外,添加Na2SiO3使营养液中的SiO2浓度为0.1 mmol/L,并加入1 mg/L双氰胺(DCD)作为硝化抑制剂。2种水分处理:正常水分条件,模拟水分胁迫条件[在营养液中添加100 g/L聚乙二醇6000(PEG 6000),渗透势约相当于-0.15 MPa]。试验共设置4个处理,即铵态氮处理、硝态氮处理、铵态氮加PEG 6000处理、硝态氮加PEG 6000处理,分别以A、N、AP、NP表示。
种子消毒后均匀铺于育苗框中,常温条件下避光发芽;培养至2叶1心时,选取长势一致的幼苗移栽至不透光的塑料桶中(容积为3 L),每桶移栽6穴,每穴2株。在1/2浓度完全营养液(NH4+-N和NO3--N浓度均为10 mg/L)中缓苗至3叶1心,完全营养液(NH4+-N和NO3--N浓度均为20 mg/L)预培养3 d后进行不同形态氮素处理,营养液pH每天调节至5.50±0.05,每3 d更换1次营养液。处理9 d后(至不同处理间出现明显的生物学性状差异)进行水分胁迫处理,每处理重复5次,再培养12 d进行各项指标的测定。
1.3 测定项目
1.3.1 生物量 样品分地上部和根系两部分采集。先用去离子水清洗干净,用吸水纸吸干表面水分后,测定鲜重;然后立即于105 ℃烘箱中杀青30 min后再降温至70~80 ℃并烘至恒重,测定干重。
1.3.2 根系形态指标测定 根系形态参数采用LA1600+根系扫描仪成像。成像前,于成像扫描仪配套的凹槽中选择大小接近的根系加约1/2水,将根系浸没其中,用木质牙签或徒手将根系分开以尽量减少对根系的伤害。成像后,选择目标根系区域,通过WinRHizo2003b分析图像参数,包括根尖数、根长、根体积和根表面积。
1.3.3 根系活力的测定 根系活力的测定采用苯基四氮唑(TTC)还原法[12]:称取适量新鲜根系,放入0.4%TTC溶液和0.1 mol/L磷酸缓冲液中,在37 ℃暗箱中保温1 h后,加入2 mol/L硫酸终止反应。取出处理根系,研磨,用乙酸乙酯浸提,在722型分光光度计上485 nm下读取吸光度。以单位鲜重根系还原的TTC量(TTC还原强度)表示,并计算单株根系TTC的还原强度。
1.3.4 根系总吸收面积和活跃吸收面积的测定 根系总吸收面积和活跃吸收面积采用次甲基蓝吸附法进行测定:首先测得待测根系体积,之后顺次放入3个盛放等量0.002 mol/L次甲基蓝溶液的烧杯中,每杯浸1.5 min。从3个烧杯中各取1 mL溶液,稀释10倍,在722型分光光度计上660 nm下读取吸光度。计算方法:活跃吸收表面积(m2)=(原液浓度-第三次吸收后浓度)×所用溶液量×1.1[13]。
1.3.5 单位根系水分吸收 10%PEG模拟水分胁迫处理结束后,通过称重差减法测定2 h内(9:00~ 11:00)水稻水分吸收量,结束后称取根系鲜重量。同时在另一批对应处理的营养液中加入预先配好的1 mmol/L HgCl2溶液(使营养液中HgCl2终浓度为0.1 mmol/L),通过称重差减法测定2 h内水稻幼苗的水分吸收速率。
1.4 数据处理
所有数据运用JMP10.0(SAS Institute,Cary,NC,USA,2011)和Microsoft Excel 2016软件进行统计分析,所有数据采用LSD方法进行多重比较。
2 结果与分析
2.1 不同供氮形态及水分胁迫对水稻苗期生长的影响
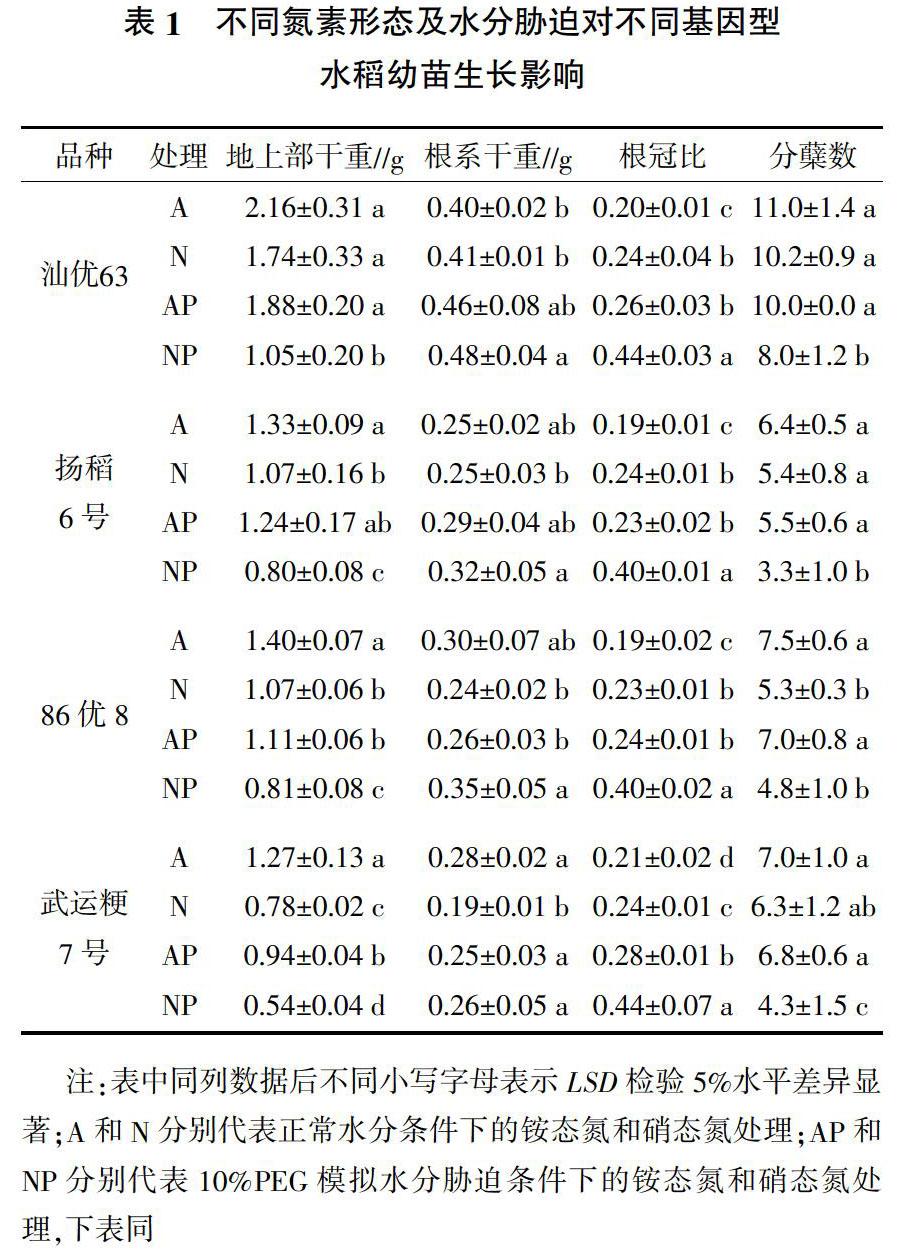
在相同水分条件下,不同供氮形态对水稻植株地上部干重均具有显著影响,除汕优63外,供铵态氮营养水稻地上部干重显著高于供硝态氮处理(表1)。水分胁迫对水稻植株地上部干重的影响与品种和供氮形态有关。就籼稻品种汕优63、扬稻6号而言,水分胁迫对供铵态氮营养水稻植株地上部干重影响较小,但显著降低了供硝态氮营养水稻植株地上部干重。就粳稻品种86优8和武运粳7号而言,水分胁迫后供铵态氮和硝态氮营养水稻植株地上部干重均显著降低。此外,由表1可知,在非水分脅迫条件下,供铵态氮和硝态氮营养水稻单株分蘖数没有显著差异(除86优8外);水分胁迫后,供硝态氮营养水稻分蘖数显著降低(除86优8外),但对供铵态氮营养水稻无影响。由此可见,在水分胁迫条件下,供铵态氮营养籼稻比粳稻表现出更强的抗旱性。
由表1可知,在非水分胁迫条件下,供铵态氮和硝态氮营养水稻根系干重没有显著差异(除武运粳7号外)。水分胁迫后,供铵态氮营养水稻根系干重没有显著变化,但供硝态氮营养水稻根系干重显著增加,且对粳稻品种的促进作用显著大于籼稻品种。与非水分胁迫处理相比,供硝态氮营养粳稻品种在水分胁迫后根系干重分别增加46%(86优8)和37%(武运粳7号),而籼稻品种仅增加17%(汕优63)和28%(扬稻6号)。此外,水分胁迫条件下,供铵态氮和硝态氮营养水稻根冠比均显著升高;但无论是在非水分胁迫还是在水分胁迫条件下,供硝态氮营养水稻根冠比均显著高于供铵态氮营养水稻。
2.2 不同供氮形态及水分胁迫对水稻根系形态的影响
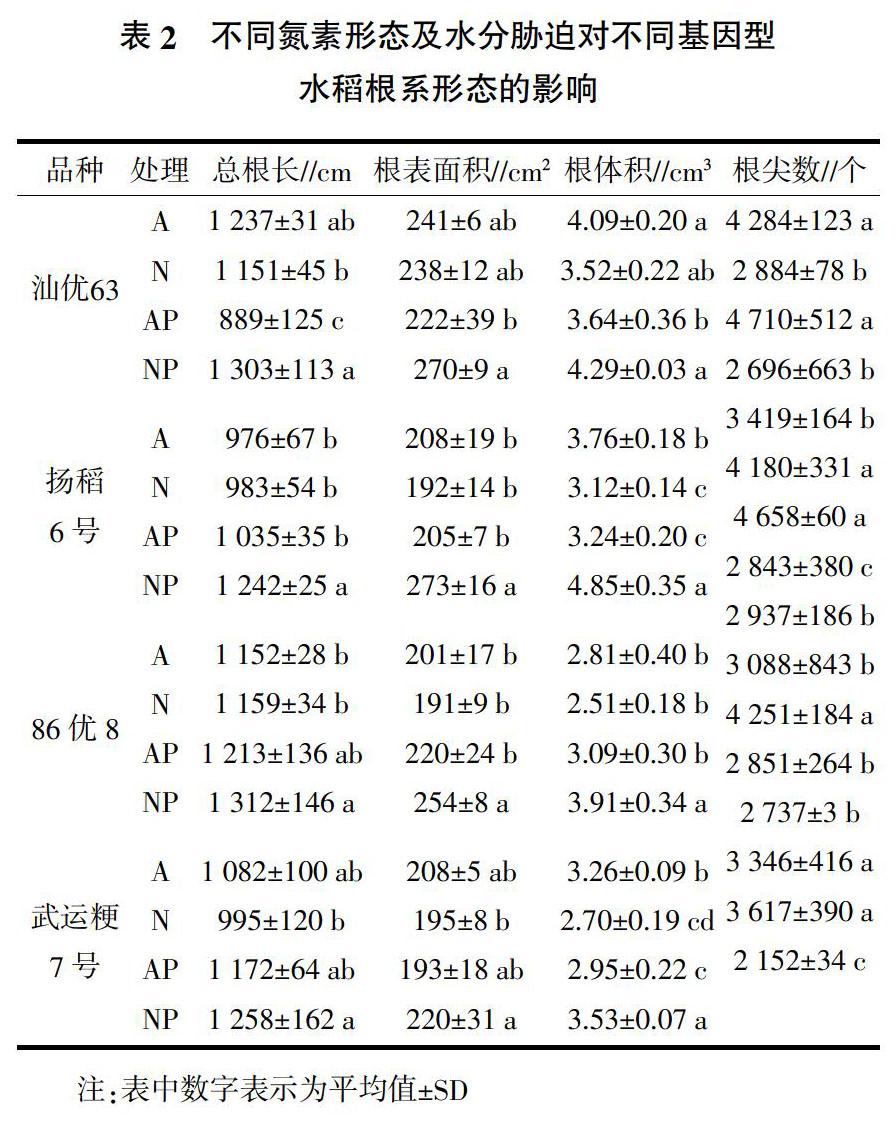
在水分胁迫条件下,除汕优63外,供硝态氮营养水稻根系总根长、根表面积和根体积均显著高于非水分胁迫处理(表2);而供铵态氮营养水稻根系总根长和根表面积没有显著变化,但其根体积显著低于非水分胁迫处理(除86优8外)。此外,水分胁迫条件下,供铵态氮营养水稻根尖数显著增加(除汕优63外),而供硝态氮营养水稻根尖数呈降低的趋势,其中扬稻6号和武运粳7号显著低于非水分胁迫处理。
2.3 不同供氮形态及水分胁迫对水稻根系吸收面积的影响
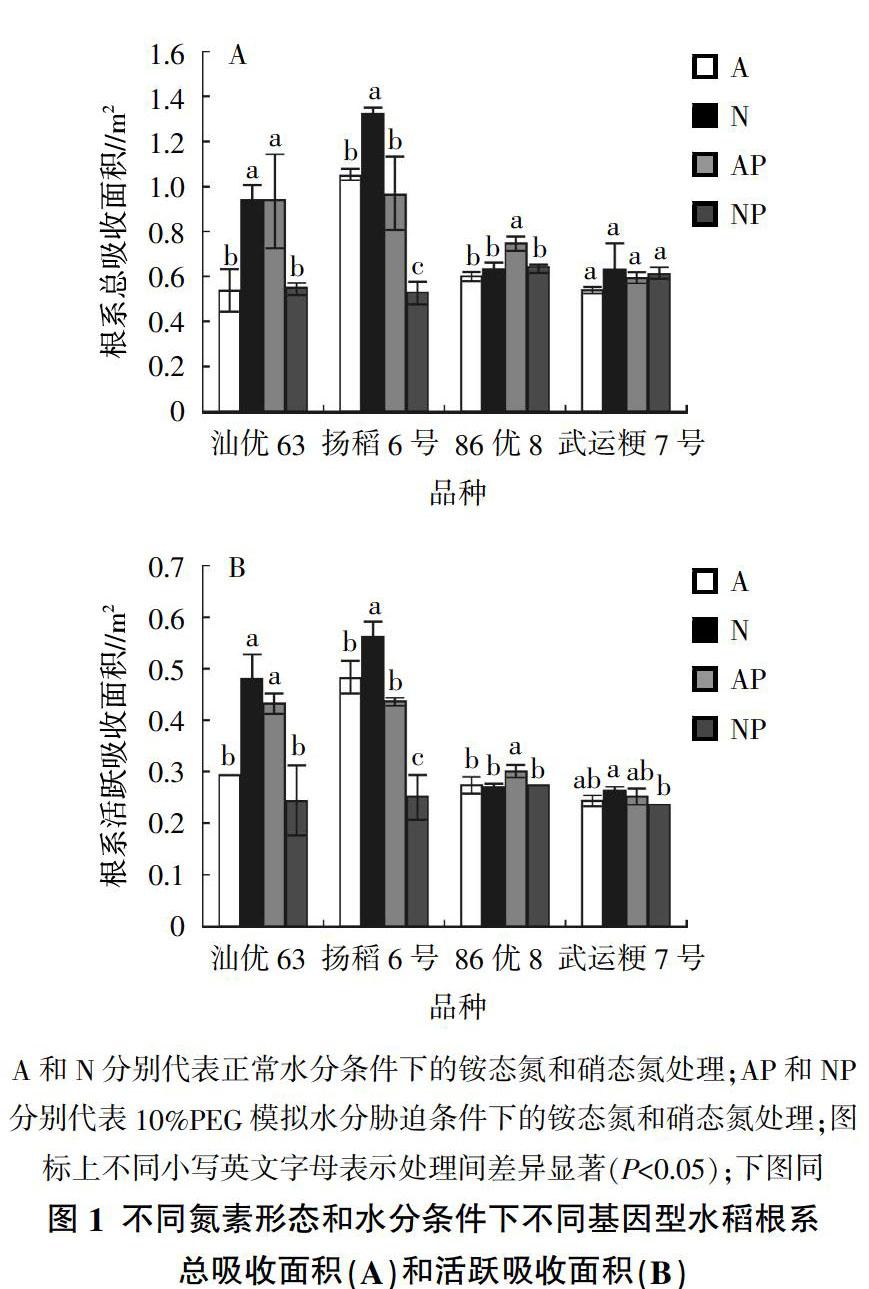
由图1可知,不同供氮形态及水分胁迫对水稻根系吸收面积的影响存在品种间差异。就籼稻品种汕优63和扬稻6号而言,非水分胁迫条件下,供铵态氮营养籼稻根系的总吸收面积(图1A)和活跃吸收面积(图1B)均显著低于供硝态氮营养籼稻处理;水分胁迫条件下,供铵态氮营养籼稻吸收面积显著升高(汕优63)或没有显著变化(扬稻6号),而供硝态氮营养籼稻的根系吸收面积则显著降低,其中汕优63根系总吸收面积和活跃吸收面积分别比非水分胁迫处理下降42%和49%,扬稻6号分别下降60%和55%。
就粳稻品种86优8、武运粳7号而言,非水分胁迫条件下,供硝态氮和铵态氮营养水稻之间根系的总吸收面积(图1A)和活跃吸收面积(图1B)均无显著差异。与非水分胁迫条件相比,水分胁迫显著提高了杂交粳稻86优8供铵态氮营养处理的根系总吸收面积和活跃吸收面积,但对供硝态氮营养处理无影响。而对于常规粳稻武运粳7号来说,水分胁迫对供铵态氮和硝态氮营养水稻根系总吸收面积均无影响,但供硝态氮营养水稻根系的活跃吸收面积显著降低。
2.4 不同供氮形态及水分胁迫对水稻根系活力的影响
由图2可知,与非水分胁迫条件相比,除武运粳7号外,水分胁迫显著降低了供铵态氮和硝态氮营养水稻根系活力(即TTC还原强度)。无论在非水分胁迫条件还是在胁迫水分条件下,供铵态氮营养水稻的根系活力均显著高于硝态氮营养水稻(除武运粳7号外)。因此,在水分胁迫条件下,供铵态氮营养有利于增强水稻根系活力。
2.5 不同供氮形态及水分胁迫对水稻根系水分吸收的影响
水分胁迫显著影响水稻根系水分吸收(表3)。在非水分胁迫条件下,供铵态氮和硝态氮营养水稻之间单位根系吸水量无显著差异(汕优63除外);模拟水分胁迫处理后,供铵态氮和硝态氮营养水稻根系水分吸收显著下降,但不同供氮形态下降幅度不同。在供铵态氮营养条件下,汕优63、扬稻6号、86优8和武运粳7号根系水分吸收分别下降30%、23%、22%和26%,供硝态氮营养条件下则分别下降40%、44%、35%和40%。可见,水分胁迫条件下,供铵态氮营养可有效缓解水稻根系水分吸收能力的显著下降。
HgCl2是水通道蛋白的专性抑制剂。通过向营养液添加0.1 mmol/L HgCl2后,不同供氮形态和水分处理条件下水稻单位时间内单位根系水分吸收量均呈下降的趋势(表3)。通过计算添加HgCl2后,水稻根系水分吸收下降百分率可推测不同处理条件下水稻根系通过水通道蛋白跨膜运输途径吸收水分的比例。从表3可以看出,在非水分胁迫条件下添加HgCl2处理后,供铵态氮和硝态氮营养水稻根系水分吸收下降比例差异较小;在水分胁迫条件下添加HgCl2处理后,供铵态氮营养水稻根系水分吸收下降比例变化较小,而供硝态氮营养水稻根系水分吸收下降比例显著降低(扬稻6号除外),尤其是粳稻品种(86优8和武运粳7号)。据此可推测,水分胁迫条件下,供铵态氮营养可能有助于维持水稻根系较高的水通道蛋白活性。
3 讨论
3.1 不同供氮形态及水分胁迫对水稻苗期生长的影响
根系是植物吸收水分的重要器官,植物对水分的吸收主要由根系的形态、根系活性和吸收面积以及根系吸收水分的方式和途径决定。干旱条件下,由于水稻生长对逆境产生的抗性以及根系生长的向水性,促进了根系生长,并向纵深和广处延伸,特别是促进了吸水根的生长[14]。然而也有研究表明,在轻度水分胁迫条件下,光合同化产物在根系和地上部的分配不受影响,而在重度胁迫条件下,向根系运输的干物质量甚至出现减少的现象[15]。本试验结果表明,与非水分胁迫条件相比,水分胁迫后供硝态氮营养水稻根系干重、总根长、根体积及根表面积均显著增加,而对供铵态氮营养水稻根系生长无影响。从水稻根尖数量来看,水分胁迫显著增加了供铵态氮营养水稻根尖数。由此可见,水分胁迫对供硝态氮营养根系的影响主要表现在根系长度的增加;而对供铵态氮营养水稻根系的影响在于根尖数的增加。
就地上部生长而言,水分胁迫条件下,供铵态氮营养水稻地上部干重和分蘖数均显著高于供硝态氮营养水稻。在2种水分条件下,供硝态氮营养水稻的根冠比均高于供铵态氮营养水稻。在水分胁迫下,供硝态氮营养可以促进水稻根系的发育,但却不利于充分发挥根系抗旱能力。据报道,在干旱条件下,较大的根冠比虽然有利干植物抗旱[16,17],但是过分庞大的根系由于消耗了过多的同化物反而会影响植物的抗旱性[18,19]。
3.2 不同供氮形态及水分胁迫对水稻根系活力的影响
根系活力泛指根系的吸收能力、合成能力、氧化能力和还原能力等,是一种客观地反映根系生命活动能力的生理指标[20]。根系总吸收面积直接反映根系吸收水分和养分的能力,根系活跃吸收面积反映根系活力情况。本研究发现,不同供氮形态及水分胁迫对水稻根系吸收面积的影响存在品种间差异。非水分胁迫条件下,籼稻品种在供铵态氮时根系的总吸收面积和活跃吸收面积均显著低于供硝态氮处理,而粳稻品种在不同供氮形态下根系吸收面积均无显著差异;水分胁迫条件下,供铵态氮营养有利于提高或维持不同水稻品种根系总吸收面积和活跃吸收面积。
根系总吸收面积和活跃吸收面积虽然在一定程度上反映根系的活力,但毕竟不是直接指标。根系活力(TTC还原强度)测定的是与呼吸相关的琥珀酸脱氢酶,故TTC还原能力与根系呼吸作用有关,是檢测根系活力大小的直接指标[12]。本研究表明,水分胁迫后供铵态氮和硝态氮营养的4种基因型水稻的根系活力均有降低的趋势,但无论是否在水分胁迫条件下,供铵态氮营养水稻的根系活力均显著高于硝态氮营养水稻(武运粳7号除外)。因此,水分胁迫条件下,供铵态氮营养有利于增强水稻根系活力,从而保证根系吸收水分和养分的能力,以供应水稻地上部生长发育的需要。
3.3 不同供氮形态及水分胁迫对水稻根系水分吸收运输的影响
本试验结果表明,水分胁迫显著抑制水稻单位根系水分吸收,但在水分胁迫条件下供铵态氮营养水稻根系水分吸收能力显著高于硝态氮营养水稻,说明水分胁迫后供铵态氮营养对水稻水分吸收能力下降具有一定的缓解作用。这可能与不同氮素形态条件下根系内部结构存在较大差异有关。Ranathunge等[20]研究发现,细胞-细胞途径的水分运输有70%~90%通过水通道蛋白进行,而水通道蛋白是植物运输水分的主要方式[21]。这主要是由于水通道蛋白能降低水分跨膜运输时的膜阻力,有利于水分在细胞内的运输[22]。
HgCl2是水通道蛋白的专性抑制剂,通过向营养液中添加HgCl2可以抑制水通道蛋白的活性[23,24]。试验结果表明,添加HgCl2后水稻单位时间单位根系水分吸收量均呈下降的趋势。在非水分胁迫条件下,添加HgCl2处理后,供铵态氮和硝态氮营养水稻根系水分吸收下降比例相当;但在水分胁迫条件下,添加HgCl2处理后供硝态氮营养水稻根系水分吸收下降比例显著降低,而供铵态氮营养水稻下降比例变化不大。说明在非水分胁迫条件下,水稻在不同供氮形态时吸收水分的途径可能相似;但在水分胁迫条件下,供硝态氮营养水稻根系水分吸收途径可能以质外体途径为主,而供铵态氮营养水稻根系以水通道蛋白途径为主。这也可能正是水分胁迫条件下供铵态氮和硝态氮营养水稻根系水分吸收存在差异的主要原因。
3.4 不同基因型水稻的抗旱能力差异机理
不同基因型水稻对不同水分胁迫、不同铵硝配比的响应存在差异[25,26]。本试验结果表明,在水分胁迫条件下供铵态氮营养籼稻品种比粳稻品种表现出更强的抗旱性。首先,无论是供铵态氮营养还是硝态氮营养,水分胁迫后粳稻品种地上部干重均显著降低;但当供铵态氮营养时水分胁迫对籼稻品种植株地上部干重无影响。水分胁迫后,供硝态氮营养水稻根系干重显著增加,且对粳稻品种的促进作用显著大于籼稻品种。其次,籼稻品种在水分胁迫条件下供铵态氮营养时根系总吸收面积、活跃吸收面积和根系活力也显著高于粳稻品种。再次,在水分胁迫条件下,供硝态氮营养水稻根系水分吸收下降比例显著降低,尤其是粳稻品种。此外,在不同供氮形态及水分条件下,籼稻品种的根系活力均相对高于粳稻品种。由此可见,在水分胁迫条件下籼稻品种在供铵态氮营养时地上部干重显著增加可能与其较高的根系活力、吸收面积以及水分吸收能力有密切关系。因此,建议在发展水稻旱作时,着重考虑种植籼稻品种。
4 结论
水分胁迫条件下,与供硝态氮营养相比,供铵态氮营养水稻的根系具有较高的根系活力、吸收面积和水分吸收能力,从而保证了地上部对水分的需求。
在正常水分条件下,供铵态氮和硝态氮营养水稻根系水分吸收和运输途径相同;水分胁迫后,硝营养水稻的水分运输途径以运输阻力较大的质外体途径为主,尤其在粳稻品种中表现显著,这可能是水分胁迫后供硝态氮营养水稻根系水分吸收显著下降的主要原因。就不同基因型水稻品种而言,籼稻品种在供铵态氮营养时地上部干重显著高于粳稻品种,且籼稻品种在水分胁迫条件下供铵态氮营养时根系总吸收面积、活跃吸收面积和根系活力也显著高于粳稻品种,表现出较强的水分吸收能力。
参考文献:
[1] 茆 智.水稻节水灌溉及其对环境的影响[J].中国工程科学,2002,
4(7):8-16.
[2] 程建平,曹凑贵,蔡明历,等.水分胁迫与氮素营养对水稻生理特性的影响[J].干旱地区农业研究,2007,25(11):149-154.
[3] MAKINO A,SATO T,NAKANO H,et al. Leaf photosynthesis,plant growth and nitrogen allocation in rice under different irradiances[J].Planta,1997,203:390-398.
[4] MIYAMOTO N,STEUDLE E,HIRASAWA T,et al. Hydraulic conductivity of rice roots[J].Journal of Experimental Botany,2001,52:1835-1846.
[5] 周 毅,郭世伟,沈其荣.局部根系干旱条件下分蘖期水稻对供氮形态的生物学响应[J].水土保持学报,2005,19(6):169-173.
[6] 吴 芳,高迎旭,宋 娜,等氮素形态及水分胁迫对水稻根系生理特性的影响[J].南京农业大学学报,2008,31(1):63-66.
[7] 于秋菊,吴 锜,林忠平.植物水孔蛋白研究进展[J].北京大学学报(自然科学版),2002,38(6):855-866.
[8] 沈玉芳,王保莉,曲 东,等.水分胁迫下磷营养对玉米苗期根系导水率的影响[J].西北农林科技大学学报,2002,30(5):11-15.
[9] 李善菊,任小林.植物水分胁迫下功能蛋白的研究进展[J].水土保持研究,2005,12(3):64-69.
[10] 宋 娜,郭世伟,沈其荣.不同氮素形态及水分胁迫对水稻苗期水分吸收、光合作用及生长的影响[J].植物学通报,2007,24(4):477-483.
[11] LU Z,NEUMANN P M. Water-stressed maize,barley and rice seedlings show species diversity in mechanisms of leaf growth inhibition[J].Journal of Experimental Botany,1998,49(329):1945-1952.
[12] 张 雄.用TTC(紅四氮唑)法测定小麦根和花粉活力及其应用[J].植物生理学通讯,1982,(3):48-50.
[13] 高俊凤.植物生理学实验技术[J].西安:世界图书出版公司,2000.
[14] 周利民,罗怀彬,古璇清.水稻节水灌溉机理研究[J].广东水利水电,2003(2):22-24.
[15] ASCH F,DINGKUHN M,SOW A,et al. Drought-induced changes in rooting patterns and assimilate partitioning between root and shoot in upland rice[J].Field Crop Research,2005, 93:223-236.
[16] TURNER N C. Adaption to water deficits:A changeing perspective[J].J Aust Plant Physiol,1986,13:175-190.
[17] 张岁歧,山 仑.氮素营养对春小麦抗旱适应及水分利用的影响[J].水土保持研究,1995,2(1):31-35.
[18] 景蕊莲.作物抗旱研究的现状与思考[J].干旱地区农业研究,1999,17(2):79-85.
[19] 孙国钧,张 荣,李凤民,等.作物抗旱性、水分利用与籽粒产量的形成[J].资源科学,1999,21(5):31-36.
[20] RANATHUNGE K,KOTULA L,STEUDLE E,et al. Water permeability and reflection coefficient of the outer part of young rice roots are differently affected by closure of water channels (aquaporins) or blockage of apoplastic pores[J].Journal of Experimental Botany,2004,396,433-447.
[21] STEUDLE E,HENZLER T. Water channels in plants:Do basic concepts of water transport change?[J].Journal of Experimental Botany,1995,46:1067-1076.
[22] SMITH J A C,NOBEL P S. Water movement and storage in a desert succulent:Anatomy and rehydration kinetics for leaves of Agave deserti[J].Journal of Experiment Botany,1986,37:1044-1053.
[23] MAGGIO A,JOLY R J. Effects of mercuric chloride on the hydraulic conductivity of tomato root systems. Evidence for a channel-mediated water pathway[J].Plant Physiology,1995,109:331-335.
[24] SCHUTZ K,TYERMAN S D.Water channels in Chara coralline[J].Journal of Experimental Botany,1997,48(313):1511-1518.
[25] 段英华,张亚丽,沈其荣.增硝营养对不同基因型水稻苗期吸铵和生长的影响[J].土壤学报,2005,42(2):260-265.
[26] 李 勇,周 毅,郭世伟,等.铵态氮和硝态氮营养对水、旱稻根系形态及水分吸收的影响[J].中国水稻科学,2007,21(3):294-298.
