
模拟氮沉降对滨海湿地土壤微生物功能多样性的影响①
2018-01-17 06:50吴松芹汪成忠李梦莎
土壤 2017年6期
吴松芹,汪成忠,李梦莎
(1苏州农业职业技术学院,江苏苏州 215008;2 黑龙江省科学院自然与生态研究所,哈尔滨 150040)
土壤微生物是生态系统中的重要组成部分之一[1],是土壤元素循环与转化的主要动力之一[2-3],在生态系统功能以及全球生态系统地球化学循环中发挥着重要的作用[4-5]。土壤微生物群落多样性反映了土壤微生物群落组成和结构的变化[6],而通过对土壤微生物群落组成和结构的变化进行分析,能够为解决生态系统功能的变化、制定保护和合理利用湿地资源的措施提供理论基础[7-9]。因此,研究土壤微生物群落结构及其功能多样性,对评价土壤质量、生态系统功能和土壤元素循环具有重要意义[10-11]。
大气氮沉降改变了许多陆地生态系统类型的生物地球化学循环已是事实。了解氮沉降对湿地土壤微生物的影响及其机制对于湿地和环境管理具有重要意义。国外关于土壤微生物对氮沉降的响应研究亦是近几年才起步,大部分研究结果表明,长期氮沉降增加会给土壤微生物带来不利影响[12-13],但有关氮沉降对滨海土壤微生物的影响研究较少。本研究通过野外模拟试验,探讨氮沉降增加对滨海湿地土壤微生物的影响,为研究氮沉降对滨海湿地生态系统的影响提供参考。
1 材料与方法
1.1 试验材料
本研究试验样地位于江苏省盐城国家级自然保护区内。自然保护区位于江苏省盐城市境内,地理位置119°53′45″~121°18′12″ E,32°48′47″~34°29′28″ N,总面积约为 247 260 hm2,其中核心区面积 22 596 hm2,缓冲区面积 56 742 hm2,试验区面积 167 922 hm2。该区地处暖温带与北亚热带过渡地带,气候主要受到海洋与大陆性气候影响,年平均气温为 13.7~14.6℃,年平均降雨量 1 000 mm,雨量集中在夏季,冬天多干旱,夏季凉爽(3月至8月) ,冬季温暖(9月至次年2 月)。保护区的优势植物类型有眼子菜、狐尾藻、水车前、野菱和芡、扁杆草、草、糙叶苔草、盐蒿、獐茅和白茅等。
1.2 试验设计
试验样地主要植物为芦苇,伴生种有獐茅和白茅等。2010年5 月在试验区内设置9块10 m × 10 m 的施氮样地,进行3种处理:对照(N1,N 0 g/(hm2·a))、低氮 (N2,N 3 g/(hm2·a) ) 和高氮 (N3,N 6 g/(hm2·a)),每种处理 3次重复。当年 5月份进行施氮处理,将NH4NO3溶于水,用花洒均匀喷施。于2014年10月采集土壤样品,用直径为4 cm的土钻按混合采样法采集0~20 cm 土层,每个土样由5~8 个采集点的土壤混合而成,用四分法取适量于土袋中,迅速带回实验室。带回的土样去除植物和动物残体等杂质,混合均匀后一部分保存在4℃冰箱,用于分析土壤微生物群落功能多样性。另一部分风干,磨细,过 100目筛,用于测定土壤理化性质(表1)。
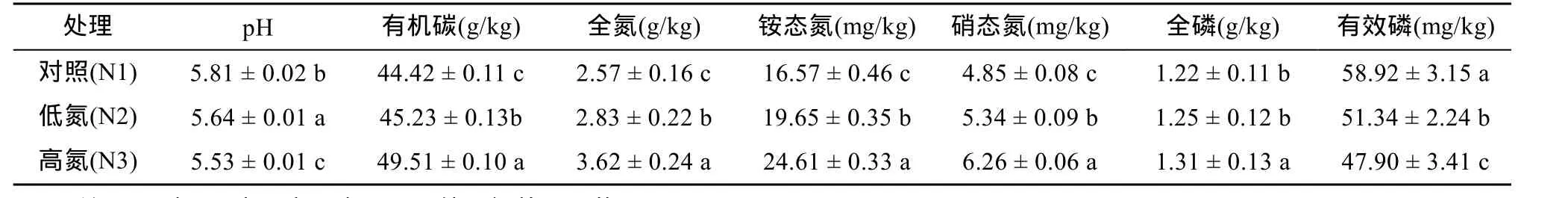
表1 模拟氮沉降条件下土壤理化性质的变化Table 1 Soil physical and chemical properties under different simulated nitrogen deposition levels
1.3 试验方法
本研究采用Biolog-ECO微平板法对不同氮沉降样地土壤微生物功能多样性进行分析。具体方法参照Classen等[11]。首先,土壤样品在25℃ 活化24 h,称取相当于10 g烘干土重的新鲜土样加入内有90 ml灭菌的生理盐水的三角瓶中,加无菌棉花塞后,将三角瓶在旋涡振荡器上震荡1 min,然后置于冰水浴中1 min,反复3次。静置2 min后按逐步稀释法,依次稀释 10-2和 10-3土壤悬浮液。用八通道移液器向ECO板每孔中加入10-3的土壤悬浮液,接种量为150µl,将接种好的微孔板放在(25±1)℃下连续培养13 d,每隔24 h用酶标仪测定590 nm处光密度值。
土壤pH测定的水土比为2.5∶1;土壤含水率采用烘干法测定;土壤有机碳利用碳氮分析仪测定(耶拿-2100S, 德国);土壤中全氮采用半微量凯氏定氮法测定;硝态氮采用酚二磺酸比色法测定;铵态氮采用氯化钾浸提-靛酚蓝比色法测定。
1.4 数据处理
微生物代谢强度采用平均颜色变化率 AWCD(average well color development)来描述,其是微生物利用单一碳源能力的重要指标。采用 Biolog微平板培养168 h的数据进行统计分析,此时即使生长较慢的微生物也已参与碳源的代谢过程,因此能较全面地反映微生物群落的信息。采用Shannon多样性指数、Simpson指数和 McIntosh指数来表征土壤微生物群落代谢功能多样性。
平均颜色变化率[14-16]:


式中:Ci为第i个碳源孔OD值;R为对照孔OD值,若Ci-R≤0,计为 0;ni是第i孔的相对吸光度值;Pi为ni与整个平板相对吸光值总和的比率。
采用SPSS 19.0软件进行主成分分析(PCA)和单因素方差分析(one-way ANOVA),用S-N-K法进行差异显著性检验,显著性水平设为 α=0.05;绘图采用Sigmaplot 12.5软件。
2 结果与分析
2.1 不同氮沉降条件下土壤微生物对碳源利用的动力学特征
从图1可以看出,培养期间AWCD随着培养时间的延长而升高,其中,24~168 h时AWCD值快速增加,说明微生物不断适应ECO板各孔的环境,处于对数生长期,碳源被大量利用,此时微生物代谢活性旺盛;随着培养时间的延长,168 h以后土壤微生物缓慢步入稳定期。比较不同氮沉降条件下土壤微生物群落 AWCD发现,高氮处理(N3)>低氮处理(N2)>对照(N1),说明随着土壤氮含量的增加AWCD值逐渐升高。
2.2 不同氮沉降条件下土壤微生物群落的多样性指数
土壤微生物群落对不同碳源的利用情况可以用多样性指数表示。表 2中显示了土壤微生物在培养168 h的Shannon、Simpson和McIntosh多样性指数,可在数量特征上相对反映土壤微生物群落物种组成和个体数量分布的情况。结果显示,在3个氮沉降湿地中,随着氮沉降的增加(表2),土壤微生物Shannon指数和 McIntosh指数均表现出升高的趋势。其中,高氮条件下的土壤微生物 Shannon指数和 McIntosh指数均最高,低氮次之,对照最低,各处理差异均达显著水平(P<0.05),而Simpson指数变化不显著。

图1 不同氮沉降条件下湿地土壤微生物的AWCDFig. 1 AWCD of soil microorganisms under different simulated nitrogen deposition levels

表2 不同氮沉降条件下湿地土壤微生物群落多样性指数Table 2 Diversity indexes of soil microbial communities under different simulated nitrogen deposition levels
2.3 不同氮沉降条件下土壤微生物群落对不同类型碳源的利用强度
Biolog-ECO板中含有31种碳源,包括碳水化合物类(10种)、氨基酸类(6种)、羧酸类(7种)、多聚物类(4种)、酚酸类(2种)和胺类(2种)。由图2可知,不同氮沉降湿地土壤微生物群落对不同类型碳源的利用强度明显不同,氮沉降浓度是影响不同碳源利用强度的重要因素,高氮样地(N3)土壤微生物对于碳水化合物类、氨基酸类和胺类的利用强度较大;低氮样地(N2)土壤微生物对碳水化合物类利用强度最大;对照样地(N1)土壤微生物对氨基酸类利用强度最大。整体来看,土壤微生物对碳水化合物类碳源利用最多,其次是羧酸类和氨基酸类,对酚酸类的利用则相对较少,可见氮沉降改变了土壤微生物对碳源类型的利用。
2.4 不同氮沉降条件下土壤微生物群落功能主成分分析

图2 不同氮沉降条件下土壤微生物群落对不同碳源的利用强度Fig. 2 Utilization intensities of different carbon sources by soil microbial communities under different simulated nitrogen deposition levels
对培养168 h的AWCD值进行微生物群落功能主成分分析,其中第一主成分(PC1)和第二主成分(PC2)的方差贡献率分别为62.18% 和9.82%,累积方差贡献率为72%,说明PC1和PC2是微生物群落碳源利用变异的主要来源,可以解释变异的绝大部分信息。从图3可以看出,不同氮沉降处理湿地土壤微生物群落在碳源利用上有明显的空间分异,而且在PC1和 PC2上的得分系数均差异显著,可见土壤微生物群落碳代谢特征受到不同氮沉降的显著影响。在PC2轴上,各样地土壤微生物碳源利用的主成分分析得分排序为:高氮处理>低氮处理>对照,与AWCD值和多样性指数的分析结果一致,总体呈现随氮浓度的增加碳源利用活性呈现上升的趋势。不同氮沉降条件下土壤微生物群落明显分为3簇。
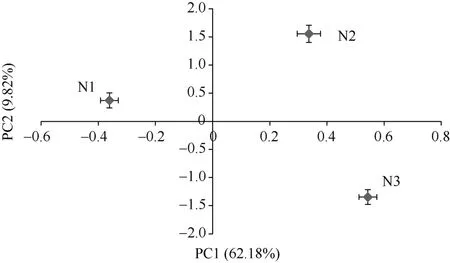
图3 不同氮沉降条件下湿地土壤微生物群落功能主成分分析Fig. 3 Principal component analyses of functions of soil microbial communities under different simulated nitrogen deposition levels
31种碳源在2个主成分上的载荷值见表3。从表 3可以看出与 PC1具有较高相关性的碳源有 13种,其中碳水化合物类6种、氨基酸类2种、羧酸类2种、多聚物类2种、酚酸类1种;与PC2具有较高相关性的碳源有9种,包括碳水类1种、氨基酸类4种、羧酸类3种、多聚物类1种、酚酸类1种。综合PC1和PC2的结果可知,不同氮沉降处理湿地中对土壤微生物群落代谢功能起作用的主要碳源类型为碳水化合物类、氨基酸类和羧酸类,不同氮沉降处理湿地的差异主要体现在对碳水化合物类、氨基酸类和羧酸类碳源的利用上,其中碳水化合物类最为突出。

表3 31种碳源与PC1、PC2的相关系数Table 3 Correlation coefficients between 31 carbon sources and PC1, PC2
2.5 不同氮沉降条件下土壤微生物代谢特性与土壤理化性质的相关性
将土壤有机碳、全氮、硝态氮、铵态氮含量及土壤含水率和pH与土壤微生物群落多样性指数进行相关分析,结果见表4,pH与微生物的代谢活性(AWCD)和 Shannon指数呈显著正相关关系;有机碳含量与Shannon指数和Simpson指数显著负相关,与AWCD显著正相关;全氮含量与Shannon指数显著负相关,而与McIntosh指数、AWCD显著正相关。
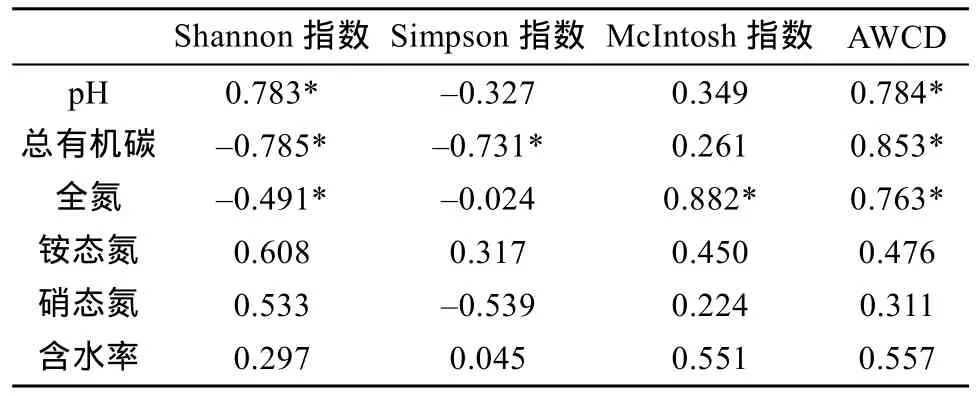
表4 不同氮沉降条件下湿地土壤微生物多样性指数与理化性质的相关性Table 4 Correlation coefficients between soil physiochemical properties and diversity indexes of soil microbial community under different simulated nitrogen decomposition levels
3 讨论
土壤微生物群落多样性是反映土壤肥力、土壤环境变化过程的重要指标之一,可以很好地揭示不同氮沉降条件影响下土壤微生物功能的差异,在生态系统功能中具有重要的意义。在本研究中,随着培养时间的延长 AWCD值逐渐上升,且随着对照-低氮-高氮湿地的变化 AWCD值呈现上升趋势,这与郁培义等[21]对不同氮添加条件下樟树林红壤土壤微生物群落多样性的研究规律一致。
Shannon指数、Simpson指数和McIntosh指数分别表征土壤中微生物群落的丰富度、优势度以及均匀度[6]。在 3个氮沉降条件下,除 Simpson指数外,Shannon指数和 McIntosh指数均差异显著。这说明土壤理化性质影响了微生物的群落结构组成,这与很多研究的结果一致[22-23]。
土壤微生物群落对不同类型碳源的利用强度存在一定的差异,在3个不同氮沉降处理中,土壤微生物均对碳水化合物类碳源的利用率最高,其为优势碳源,主要包括β-甲基-D-葡萄糖苷、D-半乳糖酸-γ-内脂、D-木糖、N-乙酰-D-葡萄糖胺、D-纤维二糖、α-D-乳糖;3个氮沉降处理土壤微生物功能多样性的差异主要体现在对碳水化合物类、氨基酸类和羧酸类碳源的利用上。这与朱凡等[22]等的研究结论是一致的。朱凡等[22]发现,不同施氮条件下土壤微生物对碳源的利用率有所不同,不同施氮条件下土壤微生物功能上的差别是由各微生物群落对碳源的总体利用率产生的差别。本研究发现氮沉降处理中土壤微生物群落对胺类碳源的利用效率也占到了20% 以上,而且对碳源的利用率具有很强的选择性,说明在氮沉降后土壤微生物的多样性发生了改变,对碳源的利用具有选择性;不同氮沉降条件下湿地土壤微生物对碳源的利用率比较均衡,这也与朱凡等[22]的结果一致。说明土壤微生物所表现出的对碳源利用的不一致性是受到土壤性质的影响,受到多因素的控制的。
土壤微生物对31种碳源的利用主成分分析结果显示,3个氮沉降条件下土壤微生物对碳源的利用特征差异比较明显,这一点可能和氮沉降进入土壤后土壤养分发生改变有一定的关系。从对照-低氮-高氮,随着施氮量的增加,土壤养分发生了变化,因而土壤微生物代谢特征发生一定的改变,使其对碳源利用的种类和强度均发生变化。郁培义等[21]和袁颖红等[23]也指出,地上的模拟氮沉降通过改变土壤中氮的含量对土壤微生物功能代谢产生重要影响。此外,通过对土壤微生物的多样性指数和AWCD值进行主成分分析可知,3个氮沉降处理明显出现分化,更为有力地证明了湿地条件下氮沉降的浓度变化会对土壤微生物的组成和功能活性产生重要的影响。
综上所述,不同氮沉降条件下湿地土壤微生物群落代谢特征具有较为明显的差异。不同氮沉降条件湿地由于其外源氮的大量输入,导致土壤理化性质发生变化,这些是影响土壤微生物代谢活性的主要因素。但由于 Biolog-Eco微平板技术的局限性,仅采用此技术很难完整地反映不同氮沉降条件下湿地土壤微生物群落功能多样性的变化规律。目前随着其他技术手段的快速发展,结合多种微生物学研究方法,如磷酸脂肪酸( phospholipid fatty acid,PLFA) 谱图分析法和分子生物学技术(如宏基因组技术),将为完整了解滨海湿地微生物群落结构和功能等信息提供很好的途径。
4 结论
综合来看,氮沉降的增加明显地改变了滨海湿地土壤微生物的功能活性,进而引起土壤微生物多样性和群落结构组成的变化。氮沉降的增加改变了土壤中有效元素组成,也改变了土壤微生物对不同碳源的利用模式,最终导致微生物功能活性发生改变。氮沉降的增加改变了湿地土壤中氮素浓度和形态,增加了土壤微生物的活性。所以,未来大气氮沉降的持续增加会对我国滨海湿地土壤微生物功能活性产生影响。
[1] Wang S, Fu B J, Gao G, et al. Responses of soil moisture in different land cover types to rainfall events in a re-vegetation catchment area of the Loess Plateau, China[J].Catena, 2013, 101(3): 122-128
[2] Nurulita Y, Adetutu E M, Kadali K K, et al. The assessment of the impact of oil palm and rubber plantations on the biotic and abiotic properties of tropical peat swamp soil in Indonesia[J]. International Journal of Agricultural Sustainability, 2015, 13(2): 150-166
[3] Papanikolaou N, Britton A J, Helliwell R C, et al. Nitrogen deposition, vegetation burning and climate warming act independently on microbial community structure and enzyme activity associated with decomposing litter in low-alpine heath[J]. Global Change Biology, 2010, 16(11):3120-3132
[4] Li J, Zhou X, Yan J, et al. Effects of regenerating vegetation on soil enzyme activity and microbial structure in reclaimed soils on a surface coal mine site[J]. Applied Soil Ecology, 2015, 87: 56-62
[5] 于树, 汪景宽, 李双异. 应用PLFA方法分析长期不同施肥处理对玉米地土壤微生物群落结构的影响[J]. 生态学报, 2008, 28(9): 4221-4227
[6] 王光华, 刘俊杰, 齐晓宁, 等. Biolog和PCR—DGGE技术解析施肥对德惠黑土细菌群落结构和功能的影响[J].生态学报, 2008, 28(1): 220-226
[7] 张海涵, 唐明, 陈辉, 等. 不同生态条件下油松(Pinus tabulaeformis)菌根根际土壤微生物群落[J]. 生态学报,2008, 27(12): 5463-5470
[8] 郝晓晖, 胡荣桂, 吴金水, 等. 长期施肥对稻田土壤有机氮、微生物生物量及功能多样性的影响[J]. 应用生态学报, 2010, 21(06): 1477-1484
[9] 范瑞英, 杨小燕, 王恩姮, 等. 黑土区不同林龄落叶松人工林土壤微生物群落功能多样性的对比研究[J]. 北京林业大学学报, 2013, 35(2): 63-68
[10] Dong D, Zhang D P, Liu W C, et al. Diversity Analysis of Bacterial Community from Permafrost Soil of Mo-he in China[J]. Indian Journal of Microbiology, 2014, 54(1):111-113
[11] Classen A T, Boyle S I, Haskins K E, et al.Community-level physiological profiles of bacteria and fungi: Plate type and incubation temperature influences on contrasting soils[J]. FEMS Microbiology Ecology, 2003,44(3): 319-328
[12] 林瑞余, 戎红, 周军建, 等. 苗期化感水稻对根际土壤微生物群落及其功能多样性的影响[J]. 生态学报, 2007,27(9): 3644-3654
[13] Xie X M, Liao M, Ma A L, et al. Effects of contamination of single and combined cadmium and mercury on the soil microbial community structural diversity and functional diversity[J]. Chinese Journal of Geochemistry, 2011, 30:366-374
[14] Liao M, Xie X M, Peng Y, et al. Characteristics of soil microbial community functional and structure diversity with coverage ofSolidago CanadensisL. [J]. Journal of Central South University, 2013, 20(3): 749-756
[15] Jin Z, Ji F Y, Xu X, et al. Microbial and metabolic characterization of a denitrifying phosphorus-uptake/side stream phosphorus removal system for treating domestic sewage[J]. Biodegradation, 2014, 25(6): 777-786
[16] Velasco A G, Probanza A, Mañero F J G, et al. Effect of fire and retardant on soil microbial activity and functional diversity in a Mediterranean pasture[J]. Geoderma. 2009,153(1): 186-193
[17] 范瑞英, 杨小燕, 王恩姮, 等. 未干扰黑土土壤微生物群落特征的季节变化[J]. 土壤, 2014, 46(2): 285-289
[18] 岳冰冰, 李鑫, 张会慧, 等. 连作对黑龙江烤烟土壤微生物功能多样性的影响[J]. 土壤, 2013, 45(1): 116-119
[19] 江玉梅, 谢晶, 曹广泮, 等. 江西退化红壤人工重建森林土壤微生物碳源代谢功能研究[J]. 土壤学报, 2014(1):158-165
[20] 王秀国, 王一奇, 严虎, 等. 多菌灵重复施药对其持久性及土壤微生物群落功能多样性的影响[J]. 土壤学报,2010, 47(1): 131-137
[21] 郁培义, 朱凡, 王志勇, 等. 氮添加对樟树林红壤微生物群落代谢功能的影响[J]. 中南林业科技大学学报,2013, 33(3): 70-74
[22] 朱凡, 李天平, 郁培义, 等. 施氮对樟树林土壤微生物碳源代谢的影响[J]. 林业科学, 2014, 8(8): 82-89
[23] 袁颖红, 樊后保, 李辉信, 等. 模拟氮沉降对杉木人工林土壤微生物的影响[J]. 林业科学, 2012, 9(9):8-14
猜你喜欢
当代水产(2022年8期)2022-09-20
环境工程技术学报(2022年3期)2022-06-05
昆明医科大学学报(2022年2期)2022-03-29
中国土壤与肥料(2021年5期)2021-12-02
数学小灵通(1-2年级)(2021年10期)2021-11-05
食品安全导刊(2021年20期)2021-08-30
昆钢科技(2021年6期)2021-03-09
数学小灵通(1-2年级)(2020年12期)2021-01-14
河南科学(2020年3期)2020-06-02
天津科技(2020年4期)2020-05-09
