
油杉人工林凋落物及其碳氮归还动态
2018-01-12 01:02:14陈金章
福建林业科技 2017年4期
陈金章
(泉州市林业局,福建 泉州 362000)
油杉(Keteleeriafortunei)为松科大乔木,喜光树种,其根系粗壮、发达,对土壤的适应性较广,适宜干旱环境。其树干端直,木材黄褐色,材质重,纹理直,耐水湿,抗腐性强,是珍贵用材树种;其树形优雅美观,枝叶茂密浓绿,具有很高的观赏价值,是非常优良的园林绿化树种[1-3]。目前,对油杉的研究主要集中在生态学和生物学特性[4-9],以及苗木培育和造林等方面[10-16],未见油杉凋落物方面的报道。凋落物是人工林养分归还林地的主要途径,植物在生长过程中,凋落物所含的营养元素经分解释放后归还土壤,并在表层富集,供植物吸收利用,凋落物中各种营养元素对土壤肥力具有重要作用[17]。本文对油杉人工林凋落物量收集养分测定,研究油杉人工林碳氮归还动态规律,为油杉人工林的科学经营提供参考。
1 材料与方法
1.1 试验地概况
试验地位于福建省永春县桃城镇大坪村湖内,海拔490~590 m,地理坐标为东经118°17′44″、北纬25°21′05″。土壤为山地红壤,土层深厚,立地质量等级为Ⅱ级,林下植被主要为亚热带常绿阔叶树种和蕨类植物。该地属南亚热带季风气候区,气候温暖湿润,雨量充沛,年均降水量1676.3 mm,极端最高温39.2 ℃,极端最低温-3.2 ℃,年均气温20.4 ℃,≥10 ℃的年均积温6984 ℃,年无霜期310 d[1-3]。
1.2 材料来源
试验林为油杉人工林群落,面积约2 hm2。2014年8月,在试验山场设立3个有代表性样地,样地面积为400 m2(20 m×20 m),样地内油杉的平均胸径为27.6 cm,平均树高为22.6 m,林龄为50 a,林分平均密度1683株·hm-2。
1.3 研究方法
1.3.1 凋落物收集与处理 在每个样地内分别设置3个规格为1 m×1 m的收集网,于2014年8月—2015年8月,每月月底收集1次凋落物,将每个收集网内的凋落物按枝、叶、杂(主要为球果以及林分中阔叶树和灌木的凋落物等)分类烘干并测定生物量,凋落物烘干后全部粉碎过40目筛,并称重取样。
1.3.2 凋落物碳氮元素的测定 将粉碎后的样品采用德国Elementar Vario MAX CNS元素分析仪,对油杉凋落物碳、氮元素的含量进行测定。
2 结果与分析
2.1 凋落物量的月动态

图1 凋落物量的月动态
油杉人工林凋落物的月动态,不但受其生物学特性的制约,而且还受所在地的气候环境的影响。从图1可以看出,油杉人工林月凋落物量有着明显的季节变化。2014年8月是全年较热且多雨时期,受台风雨活动的影响,凋落物量较多,这个时期凋落物主要是风吹雨打等机械原因造成的非生理性落枝落叶;而过了9月以后凋落物量有所增加,主要是因为这个时期生长基本结束,过渡到生长迟缓期,油杉受生物学特性影响,开始大量集中换叶和落叶;10月,油杉的月凋落物量出现了最大值,为904.83 kg·hm-2;进入11月后,油杉生理上的换叶量逐渐减少,再加上冬季雨量较小,油杉的月凋落物量在2015年1月达到最小值,为174.82 kg·hm-2;2015年1月后,受到春季换叶和降雨增多的影响,油杉的月凋落物量逐渐增加;2015年5月,油杉的月凋落物量再次出现1次高峰,为718.57 kg·hm-2;2015年5月以后,随着春季换叶和雨季的结束,油杉的月凋落物量逐渐减少。从各组分的月动态变化可以看出,凋落叶是凋落物的主要成分,各组分的动态变化与总量一致。
2.2 凋落物碳、氮含量的月动态
2.2.1 凋落物碳含量的月动态 由表1可以看出,凋落物碳含量大小排序为叶>枝>杂,凋落物碳平均含量变化较小,最大值出现在2014年9月,达498.817 g·kg-1;最小值出现在2015年7月,达469.306 g·kg-1。凋落枝和凋落叶碳含量变化较小,最大值出现在2014年9月,分别为501.468、509.836 g·kg-1;最小值出现在2015年7月,达471.608、483.332 g·kg-1。凋落杂物的碳含量变化较大,最大值出现在2015年2月,达448.381 g·kg-1;最小值出现在2015年3月,达390.407 g·kg-1。主要是因为凋落杂物的组成在不同的时间有所不同。
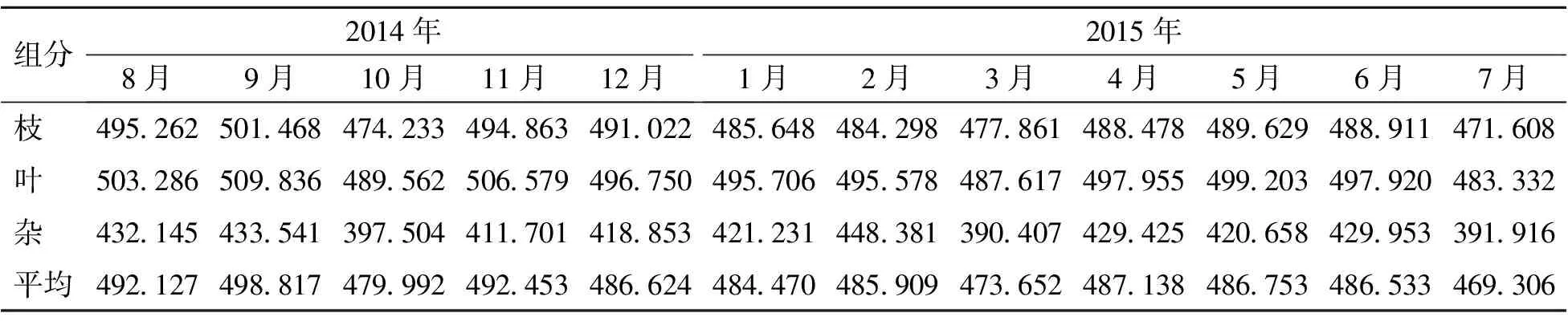
表1 凋落物碳含量的月动态 g·kg-1
2.2.2 凋落物氮含量的月动态 凋落物养分含量的高低可以反映植物对养分的利用效率,植物对养分利用效率高的,养分通常通过转移机制得到再利用。因此,凋落物养分含量的动态变化可以反映养分的转移状况[18]。油杉在生长过程中,开花和结果对养分消耗较大,造成凋落物养分含量较低。从表2可以看出,2014年8—11月,油杉处于结果的过程中,植株内养分消耗较大,凋落物氮平均含量逐渐下降,在2014年11月达到最低,为10.418 g·kg-1;11月以后,随着结果的结束,油杉植株内的养分消耗量逐渐下降,植株内的营养水平逐渐恢复,凋落物氮平均含量在2015年2月达到较大值;2015年2—3月,油杉开始开花,植株内养分消耗较大,凋落物氮平均含量逐渐下降;2015年5月,随着油杉花期结束和气温升高,植株内的营养水平迅速提高,凋落物氮平均含量快速升高达到最大值,为15.758 g·kg-1;2015年6月开始,随着新的一轮结果开始,植株内养分消耗逐渐增大,凋落物氮平均含量逐渐下降。凋落物各组分氮含量的月动态趋势与氮平均含量基本一致。
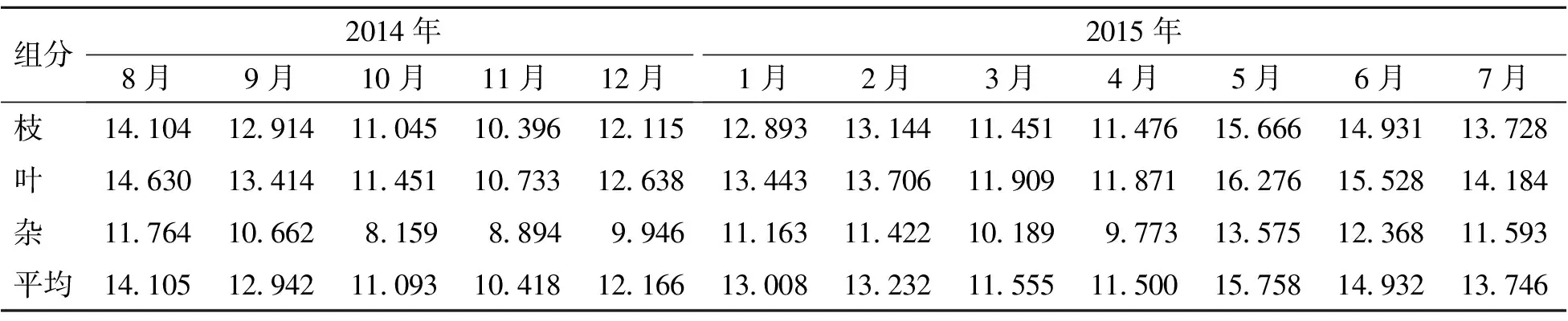
表2 凋落物氮含量的月动态 g·kg-1
2.3 凋落物碳、氮归还量的月动态
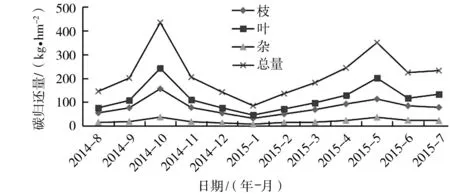
图2 凋落物碳归还量的月动态
2.3.1 凋落物碳归还量的月动态 从图2可以看出,凋落物碳归还量的月动态变化趋势与凋落物量的月动态变化趋势相似,凋落物碳归还量在全年呈现出2个峰值期,分别出现在2014年10月、2015年5月,碳总归还量分别达434.313、349.764 kg·hm-2,2个月的碳归还量之和占全年总归还量的30.41%,与凋落物总量的最高峰出现的月份相同;凋落物碳归还量2个低值则出现在2014年8月、2015年1月,碳总归还量低值分别只有145.415、84.693 kg·hm-2,2个月的碳归还量之和只占全年总归还量的8.92%。最高值是最低值的3.41倍,相差很大。

图3 凋落物氮归还量的月动态
2.3.2 凋落物氮归还量的月动态 从图3可以看出,凋落物氮归还量的月动态变化趋势与凋落物量的月动态变化趋势相似,凋落物氮归还量在全年呈现出2个峰值期,分别出现在2014年10月、2015年5月,氮总归还量分别达到10.038、11.323 kg·hm-2,2个月的氮归还量之和占总归还量的31.21%,与凋落物总量的最高峰出现的月份相同;凋落物氮归还量2个低值则出现在2014年8月、2015年1月,氮总归还量低值分别只有4.168、2.274 kg·hm-2,2个月的氮归还量之和只占全年的9.41%。最高值是最低值的3.32倍,相差很大。
3 小结与讨论
林木通过新陈代谢每年归还一定数量的枯落物,向林地输入相当数量的养分,为土壤动物、微生物提供食物来源。凋落物数量主要与样地的生产力状况相关,其它诸如空气温度、土壤水分和养分的可利用性亦限制着凋落物的数量[19]。油杉人工林凋落物量主要受油杉生理活动的季节变化和所在地气候环境的影响,油杉人工林月凋落物量有着明显的季节变化,呈双峰形态,这与大多数学者的研究结果相似[20]。2014年10月,受生物学特性影响,油杉开始大量集中换叶和落叶,油杉凋落物量出现第1次高峰。2015年5月,气温逐渐回暖,降雨增多,新叶旺盛生长,衰老叶子开始脱落[21],油杉凋落物量出现第2次高峰。
从凋落物养分含量月动态看,碳元素含量月动态比较稳定,碳元素平均含量介于469.306~498.817 g·kg-1之间;而氮元素含量受油杉生理活动的影响月动态变化较大,氮元素平均含量最大值是最小值的1.51倍。凋落物的碳、氮养分归还量月变化呈现出凋落物量及其元素含量月动态相叠加的规律。
油杉人工林碳、氮元素通过凋落物归还土壤,碳、氮元素年归还量分别为2578.325、68.434 kg·hm-2,年归还的碳、氮比为37.676。落叶是养分归还的主要组分,碳、氮元素年归还量分别为1402.528、37.643 kg·hm-2,分别占总量的54.40%、55.01%;落枝碳、氮元素年归还量分别为934.926、24.550 kg·hm-2,分别占总量的36.26%、35.87%;其它杂物凋落物碳、氮元素年归还量分别为240.872、6.240 kg·hm-2,分别占总量的9.34%、9.12%。
综上所述,凋落物的养分含量和养分归还量的月动态变化在森林树木生长和植被生态系统的养分循环研究中具有重要的生态学意义。目前,马尾松的松材线虫病害严重,而同为松科的油杉则具有木材好、生长较快、抗病害性强和抗瘠薄等优点。油杉作为具有巨大发展潜力的树种之一,对其林分的凋落物及养分归还量的长期定位监测研究具有十分重要的意义。
*:本研究得到国家林业局南方山地用材林培育重点实验室和福建省森林培育与林产品加工利用重点实验室的资助,福建省林业科学研究院肖祥希、高楠、何文广给予指导和帮助,在此一并致谢!
[1]高楠,肖祥希,何文广,等.50年生油杉人工林生长规律研究[J].广西林业科学,2015,44(3):219-224.
[2]高楠,肖祥希,何文广,等.油杉人工林生态系统养分分配格局[J].广西林业科学,2015,44(4):352-357.
[3]王丽琴.福建永春油杉人工林生物量结构特征[J].亚热带植物科学,2015,44(3):228-230.
[4]左索哺.中国油杉属分布型与植物区系分区关系的模糊分析[J].中南林学院学报,1989,11(2):199-205.
[5]吴文珊,张清其,刘剑秋.油杉染色体核型的研究[J].福建师范大学学报:自然科学版,1993,9(1):74-77.
[6]李国平,黄群策,秦广雍.油杉花粉个体发育中淀粉粒消长变化研究[J].林业科学研究,2005,18(4):416-420.
[7]李国平,黄群策.油杉花粉个体发育与传粉过程[J].林业科学,2006,42(5):42-48.
[8]李国平,黄群策,秦广雍.油杉花粉发育进程的共聚焦显微镜观察[J].北京林业大学学报,2006,28(6):151-154.
[9]牟凤娟,戴兴芬,李双智,等.油杉属植物研究动态[J].西部林业科学,2012,12(6):92-99.
[10]卢立光,彭桂华.油杉引种栽培试验初报[J].广东林业科技,1992(3):32-34.
[11]朱积余,韦增健,丘小军.油杉属树种人工造林的试验研究[J].林业科学,1993(1):67-71.
[12]韦增健,丘小军.油杉采种育苗试验研究[J].广西林业科技,1993,22(1):17-20.
[13]蔡卫兵,缪美琴.油杉容器育苗技术的研究[J].南京林业大学学报,1997,21(1):22-26.
[14]张纪卯.油杉不同苗木造林试验[J].福建林学院学报,1999(19):73-76.
[15]吴幼媚.油杉物种苗期比较试验初报[J].广西林业科学,2003(3):36-38.
[16]张纪卯.不同基质和容器规格对油杉容器苗生长的影响[J].福建林学院学报,2001,21(2):176-180.
[17]林成芳,李振,牛志鹏,等.福建柏人工林凋落物的养分动态特征[J].福建农林大学:自然科学版,2005,34(1):63-66.
[18]林宇,王雪梅,张勇,等.滨海沙地尾巨桉人工林凋落物及碳氮养分归还[J].西南林业大学学报,2013,33(5):24-28.
[19]潘辉,黄石德,洪伟,等.3种相思人工林凋落物量及其碳归还动态[J].福建林学院学报,2010,30(2):104-108.
[20]郭剑芬,陈光水,钱伟,等.万木林自然保护区2种天然林及杉木人工林凋落量及养分归还[J].生态学报,2006,26(12):4091-4098.
[21]刘东霞.万木林主要群落凋落物的动态研究[D].福州:福建农林大学林学院,2004:12-17.
猜你喜欢
地下水(2023年5期)2023-10-13 06:03:28
西部林业科学(2023年4期)2023-09-02 07:23:28
现代园艺(2017年19期)2018-01-19 02:50:08
现代园艺(2017年21期)2018-01-03 06:42:24
植物研究(2017年5期)2017-11-10 01:47:20
山东林业科技(2016年5期)2016-07-05 00:43:04
广西林业科学(2016年3期)2016-03-16 05:43:42
广西林业科学(2016年3期)2016-03-16 05:43:40
广西林业科学(2016年3期)2016-03-16 05:43:30
广西林业科学(2016年3期)2016-03-16 05:43:22
