
枯草芽孢杆菌M4诱导猕猴桃抗病相关的防御酶系研究
2018-01-06 01:06任平阮祥稳秦涛
江苏农业科学 2017年22期
任平+阮祥稳+秦涛
摘要: 采用比色法测定枯草芽孢杆菌M4发酵液不同组分诱导猕猴桃叶片中苯丙氨酸解氨酶(PAL)、过氧化物酶(POD)、多酚氧化酶(PPO)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)的活性变化。结果表明,接种M4发酵液、菌体悬液、上清液、LB培养基,猕猴桃叶片内PAL、POD、PPO、SOD、CAT活性均高于对照无菌水,PAL、PPO、SOD的活性峰值多在接种后8 d出现,POD、CAT的活性峰值多在接种后12 d出现;接种M4发酵液、菌体悬液的PAL、POD、PPO、SOD、CAT活性均明显高于接种上清液、LB培养基,且酶活变化趋势相似,说明M4发酵液中主要有效诱导组分为菌体。
关键词: 枯草芽孢杆菌;猕猴桃;多酚氧化酶;苯丙氨酸解氨酶;过氧化氢酶;超氧化物歧化酶;过氧化物酶
中图分类号:S436.634 文献标志码: A
文章编号:1002-1302(2017)22-0111-03
由丁香假单胞菌猕猴桃致病变种(Pseudomonas syringae pv. actinidiae,Psa)引起的猕猴桃细菌性溃疡病往往发病严重,流行迅速,防治相当困难。近年来,随着猕猴桃栽培面积的不断增加,细菌性溃疡病快速蔓延,自2005年以来,已在世界主要猕猴桃产区造成严重危害,成为猕猴桃栽培过程中最具毁灭性的病害 。
生物防治是控制植物病害、减少化学农药污染的有效途径,而诱导抗性又是植物病害生物防治的重要机理之一[3],一些微生物可作为植物激活剂,诱导植物产生系统抗性而具有良好的防病效果[4]。植物的抗病性与木质素及相关苯丙烷类代谢物质的积累、病程相关蛋白(PRP)的表达、磷酸戊糖途径、乙醛酸循环等生理过程密切相关[5-6],参与植物抗病代谢的关键酶有苯丙氨酸解氨酶(PAL)、过氧化物酶(POD)、多酚氧化酶(PPO)等,其活性变化是反映植物抗病能力强弱较直接和有效的指标之一。
近年来研究发现,芽孢杆菌(Bacillus ssp.)是生防菌的优势菌种,诱导植物抗性是其生防作用实现的重要机制之一[7-8]。陕西省科学院酶工程研究所微生物实验室保藏的枯草芽孢杆菌M4对猕猴桃溃疡病具有较好的生防效果,但该菌株是否具有诱导抗性作用,有关其不同组分对猕猴桃抗病相关酶活影响的研究国内外还未见报道。因此,本研究选择PPO、POD、PAL、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)这5种酶作為猕猴桃抗病性反应指标,研究M4菌株发酵液不同组分对猕猴桃叶片内抗病相关防御酶系的影响,以期揭示其诱导抗病性机理,为指导其田间应用提供理论依据。
1 材料与方法
1.1 试验材料
枯草芽孢杆菌M4,陕西省科学院酶工程研究所微生物实验室保藏;试验植株为2年生盆栽秦美猕猴桃。LB培养基:胰蛋白胨10 g,酵母浸膏5 g,氯化钠5 g,蒸馏水1 000 mL,pH值为7.0~7.2。
1.2 菌液的制备及接种
将菌株M4接种于营养肉汤培养基中,37 ℃、160 r/min振荡培养24 h;将发酵液分成3份,1份保留,另2份合并,4 ℃ 10 000 r/min离心30 min;分别收集菌体和上清液;菌体用无菌水漂洗,再次离心收集,用无菌水稀释成108 CFU/mL的菌液,而上清液用孔径为0.22 μm的细菌过滤器过滤,待温度升至室温,备用。试验设108 CFU/mL发酵液(F-0)、108 CFU/mL 菌体悬液(F-1)、无菌发酵上清液(F-2)3个处理,分别以无菌水、LB培养基处理为对照1(CK1)、对照2(CK2),选取长势一致、健壮的秦美猕猴桃植株,采用叶面喷施接种法进行诱导抗性试验,将菌液均匀喷在猕猴桃叶片上。
1.3 PAL、POD、PPO、SOD、CAT活性的测定
接种后0、4、8、12、16、20 d分别采样,测定猕猴桃叶片PAL、POD 、PPO、SOD、CAT的活性。PAL提取及活性测定参照薛应龙等的方法[9]进行,以1 g鲜质量D290 nm值变化0.01所需酶量为1个酶活力单位(U)。POD提取及活性测定参照张龙翔等的方法[10]进行,以1 g鲜质量1 min内D470 nm值变化0.01作为1个酶活力单位(U)。PPO提取及活性测定参照朱广廉等的方法[11]进行,以1 g鲜质量1 min内D525 nm值变化0.01为1个酶活力单位(U)。SOD提取及活性测定参照朱广廉等的方法[11]进行,以抑制氮蓝四唑(NBT)还原的50%酶量为1个酶活力单位,以不加酶液而用缓冲液代替酶液的处理为空白对照,酶活计算公式为:
N=2×(D560 nm空白-D560 nm处理)/D560 nm空白。
CAT提取及活性测定参照李合生的方法[12]进行,采用紫外吸收法,以1 g鲜质量1 min内D240 nm减少0.1的酶量为1个酶活单位(U)。
2 结果与分析
2.1 M4菌株对苯丙氨酸解氨酶(PAL)活性的影响
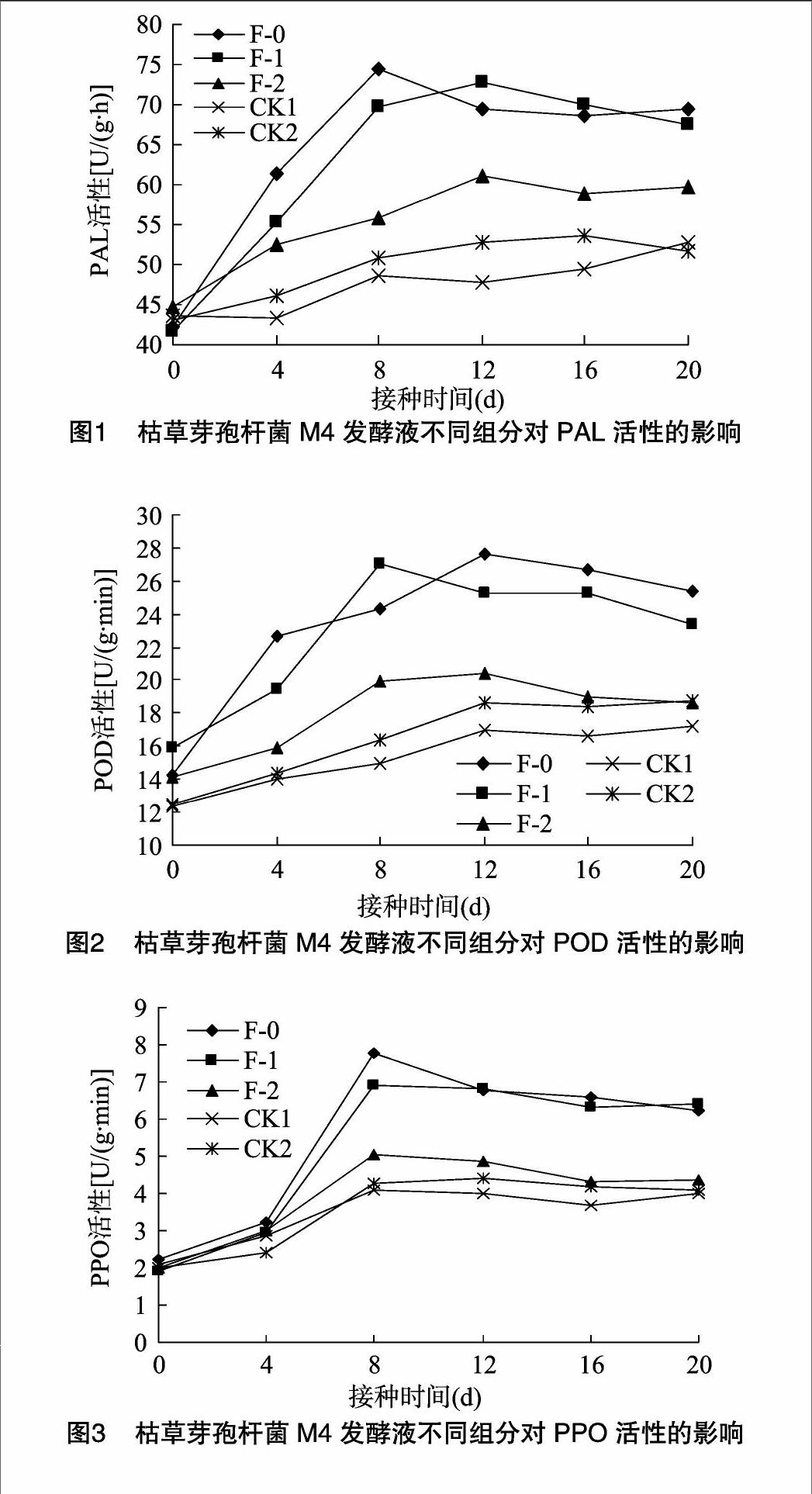
由图1可知,猕猴桃叶片接种发酵液不同组分,其PAL活性均高于对照无菌水(CK1);菌株M4发酵液(F-0)接种后8 d,猕猴桃叶片内PAL活性值达到最高峰,为 74.42 U/(g·h),比CK1提高53.0%;菌体悬液(F-1)接种后12 d,猕猴桃叶片内PAL活性相对较高,为 72.65 U/(g·h),比CK1提高了51.7%,后缓慢下降;接种后20 d,接种M4发酵液、菌体悬液、上清液的猕猴桃叶片PAL活性仍高于CK1、CK2;接种后4~20 d,发酵液、菌体悬液处理的PAL活性总体高于上清液、无菌水、LB培养基;接种M4发酵液、菌体悬液处理的PAL活性变化趋势相似,接种上清液、LB培养基处理的PAL活性及其变化趋势相似。菌株M4发酵液和菌体悬液处理能明显提高猕猴桃植株内PAL活性。
2.2 M4菌株对过氧化物酶(POD)活性的影响
由图2可知,与CK1相比,接种M4发酵液(F-0)、菌体悬液(F-1)、上清液(F-2)、LB培养基(CK2)的猕猴桃叶片,在接种后0~12 d内能明显提高叶片内POD活性;菌体悬液处理的猕猴桃叶片POD活性在接种后8 d达到最大值,为27.02 U/(g·min),M4发酵液处理的猕猴桃叶片POD活性在接种后12 d达到峰值,为27.67 U/(g·min),后呈下降趋
势;接种M4发酵液、菌体悬液、上清液后12 d,猕猴桃叶片POD活性分别比CK1提高62.4%、48.6%、19.7%;接种M4发酵液、菌体悬液后4~20 d,猕猴桃叶片POD活性总体高于上清液、无菌水、LB培养基,上清液和LB培养基处理的POD活性变化趋势相似。菌株M4发酵液和菌体悬液处理能明显提高猕猴桃植株内POD活性。
2.3 M4菌株对多酚氧化酶(PPO)活性的影响
由图3可知,接种M4发酵液(F-0)和菌体悬液(F-1)后0~8 d,猕猴桃叶片内的PPO活性有明显提高,并在接种后8 d达到最高峰,PPO活性分别为7.79、6.93 U/(g·min),比CK1分别提高90.9%、69.9%;接种后8~12 d,叶片内的PPO活性开始下降,接种后12~20 d内的猕猴桃叶片PPO活性变化差异不明显;接种后4~20 d,M4发酵液、菌体悬液处理的PPO活性总体上高于清液、无菌水、LB培养基,上清液与LB培养基处理的PPO活性变化趋势相似。M4发酵液和菌体悬液处理能明显提高植株内PPO活性。
2.4 M4菌株对超氧化物歧化酶(SOD)活性的影响
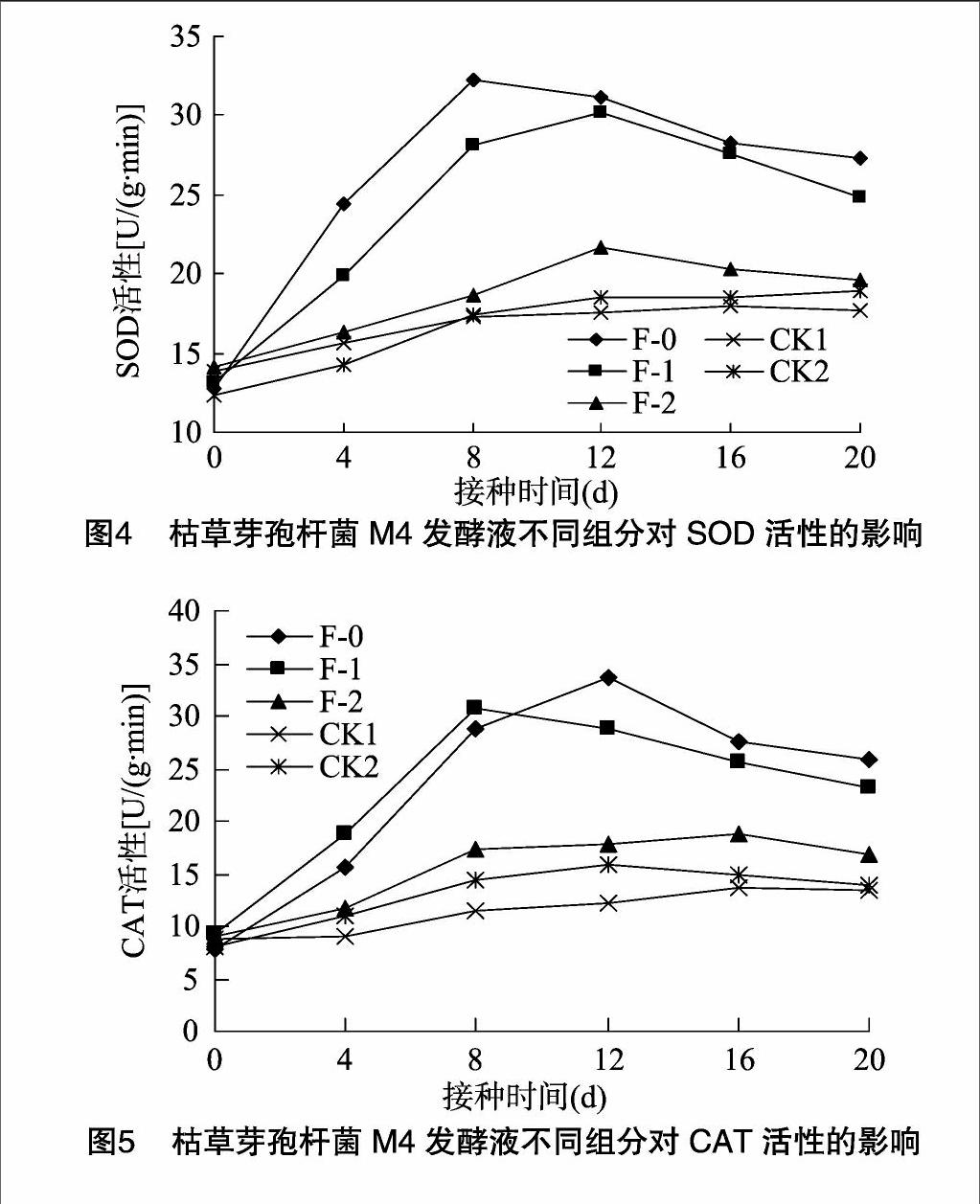
由图4可知,接种发酵液不同组分后4~8 d,猕猴桃叶片内的SOD活性呈增加趋势;接种发酵液(F-0)后8 d,猕猴桃叶片内的SOD活性达最大值,为32.22 U/(g·min),比CK1提高86.1%;接种菌体悬液(F-1)后12 d,猕猴桃叶片内的SOD活性达最大值,为 30.21 U/(g·min),比CK1提高 72.3%;接种后4~20 d,接种M4发酵液的叶片SOD活性高于其他处理,M4发酵液、菌体悬液处理的SOD活性总体高于上清液、无菌水、LB培养基。M4发酵液和菌体悬液处理能明显提高植株内SOD活性。
2.5 M4菌株过氧化氢酶(CAT)活性的影响
由图5可知,接种发酵液不同组分后4~8 d,猕猴桃叶片内的CAT活性呈增加趋势,其中接种菌体悬液(F-1)的猕猴桃叶片CAT活性高于其他处理,且再接种后8 d CAT活性达最大值,为30.64 U/(g·min);接种M4发酵液(F-0)后 12 d 猕猴桃叶片的CAT活性达最大值,为33.78 U/(g·min), 比CK1提高176.7%;接种后4~20 d,M4发酵液、菌体悬液处理的CAT活性总体高于上清液、无菌水、LB培养基,菌体、上清液处理的CAT活性变化趋势相似。M4发酵液和菌体悬液处理能明显提高植株内CAT活性。
3 结论与讨论
研究枯草芽孢杆菌M4发酵液、菌体悬液、上清液、LB培养基处理后猕猴桃叶片内的苯丙氨酸解氨酶(PAL)、过氧化物酶(POD)、多酚氧化酶(PPO)、超氧化物歧化酶(SOD)、過氧化氢酶(CAT)的活性变化发现,与清水处理(CK1)相比,猕猴桃叶片内这5种酶的活性有明显提高,并在达到峰值后呈下降趋势;PAL、PPO、SOD的活性峰值多在接种后8 d出现,POD、CAT的活性峰值多在接种后12 d出现。比较不同组分接种处理诱导产生的酶活性强弱发现,接种M4发酵液、菌体悬液的5种酶活性均明显高于接种上清液、LB培养基,且酶活变化趋势相似,说明M4发酵液中主要有效诱导组分为菌体。
植物病害生物防治机制包括抗生作用、溶菌作用、重寄生作用、竞争作用、交互保护作用及促生作用,诱导系统抗性也被认为是生防菌的一种重要防病机制。植物的诱导抗病包括木质化、防御酶活性、病程相关蛋白、植保素等多种生理生化因子的合成。有大量研究表明,参与植物体内多种防卫反应的PAL、PPO、POD等酶系与植物抗病性密切相关[13],这些酶的活性与植株抗病性呈正相关,在植物的抗病反应中起到非常重要的作用。猕猴桃植株接种M4发酵液和菌体悬液,防御酶活性升高,说明菌株M4诱导的系统抗性可使植株内酶活性增强,这可能是其生物防治作用的机制之一。
枯草芽孢杆菌M4发酵液不仅具有抑菌作用,还可以通过诱导植物防御酶活性的提高,从而增强植株的抗病性,这些防御酶在拮抗猕猴桃溃疡病菌的过程中可能激活了猕猴桃本身的抗病代谢过程,使猕猴桃植株体内发生一系列的生理生化反应,产生相关抗性物质,诱导抗病效果明显,但是,植株体内产生何种抵抗物质尚未明确,有待进一步研究。
参考文献:
[1] 李 淼,檀根甲,李 瑶,等. 不同抗性猕猴桃品种感染溃疡病前后几种保护酶活性变化[J]. 激光生物学报,2009,18(3):370-378.
[2]关 侠,高小宁,黄丽丽. 陕西省猕猴桃细菌性溃疡病的发生及防治[J]. 陕西农业科学,2010,56(1):78-79.
[3]谭可菲,段玉玺,朱晓峰,等. 根结线虫生防菌snef8对番茄诱导抗性的初步研究[J]. 黑龙江农业科学,2011(1):22-24.
[4]令利军,冯 蕾,雷 蕾,等. 地衣芽孢杆菌TG116诱导黄瓜抗病性相关防御酶系的研究[J]. 西北师范大学学报(自然科学版),2016,52(1):100-104.
[5]Dong H Z,Li W J,Zhang D M,et al. Differential expression of induced resistance by an aqueous extract of killed Penicillium chrysogenum against Verticillium wilt of cotton[J]. Crop Protection,2003,22(1):129-134.
[6]Koike N,Hyakumachi M,Kageyama K,et al. Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi:lignification and superoxide generation[J]. European Journal of Plant Pathology,2001,107(5):523-533.
[7]齐爱勇,赵绪生,刘大群. 芽孢杆菌生物防治植物病害研究现状[J]. 中国农学通报,2011,27(12):277.
[8]朱明妍,刘 皎,杜春梅. 芽孢杆菌生物防治植物病害研究进展[J]. 安徽农业科学,2013,40(34):16635.
[9]薛应龙,欧阳光察,澳绍根. 植物苯丙氨酸解氨酶的研究Ⅳ.水稻幼苗中PAL活性的动态变化[J]. 植物生理学报,1983,9(3):301-305.
[10] 张龙翔,张庭芳,李令媛. 生化实验方法和技术[M]. 2版.北京:高等教育出版社,1997.
[11]朱广廉,钟海文,张海琴. 植物生理学实验[M]. 北京:北京大学出版社,1990.
[12]李合生. 植物生理生化试验原理和技术[M]. 北京:高等教育出版社,2000:32-33.
[13]林陈强,李占飞,张 慧,等. 枯草芽孢杆菌CS16诱导香蕉抗病性相关防御酶系的研究[J]. 福建农业学报,2013,28(6):570.
猜你喜欢
快乐语文(2021年36期)2022-01-18
动漫星空(兴趣百科)(2020年12期)2020-12-12
作文小学中年级(2019年9期)2019-10-14
热带农业科学(2016年11期)2017-01-21
创新作文(小学版)(2016年23期)2016-12-01
中国实用医药(2016年2期)2016-01-05
分析化学(2015年8期)2015-08-13
中国当代医药(2015年18期)2015-08-06
