
温度和激素对不同种源连香树种子发芽的影响
2018-01-06 08:51陈学龙丁全定付作霖
四川林业科技 2017年6期
陈学龙,王 飞*,丁全定,付作霖,齐 昊
(1.甘肃省白龙江林业管理局林业科学研究所,甘肃 兰州 730070;2.甘肃白龙江森林生态系统国家定位观测研究站,甘肃 舟曲 746300)
温度和激素对不同种源连香树种子发芽的影响
陈学龙1,2,王 飞1,2*,丁全定1,2,付作霖1,2,齐 昊1,2
(1.甘肃省白龙江林业管理局林业科学研究所,甘肃 兰州 730070;2.甘肃白龙江森林生态系统国家定位观测研究站,甘肃 舟曲 746300)
为研究不同温度和激素对白龙江不同种源地连香树种子发芽的影响,本研究采用2种外源激素GA和PEG,在不同温度梯度下的连香树种子发芽实验表明:(1)GA更有利于连香树种子发芽。(2)舟曲县连香树种子活性优于宕昌县种子。(3)在同一温度下随着激素浓度增大,发芽率表现出先增后减的趋势;随着温度的增加同一浓度种子发芽率表现出增大的趋势;同一温度下发芽指数和发芽势变化一致。(4)随着激素浓度和温度的增加平均发芽时间均减小,及高浓度、高温有利于缩短种子发芽时间。
激素;温度;连香树;种子;发芽
连香树(CercidiphyllumjaponicumSieb.et Zucc.),系连香树科(Cercidiphyllaceae)连香树属(CercidiphyllumSieb. et Zucc. )落叶大乔木,属第三纪孑遗植物,是东亚特有的珍稀濒危树种[1],被列为我国二级保护植物[2]。连香树起源古老,具有较高的科研和利用价值,而且具有香料、造纸、药用和木材等多种经济效益,以及绿化、美化、优化环境等景观效果[3~4]。该种目前分布于我国的川、甘、晋、陕、鄂、湘、赣、皖、豫、浙等多省,但种群明显斑块化,且种群规模极其有限,多数分布地甚至呈单株分布[2]。该树种以有性繁殖和无性繁殖,有性繁殖主要以种子为主;种子发芽状况也是判定种子质量、确定播种量的一项重要指标,也是种群自然更新的基础,对连香树种群保护和恢复具有重要意义;无性繁殖主要以扦插为主。
白龙江流域是甘肃连香树种群重要生长地之一,该区种群分布明显板块化,分布规模不大,多数呈现单株分布,且因放牧或人为砍伐,该种群数量减少,且无明显主干,多以簇状生长,树高多10 m以上。
1 材料与方法
1.1 材料采集
本文所用连香树种子于2016年10月在甘肃省武都区和舟曲县两地采集,将同一地区不同株树收集的种子混合,然后将采集的种子带回实验室自然风干,装入报纸后放入通风、干燥、遮光处保存,实验于2017年3月进行种子萌发实验,不同种源地基本概况如表1所示。
1.2 方法
用聚乙二醇(Polyethylene glycol,简写PEG)和赤霉素(Gibberellin,简写GA)处理连香树种子,PEG和GA属植物生长调节剂,具有促进植物营养器官的生长和发育的作用[5~7],同时,此类外源激素也表现出对种子发芽相应的影响,本文在不同温度下对不同种源地种子的发芽能力进行研究,以其为种子选育和萌发提供理论依据。
将饱满无机械损伤的连香树种子每组50粒分别装入50 ml小烧杯中、编号,然后采用0.2%的高锰酸钾(K2MnO4)消毒30 min[8],消毒后用蒸馏水清洗2次~3次,再用40℃蒸馏水浸泡催芽24 h;将聚乙二醇和赤霉素分别配置成浓度为25 mg·L-1、50 mg·L-1、100 mg·L-1和200 mg·L-14种;过滤浸泡催芽的种子,然后将种子放入烧杯,用不同浓度的激素浸泡12 h,再用蒸馏水清洗2次~3次,放入70 mm培养皿(底部垫2层纱布)中,加入少量蒸馏水保湿,对照组不加激素处理其他处理一致,每组处理3个,然后将装有种子的培养皿放入15℃、20℃、25℃和30℃的恒温箱中培养,总计240个处理,每天观察1次,加水1次保持湿度。
表1不同种源地基本概况
Tab. 1
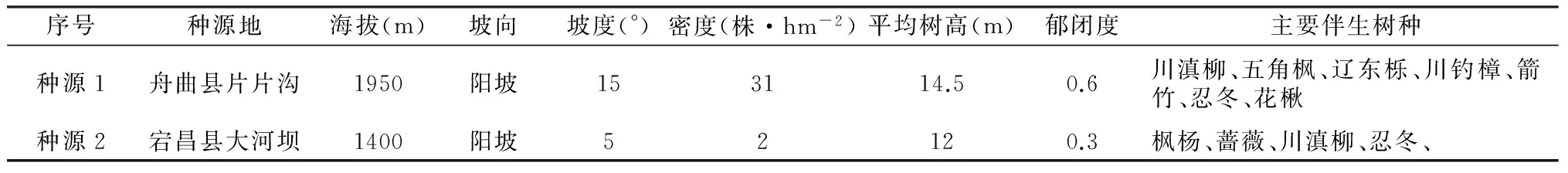
The basic situation of different provenance
注:本文图中种子来源于舟曲县的种子为种源1,种子来源于宕昌县的种子为种源2。
1.3 数据记录和处理
每粒种子以胚根突破种皮作为种子发芽的标志,以连续5 d没有种子发芽作为种子发芽结束标志,每天观察并记录种子发芽状况,按照以下公式计算种子的发芽率、发芽势、发芽指数[9]和平均发芽时间[10]。
发芽率=(正常发芽的种子数/供试种子数)×100%
发芽势=(规定时间内发芽的种子数/供试种子数)×100%
发芽指数(GI)=∑(Gt/Dt)式中Dt表示时间t日的发芽数,Gt表示相应的发芽日数。
平均萌发时间(MGT)=(A1T1+A2T2+…+AnTn)/(A1+A2+…+An)式中An为培养第n天的发芽数,Tn为相应的发芽天数。
数据处理采用Excel和spss13.0完成。
2 结果与分析
2.1 不同激素浓度在不同温度下对连香树种子发芽率的影响
植物激素是植物自身合成的调节物质,在种子发育过程中起着重要的调控作用[11],温度对激素活性也起着重要作用,不同温度下激素活性不同[12]。在不同温度下不同激素随着浓度的增加连香树发芽率如图1所示。
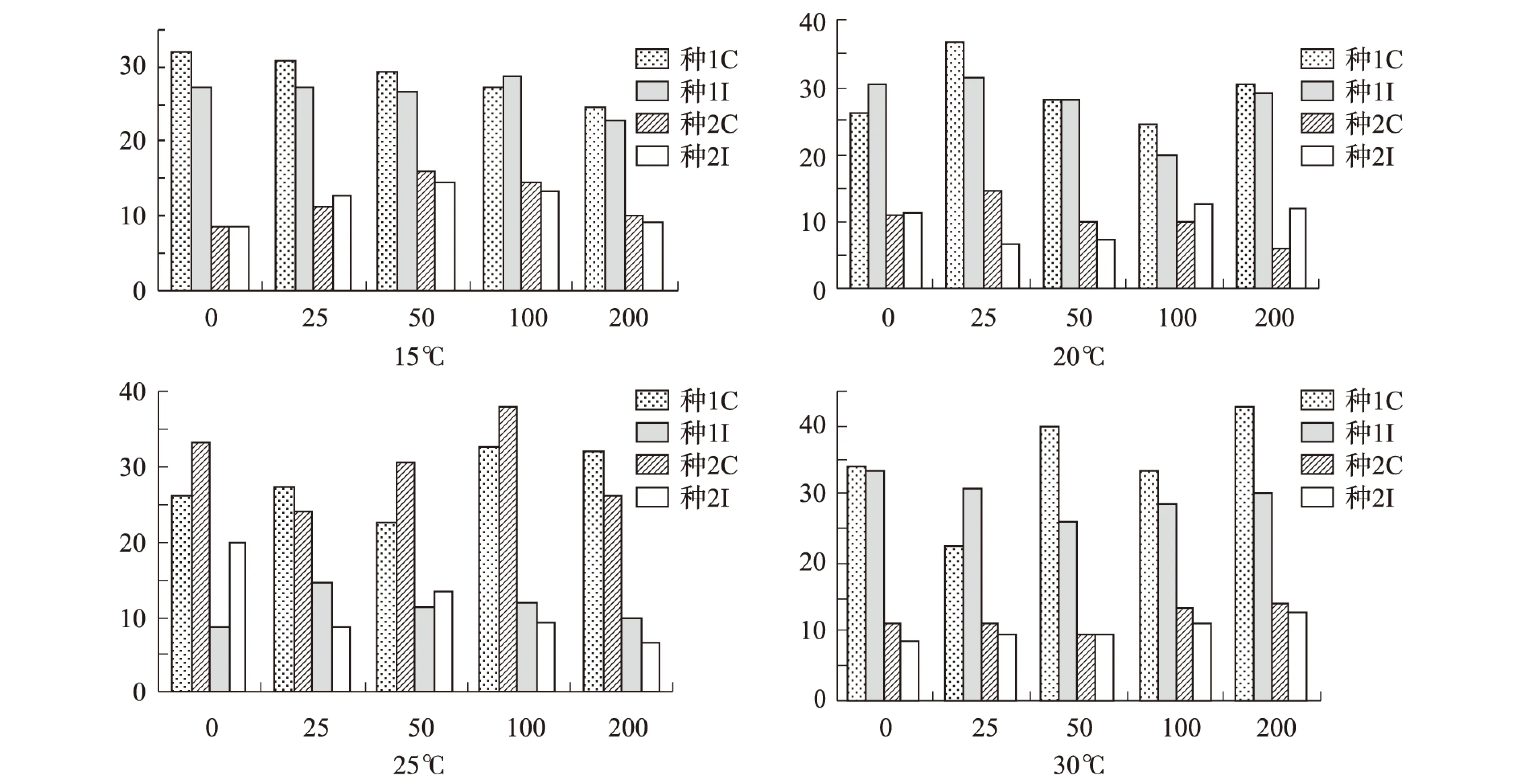
注:种1C指种1用GA处理,种1I指种1用PEG处理,种2C指种1用GA处理,种2I指种1用PEG处理,同下图1 不同温度随着激素浓度增加连香树种子的发芽率变化Fig.1 Katsura tree seed germination rate changes under different temperature with the increase of hormone concentration
由图1可知:在不同温度下不同种源地种子随着不同激素浓度增加种子发芽率不同。在15℃时未用GA处理的种源1发芽率最大(32.00%),用GA处理的种子随着GA浓度的增加种子发芽率逐渐减小,发芽率与GA浓度呈负相关性;用PEG处理的种源1随着浓度的增加和对照组相似变化不大,在浓度为100 mg·L-1时发芽率最大(28.67%),随后减小;种源2在15℃时用GA和PEG两种激素处理的结果一致,随着浓度的增加都表现出现增加后减小的趋势,且在两种激素浓度为50 mg·L-1时发芽率最大,分别是16.00%和14.67%,在同一激素处理下用GA处理的发芽率略大于用PEG,随着浓度的增加种源2发芽率表现出先增加后减小的趋势,这与杨玲[12]等的研究一致。 在20℃时用GA和PEG处理的种源1随着浓度的增加发芽率表现为先增加后减小的趋势,在浓度为25 mg·L-1时发芽率最大,分别是36.67%和31.33%;用GA处理的种源2发芽率变化和种源1一致,在浓度为25 mg·L-1时发芽率最大(14.67%),用PEG处理的种源2发芽率先减小后增大最后又变小,即在浓度为0 mg·L-1时发芽率大于在浓度25 mg·L-1和50 mg·L-1,在浓度为100 mg·L-1时发芽率最大(12.67%),用低浓度PEG处理种源2发芽率不及对照。在25℃时用两种激素处理的种源1在浓度为100 mg·L-1时发芽率最大,分别是32.67%和38.00%,与20℃时PEG处理的种源2一致,对照组大于在浓度25 mg·L-1和50 mg·L-1的发芽率。用GA处理的种源2在25 mg·L-1时发芽率最大(14.67%),用PEG处理的种源2在50 mg·L-1时发芽率最大(13.55%)。在30℃时用GA处理的种源1随着浓度的增加发芽率变化不大,且在浓度200 mg·L-1时发芽率最大(42.67%),其次为在浓度100 mg·L-1时发芽率为40.00%,然后是对照为34.00%,浓度25 mg·L-1最小为22.67%;其他3种处理是都表现出先减小后增大的趋势,在浓度50 mg·L-1时发芽率最小,种源1在浓度为0 mg·L-1发芽率最大为33.33%,种源2在两种激素浓度为200 mg·L-1发芽率最大,分别是14.00%和12.67%。只有在25℃时出现用PEG处理的种子发芽率大于GA处理,其他处理均表现出用GA处理的种子发芽率大于其他。
2.2 不同激素浓度在不同温度下对连香树种子发芽势和发芽指数的影响
种子的发芽势和发芽指数是重要的生物学指标[13],是反映种子质量优劣的主要指标之一,故二者比发芽率能更好的表现种子活力。由图2可知:在15℃时两种种子发芽势和发芽指数变化基本一致,用GA处理种源1的发芽指数和发芽势先升高后减小,在浓度25 mg·L-1时发芽指数和发芽势最大,分别是2.32和26.00%,发芽率也略小于对照组而大于其他处理,在15℃时用GA浓度25 mg·L-1处理的种源1活性最大;两种激素处理的种源2随着激素浓度的增加发芽指数和发芽势变化规律不明显,但用GA处理的种源2在浓度50 mg·L-1时发芽指数和发芽势最大,分别是1.23和12.00%,说明在15℃时用GA浓度50 mg·L-1处理的种源2活性最大。
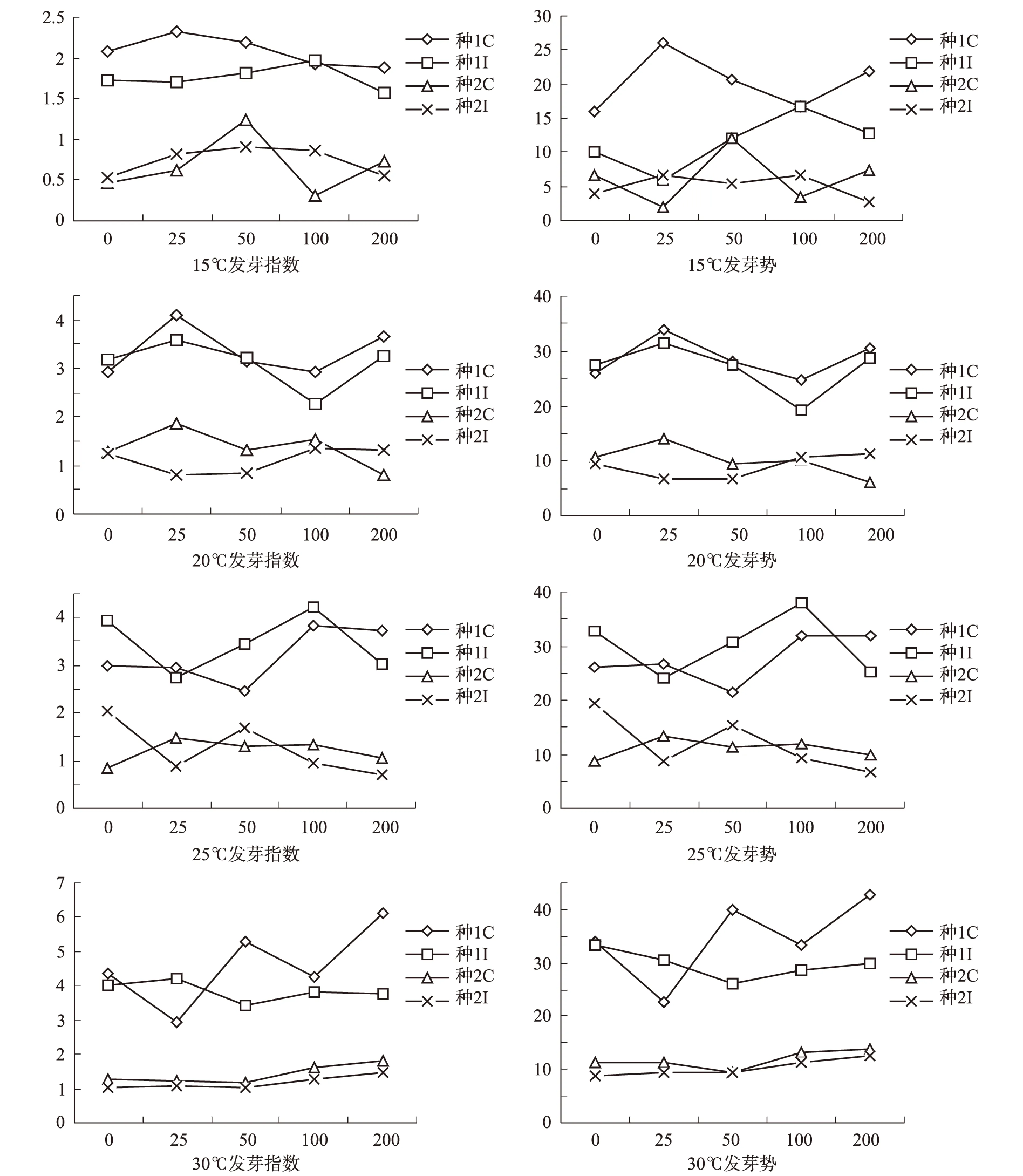
图2 不同温度随着激素浓度增加连香树种子的发芽指数和发芽势变化Fig.2 The katsure tree seed’s germination index and germination potential change under different temperature with the increase of hormone concentration
在20℃时两种种子发芽势和发芽指数变化一致,用PEG处理的种源1在浓度为25 mg·L-1时发芽势和发芽指数均最大,发芽指数为3.59,发芽势为31.33%;用GA处理的种源2也在浓度25 mg·L-1时发芽势和发芽指数均最大,分别是1.86和14.00%,用两种激素处理的种子,种源1的发芽势和发芽指数均大于种源2,在20℃时种源1的种子活性大于种源2。
在25℃时两种种子发芽势和发芽指数变化也一致,用PEG处理的种源1在浓度为100 mg·L-1时发芽势和发芽指数均最大,发芽指数为3.83,发芽势为32.00%;种源2不用激素处理时发芽势和发芽指数最大,分别是2.04和19.33%,用GA处理的种源2随着浓度的增加有减小的趋势,用PEG处理的种源2随着浓度的增加先增大后减小,在50 mg·L-1时为转折点,此时发芽率最大,发芽势和发芽指数较小。
在30℃时两种种子发芽势和发芽指数变化基本一致,用两种激素处理的种源1发芽势和发芽指数变化一致,但随着浓度升高用GA处理的种源1的2种指数无明显的变化规律,在浓度在200 mg·L-1时发芽势和发芽指数最大为6.12和42.67%;种源1随着PEG浓度的增加先减小后增加,在50 mg·L-1时最小;用两种激素处理的种源2在激素浓度为200 mg·L-1时最大,且用GA略大于PEG,分别是1.83和12.67%;随着温度的增加发芽势和发芽指数最大值也随着激素浓度的增加而加大。
2.3 不同激素浓度在不同温度下对连香树种子发芽时间的影响
不同激素浓度处理在不同温度条件下对种子的萌发时间有一定的影响[14~15],由图3可知:同一温度下随着浓度的增加平均发芽时间减少,对照组平均发芽时间最大,激素在200 mg·L-1时平均发芽时间最短;在15℃时用GA处理的种源1在200 mg·L-1时平均发芽时间最小为6.75 d;在20℃时用GA处理的种源2在200 mg·L-1时平均发芽时间最小为4.21 d;在25℃时用GA处理的种源1在200 mg·L-1时平均发芽时间最小为4.37 d;在30℃时用GA处理的种源1在200 mg·L-1时平均发芽时间最小为3.52 d,随着温度的升高同浓度平均发芽时间减小,高浓度激素有利于提高种子的发芽时间。
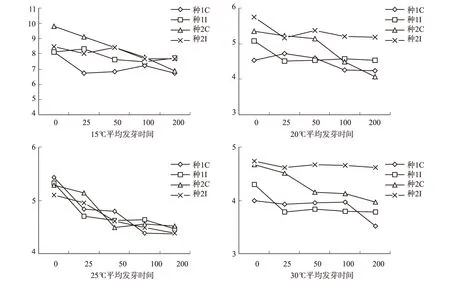
图3 不同温度随着激素浓度增加连香树种子的平均发芽时间变化Fig .3 The average seed germination time of katsura trees under different temperature with the increase of hormone concentration
3 结论与讨论
通过不同激素对不同种源地连香树种子在不同温度下种子发芽实验研究表明:不同种源地连香树种子发芽率与激素种类、激素浓度和温度都有一定的关系。
(1)GA处理的种子发芽率、发芽势和发芽指数均大于PEG处理的种子,说明GA比PEG更有利于连香树种子发芽。
(2)在同一温度下,随着激素浓度的增加,发芽率基本上表现出先增加后减小的趋势,不同温度下最大发芽激素浓度不同,种源1用GA和PEG发芽率均大于种源2,只有在25℃时出现用PEG处理的种子发芽率大于GA处理,其他处理均表现出用GA处理的种子发芽率大于其他;随着温度的升高,同一浓度激素处理的种子发芽率有升高的局势,及温度有利于激素活性,有助于种子发芽,这与黄建等研究基本一致[14],在30℃时用GA处理的种源1在浓度200 mg·L-1时发芽率最大为42.67%,种源2在25℃在GA浓度为25 mg·L-1发芽率最大为14.67%。
(3)同一温度下发芽势和发芽指数变化基本一致,两种激素处理的种源1发芽势和发芽指数均大于种源2,随着温度的增加发芽势和发芽指数最大值也随着激素浓度的增加而加大。
(4)同一温度下随着浓度的增加平均发芽时间减少,对照组平均发芽时间最大,随着温度的升高同浓度平均发芽时间减小,高浓度激素有利于提高种子的发芽时间。
不同种源地种子活力不一致,种源1大于种源2及舟曲县的连香树种子活力优于宕昌县的种子,GA更有利于连香树种子发芽,在不同的温度不同激素浓度下连香树种子发芽率、发芽势和发芽指数都不同,在种子催芽过程中找到最优发芽条件更有利于种子萌发,本研究仅对白龙江流域连香树种子萌发提供理论依据。
[1] 麦苗苗, 石大兴, 王米力, 等. PEG 处理对连香树种子萌发与芽苗生长的影响[J]. 林业科学, 2009, 45(10): 94~99.
[2] 李文良, 张小平, 郝朝运, 等. 珍稀植物连香树 (Cercidiphyllum japonicum) 的种子萌发特性[J]. 生态学报, 2008, 28(11): 5445~5453.
[3] 王静蓉, 段金廒. 连香树树皮化学成分的研究[J]. 植物学报, 1999, 41(2): 209~212.
[4] Ohtani T, Kamasaki K, Tanaka C. Effect of counterface material for abrasive wear of Cercidiphyllum japonicum wood on three-body abrasion[J]. Precision engineering, 2004, 28(1): 73~77.
[5] 李勇. 植物生长调节剂的作用及正确使用要点[J]. 新农村 (黑龙江), 2013 (22): 46~46.
[6] 张薇, 王文俊, 李连芳, 等. 外源激素浸种对云南松种子发芽的影响[J]. 种子, 2015, 4: 001.
[7] 李俊南, 熊新武, 习学良, 等. 植物激素对薄壳山核桃种子萌发及幼苗生长的影响[J]. 经济林研究, 2013, 31(1): 81~86.
[8] 袁宝东. 不同处理对厚朴种子发芽率的影响[J]. 防护林科技, 2017 (2): 27~28.
[9] 田宏, 刘洋, 张鹤山, 等. 扁穗雀麦种子萌发条件的研究[J]. 草业科学, 2009, 26(07): 88~93.
[10] 张蕾, 张春辉, 吕俊平, 等. 光照强度对青藏高原东缘九种紫草科植物种子萌发的影响[J]. 兰州大学学报: 自然科学版, 2011, 47(5): 67~72.
[11] 李晓宇, 蔺吉祥, 穆春生, 等. 外源植物激素对羊草种子形成及萌发的影响[J]. 中国农学通报, 2012, 28(23): 11~14.
[12] 杨玲, 崔晓涛, 沈海龙. 外源激素与温度对花楸树种子萌发的影响[J]. 植物生理学通讯, 2009 (6): 555~560.
[13] 黄光群, 黄晶, 张阳, 等. 沼渣好氧堆肥种子发芽指数快速预测可行性分析[J]. 农业机械学报, 2016, 47(5): 177~182.
[14] 黄建, 刘洪见, 钱仁卷, 等. 不同浸种温度及时间预处理对瞿麦种子发芽的影响[J]. 北方园艺, 2011 (15): 95~97.
[15] 张立磊, 王少平. 不同浸种温度和时间对观赏向日葵种子发芽的影响[J]. 种子, 2013, 32(12): 73~75.
EffectofTemperatureandHormoneofDifferentProvenancetoCercidiphyllumJaponicumSieb.etZucc.SeedGermination
CHEN Xue-long1,2WANG Fei1,2*DING Quan-ding1,2FU Zuo-lin1,2QI Hao1,2
(1.Institute of forestry Science,Bailongjiang Forestry Management Bureau of Gansu Province,Lanzhou,Gansu 730070,China;2.Gansu Bailongjiang National Forest Ecosystem Research Station, Zhouqu, Gansu 746300,China)
In order to study the influence of different temperature and hormone on the germination of the seeds in different area of the White Dragon River,two exogenous hormones, GA and PEG were used in this study.The germination experiment of the seed germination under different temperature gradient indicated that (1) the GA was more advantageous to the seed germination;(2) seed activity from Zhouqu was better than that of Dangchang seeds;(3) under the same temperature,as the hormone concentration increases, the germination rate showed a trend of decrease after increase first.With the increase of temperature,under the same concentration,seed germination rate showed increasing trend;Under the same temperature germination index and germination potential changes were consistent;(4) with the increase of hormone concentration and temperature,average germination time were reduced, and high concentration and high temperature could shorten seed germination time.
Hormones,Temperature,CercidiphyllumjaponicumSieb,Seed,Germination
2017-10-31
甘肃省青年科技基金计划项目(145RJYK278),甘肃省林业科技项目(2015kj048), 甘肃省林业科技项目(2015kj051),甘肃省林业科技项目(2016kj058), 甘肃省林业科技项目(2016kj062)。
陈学龙(1985-),男,本科,工程师,主要从事林业生态研究。E-mail:173763825@qq.com。
*通讯作者:王飞,男,工程师,主要从事林业土壤研究。E-mail:1336190642@qq.com。
10.16779/j.cnki.1003-5508.2017.06.009
S722.3
A
1003-5508(2017)06-0034-06
猜你喜欢
特产研究(2022年6期)2023-01-17
宁夏医科大学学报(2022年5期)2022-11-30
宁夏医科大学学报(2022年6期)2022-11-22
宁夏医科大学学报(2022年8期)2022-11-22
辽宁林业科技(2021年1期)2021-03-16
现代园艺(2020年19期)2020-10-02
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
武夷学院学报(2017年9期)2017-11-29
