
极小种群野生植物圆叶玉兰的生态位研究
2018-01-06 02:34:43陈小红
四川农业大学学报 2017年2期
曾 洪,陈小红
(四川农业大学林学院,四川温江 611130)
极小种群野生植物圆叶玉兰的生态位研究
曾 洪,陈小红*
(四川农业大学林学院,四川温江 611130)
【目的】研究极小种群圆叶玉兰(Oyama sinensis)的生态位特征,以期为圆叶玉兰野生资源的保护和种群扩大提供理论依据。【方法】在大川镇圆叶玉兰分布区域的7个海拔梯度布设21个20 m×20 m的样地,进行群落调查,研究群落中主要种群的生态位宽度和生态位重叠。【结果】研究区内圆叶玉兰在各海拔分布均匀;贴毛箭竹(Fargesia adpressa)、圆叶玉兰、领春木(Euptelea pleiospermum)、川莓(Rubus setchuenensis)、蜡瓣花(Corylopsis sinensis)有较大的生态位宽度,分别为1.907、1.870、1.778、1.746、1.697,在群落中具有种群优势;圆叶玉兰和领春木、贴毛箭竹、红花悬钩子(Rubus inopertus)等主要的伴生树种之间的生态位重叠较高,生态位重叠值均在0.5~1之间。【结论】目前,圆叶玉兰生存状况良好,生态适应能力较强,在群落中具有重要的生态地位和作用。
圆叶玉兰;重要值;生态位宽度;生态位重叠
生态位作为生态学领域研究的热点,能定量地反映物种之间和物种与环境之间相互作用的关系[1-2]。生态位理论在植物种间关系、群落结构、群落演替、生物多样性和濒危物种评价等方面得到广泛运用,在生物多样性保护和评价方面具有较高的生态学价值[3-4]。近年来关于小种群濒危植物的种群生态位的研究逐渐增多,这些研究主要从生态位角度来探讨濒危植物的种间竞争、共存特征及濒危机制,并取得了一些成果[5-8],但并未能提出一些普适性的对策和方法。
木兰科植物具有种群的原始性、生存上的濒危性、数量上的稀有性和应用的广泛性等特点,被列入我国重点保护珍稀濒危植物名录的有39种[9]。近年来,已有学者对木兰科小种群的濒危机制进行了一些研究。田昆等[10]对华盖木(Pachylarnax sinica)的生境脆弱性进行了深入分析;黄久香等[11]研究观光木(Michelia odora)的生态特征,认为其自然种群保持有较高的遗传多样性水平;王剑伟等[12]对宝华玉兰(Yulania zenii)小种群的种群分布格局和群落特征进行调查研究,分析其濒危现状;刘春生[6]对黄山木兰(Yulania cylindrica)的种群生态学进行研究;金俊彦等[13]对单性木兰(Woonyoungia septentrionalis)群落优势种群生态位进行了研究;潘春柳[14]对单性木兰的生殖生态学做了较系统的研究;郭连金等[15]对小花木兰(Oyama sieboldii)的生殖对策进行了研究;王立龙等[16-17]分别对小花木兰和单性木兰进行传粉生态学的研究等。然而这些研究相对集中于少数树种,若要为木兰科植物的保护和利用提供更为科学的理论依据,有必要对更多的木兰科珍稀种群展开全面且有针对性的研究。
圆叶玉兰(Oyama sinensis(Rehder et E.H.Wilson)N.H.Xia et C.Y.Wu)系木兰科(Magnoliaceae)天女花属(Oyama)落叶灌木或小乔木,国家Ⅱ级重点保护植物,是四川省特有种和优先拯救保护极小种群野生植物,分布于海拔1 800~2 500 m的林缘或灌丛[18-20]。圆叶玉兰地理分布狭窄,生境条件恶劣,数量稀少;其树皮可代厚朴药用,自然资源遭人为破坏严重;嫩芽、花茎、果实容易遭小动物啃食,这加大了圆叶玉兰种群的破坏程度。目前,有关极小种群野生植物圆叶玉兰种群生态学相关的研究和文献极其缺乏,圆叶玉兰生态位的研究更未见报道。因此,开展圆叶玉兰相关生态学的研究,探讨其濒危机制,对保护其原始生境和种质资源的意义重大。本文通过对大川镇圆叶玉兰分布群落的调查,研究分析了圆叶玉兰和其主要伴生种群的生态位特征、圆叶玉兰所在群落的稳定性及其在群落中的地位和作用,为圆叶玉兰的拯救保护和科学管理提供理论依据。
1 材料和方法
1.1 研究区自然概况
研究区位于大川镇黄水河大熊猫基因交流走廊带(E102°52'~103°11',N30°32'~30°49'),地处青衣江上游,坐落于西岭雪山腹地,面积约400 hm2,海拔范围1 800~2 500 m。所属中纬度内陆亚热带湿润气候,雨量充沛,四季分明,无霜期长,日照偏少。7、8月年降雨可达1 000~1 800 mm;年均气温为11℃,最高气温30℃,最低气温-3℃。土壤类型为山地黄棕壤,呈微酸性。该区域为天然次生林,植被分层现象不明显,位于山地针叶阔叶混交林带。圆叶玉兰多生长在沟谷的崖壁、陡坡处(马桑坪-汇源电站、小牛井公路沿线,小沟、磨坊沟、鹿厂沟沟谷两旁);该区地质活动频繁,自然环境复杂,生境条件恶劣。据调查,总面积为0.84 hm2的样地内,维管束植物共236种,隶属64科144属。木本植物34科126种占51.40%,草本植物33科110种占48.61%;其中蕨类9科13种,藤本植物4科9种;苔藓层盖度为25%~50%。该群落中常见优势科有蔷薇科(Rosaceae)、木兰科(Magnoliaceae)、樟科(Lauraceae)、杨柳科(Salicaceae)、槭树科(Aceraceae)、忍冬科(Caprifoliaceae)、禾本科(Gramineae)、菊科(Compositae)和虎耳草科(Saxifragaceae)。藤本多为五味子属(Schisandra)和猕猴桃属(Actinidia)植物。
1.2 研究方法
1.2.1 样方设置与调查
在对研究区全面踏查的基础上,采用典型样地法对圆叶玉兰分布群落进行样地设置和调查。样地面积设置为20 m×20 m,分别在7个海拔梯度(1 800、1 900、2 000、2 100、2 200、2 300、2 400 m) 处各设置3个样地点,共21个样地点,总面积为0.84 hm2。在每个样地内设置4个10 m×10 m的乔木层样方,在样地四角设置4个5 m×5 m的灌木样方,在样地四角和中间设置6个1 m×1 m的草本小样方。参照刘春生等[6-7]的方法,调查并记录样地内植物的种类、高度、胸径、地径和数量等指标,其中纳入乔木层统计的木本植物需符合DBH≥5 cm和H≥5 m的标准。对研究区重要值排名靠前,且出现在2个以上不同海拔梯度的主要种群进行的生态位研究,如贴毛箭竹(Fargesia adpressa)、圆叶玉兰(Magnolia sinensis)、卵叶钓樟(Lindera limprichtii)、川莓(Rubus setchuenensis)、领春木(Euptelea pleiospermum)、亮叶忍冬(Lonicera ligustrina)等。本文以这7个海拔梯度作为“资源位”,计算圆叶玉兰及其主要伴生种在不同海拔的重要值,进而计算它们的生态位宽度以及生态位重叠度,并分析它们在群落中的地位和资源利用情况。
1.2.2 数据分析
本文以重要值作为计策的状态指标,其计算公式为:重要值(IV)=(相对密度+相对频度+相对高度)/3[5];并以研究区圆叶玉兰所在的7个海拔梯度为“资源位”,研究群落内主要种群的生态位[8,21]。生态位宽度采用Shannon-Wiener计算公式:

式中,Bi为种i的生态位宽度;Pij=nij/Ni(nij为物种i在资源j上的重要值,Ni为物种i在所有资源上的重要值之和),Pij代表种i在资源j上的重要值占该种在所有资源上的重要值的比例,r为资源位总位数。
生态位重叠系数:指一定资源序列上,两个物种利用同等级资源且相互重叠的情况。计算公式如下:
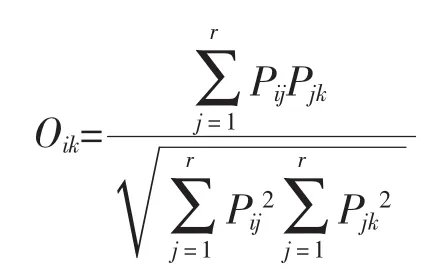
式中,Oik为物种i和物种k的生态位重叠系数;Pij和Pjk分别为物种i与物种k在资源j上的重要值;r为资源位总位数。
2 结果与分析
2.1 研究区木本层主要种群的重要值特征
由表1可知,领春木、圆叶玉兰、泡花树(Meliosma cuneifolia)和丝毛柳(Salix luctuosa)在乔木层和灌木层均出现,在7个海拔均有分布,且在灌木层具有较大的重要值和种群数量,说明这些种群在林内更新状况良好。占据灌木层第1位的贴毛箭竹的重要值之和为172.50,在各海拔分布均匀,种群数量多。相比之下,占据灌木层第2位,重要值之和为59.79的圆叶玉兰优势并不明显,其种群数量也较少。圆叶玉兰在乔木层的重要值排在第6位,在各海拔分布不均,重要值在海拔2 400 m处最大。结合圆叶玉兰在各海拔乔、灌层重要值的分析,圆叶玉兰在海拔2 100~2 400 m的重要值之和,在灌木层占其重要值总和的56.78%,在乔木层占其重要值总和的72.10%。由此可知,研究区内圆叶玉兰分布均匀,大树主要分布在2 100~2 400 m的海拔梯度上。
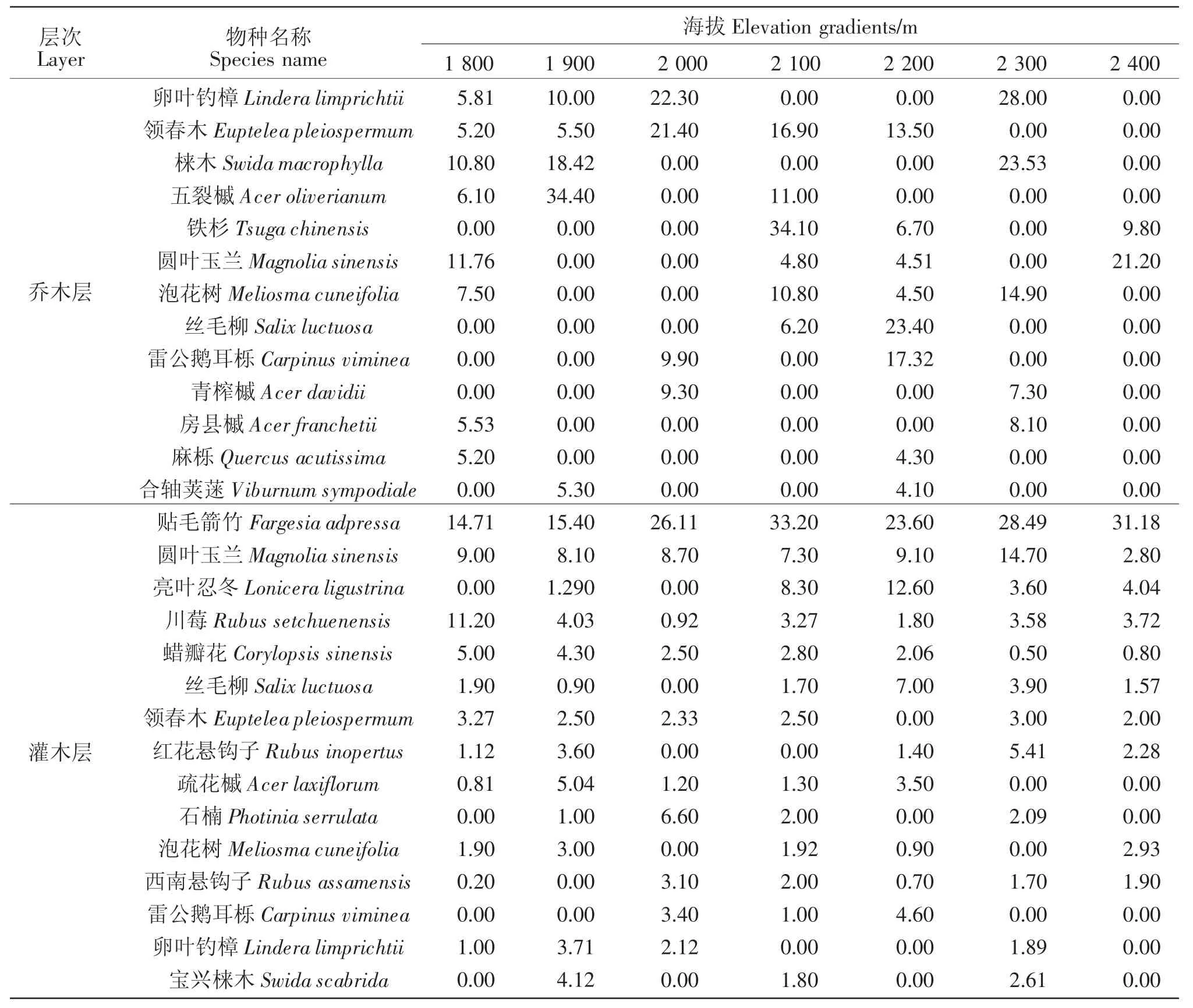
表1 木本层主要种群在不同海拔梯度中的重要值Table 1 Important values of the main population in the woody layer along the elevation gradients
2.2 种群生态位
2.2.1 生态位宽度
乔木层共37种植物,表2的12种植物的重要值之和占了总重要值的67.02%。由表2可知,乔木层卵叶钓樟、领春木、泡花树和圆叶玉兰的生态位宽度值于1.10~1.50之间,它们的海拔分布范围相对较广,生态适应力比其他树种强,在构建群落结构上起到重要作用。其余没有列入生态位宽度分析的种群,重要值偏低,或出现在两个以下海拔梯度,种群数量极少或为偶见种,它们生态适应能力不强或较弱,对生存环境和资源条件要求较为苛刻。
灌木层共89种植物,表2的15种植物的重要值之和占了总重要值的60.63%。通过表2可以看出,生态位宽度的大小与资源位占有的大小密切相关,物种的重要值越大,生态位宽度一般也越大。灌木层贴毛箭竹、领春木、蜡瓣花和川莓的生态位宽度较大,均在1.60~1.90之间,在各个海拔梯度上均有分布,生态适应力相对较强。在实际调查中发现以上4种植物为圆叶玉兰的主要伴生物种,重要值靠前,植株分布均匀。但亮叶忍冬、丝毛柳和石楠等物种,虽重要值排序较其他物种靠前,但生态位宽度较小,在各海拔分布不均匀,表明其生态适应区范围较窄,对生存环境要求稍高。而泡花树和西南悬钩子(Rubus assamensis)的重要值靠后,生态位宽度却远大于排序靠前的物种,在各个海拔梯度分布均匀,表明它们对不同环境的生态适应能力较强。
圆叶玉兰在灌木层的生态位宽度为1.870,在乔木层的生态位宽度为1.187,排名均靠前(见表2),在研究区内海拔分布范围较广,说明其生态适应能力较强。资源利用能力强的种群分布广且均匀,竞争能力也较强。圆叶玉兰、卵叶钓樟、泡花树、领春木在乔木层和灌木层均占有较高的生态位宽度,表现出种群优势,是群落中的建群种。资源利用能力弱的种群生态位较为狭窄,分布不均,如石楠(Photinia serrulata)、雷公鹅耳栎(Carpinus viminea)、宝兴梾木(Swida scabrida)等物种,生态位为宽度值排名靠后,且只在少数几个海拔集中分布。这基本反映出大川镇研究区群落中物种的优劣势地位及其生态特征。
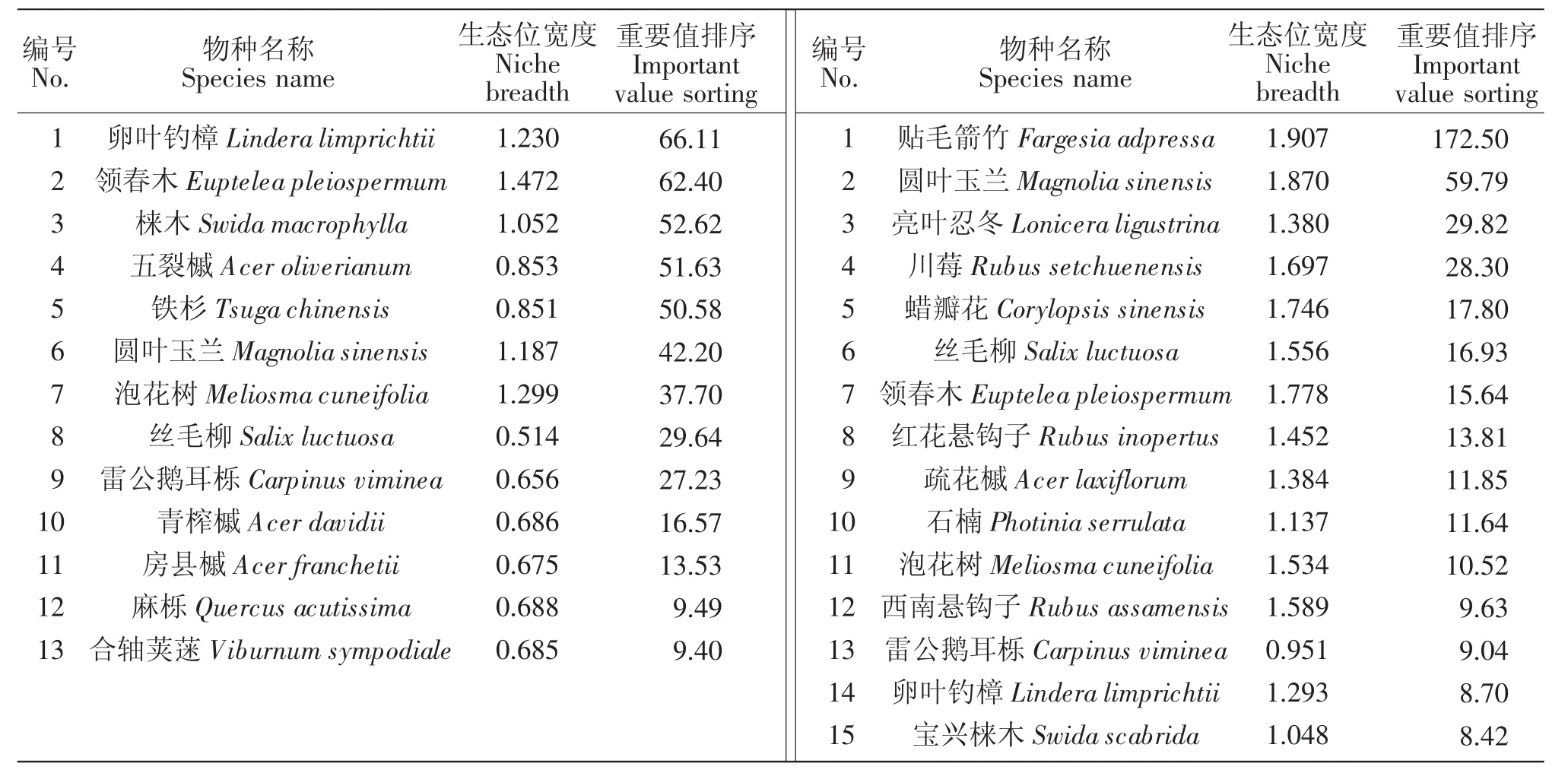
表2 木本层优势种群的重要值和生态位宽度Table 2 Important value and niche breadth of dominant population in the woody layer
2.2.3 生态位重叠
由表3可知,乔木层的生态位重叠值在0.8以上的有4对,分别是卵叶钓樟和青榨槭(Acer franchetii),梾木(Swida macrophylla)和房县槭,房县槭和泡花树,丝毛柳和雷公鹅耳枥;生态位重叠值在0.6~1之间的有12对;多数种对生态位重叠值偏低,在0.1~0.4之间,共48对,占总数的45.92%;还有16个种对之间的生态位重叠值不到0.1,如丝毛柳和卵叶钓樟、梾木和雷公鹅耳栎、五裂槭和雷公鹅耳枥等;乔木层物种间的生态位重叠度整体较低。由表4可知,灌木层生态位重叠值在0.8以上的有11对,分别是贴毛箭竹和圆叶玉兰,圆叶玉兰和领春木,圆叶玉兰和红花悬钩子等种对;在0.6~1的有47对;少数种对生态位重叠值较低,在0.1~0.4之间,有18对,占总数的23.10%;灌木层种对间的生态位重叠值均大于0.1,灌木层物种之间的生态位重叠度较高。
由图1看,木本层植物生态位重叠值低于0.1的种对只出现在乔木层,占总数的11.48%;灌木层生态位重叠值在0.4~1之间的种对比值明显高于乔木层,乔木层生态位重叠值介于0~0.4之间的种对明显多于灌木层;灌本层物种间的生态位重叠度较乔木层高。
生态位宽度较大的物种,它们之间的生态位重叠值也较大,如:贴毛箭竹和圆叶玉兰、川莓和蜡瓣花之间,其生态位重叠值均在0.850 0以上;而一些生态位宽度较小的物种间的生态位重叠值也较大,如房县槭和泡花树,其生态位重叠值达到了0.812 6。这与它们自身利用环境资源的相似程度,或与其物种自身的生物学特性有关[1,9]。乔木层圆叶玉兰与其他物种间的生态位重叠值多在0~0.4之间,生态位重叠度整体较低。圆叶玉兰与麻栎的重叠值最大,其次是铁杉、泡花树、房县槭、领春木、梾木、雷公鹅耳枥、五裂槭、合轴荚蒾、卵叶钓樟、丝毛柳;灌木层中,圆叶玉兰和其他树种生态位重叠值均在0.5~1之间,具有较高的生态位重叠度,与领春木的重叠值最大,其次为贴毛箭竹、红花悬钩子、蜡瓣花、丝毛柳、川莓、卵叶钓樟、西南悬钩子、宝兴梾木、亮叶忍冬(Lonicera ligustrina)、疏花槭(Acer laxiflorum)、石楠、泡花树。生态位重叠值越大,表明圆叶玉兰与它们对生态条件的要求越相似,是与圆叶玉兰共存的主要伴生物种。
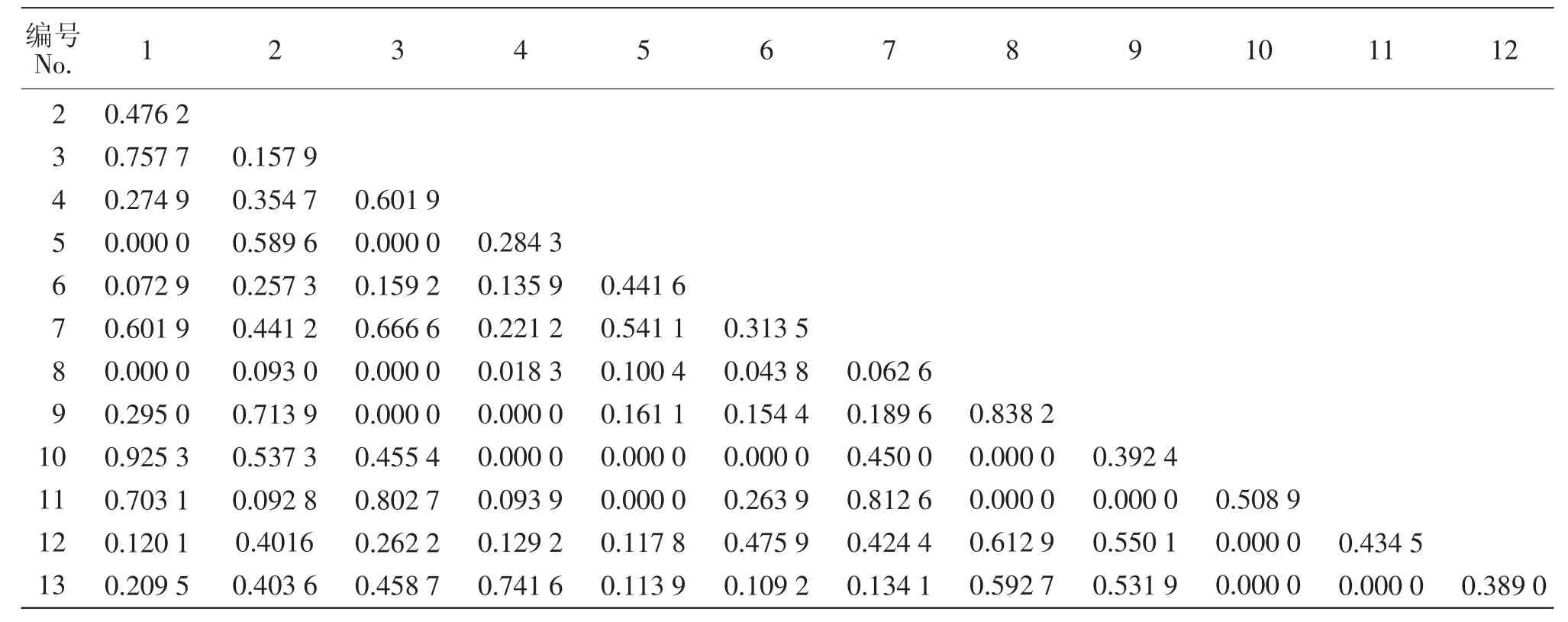
表3 乔木层13种植物的生态位重叠Table 3 Niche overlap of the 13 species in the shrub layer

表4 灌木层15种植物的生态位重叠Table 4 Niche overlap of the 15 species in the shrub layer

图1 木本层种间生态值重叠分配情况Figure 1 Distribution characteristics of value of niche overlap among population in the woody layer
3 讨论
研究区内物种数丰富,天然次生林林下更新良好,群落相对稳定。圆叶玉兰分布群落中泡花树、领春木、卵叶钓樟、贴毛箭竹、蜡瓣花、丝毛柳等树种,是圆叶玉兰主要伴生种和该区天然次生林的建群种。此外,圆叶玉兰分布区位于大川镇沟谷地段,生境条件恶劣,山体滑坡、坍塌和山洪等自然灾害频发,需对此采取相应的防范与补救措施,以防种群数量加速减少。
种群的生态位宽度,是对该物种资源利用和环境适应能力的一个衡量,是在群落中地位和作用的数量表达,以及间接反映种群在群落演替过程中的动态趋势[22-23]。本文研究发现,种群的生态位宽度与海拔分布范围密切相关。研究区内的乔木物种,如:铁杉、五裂槭,虽重要值排序靠前,但只分布在少数几个海拔梯度内,导致其生态位宽度较小。这与陈玉凯等[8]种群生态位宽度与自身地理分布范围密切相关的研究结果基本吻合。圆叶玉兰虽然在坡度较陡的沟谷生境中生长繁殖,但其生态位宽度值排名靠前,在适宜生境内分布较为均匀,表明其在特殊的生境中具有较强的适应能力,显示出种群优势。这符合一些生长于特殊生境中的珍稀保护植物具有较大生态位宽度的研究结果[8,24]。刘春生[6]研究的小种群濒危物种黄山木兰,其种群生态位宽度在群落中仍然较大;王立龙等[7]研究的极小种群植物小花木兰,其种群生态位宽度大小较小,但环境适应能力并不弱;本文研究的极小种群圆叶玉兰在研究区内也拥有较大的生态位宽度和较强的生态适应力。以上研究结果与毕晓丽[25]对珍稀植物群落学的研究结果一致,生态位宽度大小与植物濒危原因没有必然的联系。
生态位重叠值越大,物种之间对资源的利用程度越相似;相反,则表明物种之间对环境资源的需求差异大。从物种竞争角度看,生态位重叠值越大表明物种之间竞争越剧烈,但经过长期的自然选择,它们能够彼此互存,利用资源而彼此不相损害,形成相互影响且稳定的生态系统[1,26]。通过对圆叶玉兰分布群落内种群生态位重叠值情况和群落稳定情况的分析,本文认为在资源充足时,生境要求相似的种群能形成一定的共存关系,种间的生态位重叠值并不能完全反映种群间的竞争程度,这符合生态位重叠与竞争不成相关关系的学说[7,28]。圆叶玉兰是落叶灌木或小乔木,调查显示,自然生境中高度超过5 m以上的小乔木很少,大部分植株呈灌木状;在海拔1 800 m以上有分布,周围常见的伴生木本植物为贴毛箭竹、领春木、川莓、杜鹃属(Rhododendron)植物等;分枝低且平展,叶片和树冠阔大。这些特征表现出它的耐荫性和耐寒性,是适应山地沟谷阴湿环境,与其他种群形成共存关系的基础。群落在发展与演替中,在环境压力和种间相互适应的双重作用下,同一群落中不同物种通过不同途径利用资源,物种间能协同进化形成和谐统一的环境[25-26]。这可能是大川镇圆叶玉兰分布群落中许多生态位重叠的物种能够共存,形成相对稳定状态的原因;也是圆叶玉兰能够在群落中稳定生长和繁殖的关键。
综上所述,圆叶玉兰具有较强的生态适应能力,在群落中具有重要的生态地位和作用。但其地理分布狭窄,数量较少,对此类小种群濒危物种的保护重点应是加强其自然生境的保护和生殖繁育方面的研究,以便维持群落的连续性,保证圆叶玉兰种质资源的可持续性。此外,由于种群生态位的研究受时间的限制,随着群落的演替发展,种群的生态位宽度会发生变化,本文仅对圆叶玉兰目前生存现状进行研究;今后还需对圆叶玉兰的种群动态、生殖生理方面做进一步的探究,为保护濒危植物圆叶玉兰提供更多的科学依据。
[1]冶民生,关文彬,吴斌.岷江干旱河谷主要灌木种群生态位研究[J].北京林业大学学报,2006,28(1):7-13.
[2]苏志尧,吴大荣,陈北光.粤北天然林优势种群生态位研究[J].应用生态学报,2003,14(1):25-29.
[3]ALMEIDA R,GONALVES S,ROMANO A.In vitro micropropagation of endangered Rhododendron ponticum L.subsp.baeticum(Boissier&Reuter)Handel-Mazzett[iJ].Biodiversity and Conservation,2005,32(14):1059-1069.
[4]廖秋林,沈守云,卜国华,等.多花木兰人工生态恢复植物群落种群竞争及其生态位研究[J].中国农学通报,2012,28(31):71-74.
[5]陈俊华,刘兴良,何飞,等.卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征[J].林业科学,2010,46(3):23-28.
[6]刘春生.九龙山自然保护区珍稀濒危植物黄山木兰种群生态学研究[D].金华:浙江师范大学,2010.
[7]王立龙,王广林,黄永杰,等.黄山濒危植物小花木兰生态位与年龄结构研究[J].生态学报,2006,26(6):1864-1871.
[8]陈玉凯,杨琦,莫燕妮,等.海南岛霸王岭国家重点保护植物的生态位研究[J].植物生态学报,2014,38(6):576-584.
[9]刘玉壶.木兰科植物及其珍稀濒危种类的迁地保护[J].热带亚热带植物学报,1997,5(2):1-12.
[10]田昆,张国学,程小放,等.木兰科濒危植物华盖木的生境脆弱性[J].云南植物研究,2003,25(5):551-556.
[11]黄久香,庄雪影.观光木种群遗传多样性研究[J].植物生态学报,2002,26(4):413-419.
[12]王剑伟,张光富,陈会艳.特有珍稀植物宝华玉兰种群分布格局和群落特征[J].广西植物,2008,28(4):489-494.
[13]金俊彦,覃文更,罗柳娟,等.濒危植物单性木兰群落优势种群生态位研究[J].广西植物,2013,33(3):300-305.
[14]潘春柳.珍稀濒危植物单性木兰生殖生态学研究[D].南宁:广西大学,2007.
[15]郭连金,贺昱,徐卫红.三清山濒危植物天女花种群生殖对策研究[J].植物科学学报,2012,30(2):153-160.
[16]王立龙,王广林,刘登义,等.珍稀濒危植物小花木兰传粉生物学研究[J].生态学杂志,2005,24(8):853-857.
[17]赖家业,潘春柳,覃文更,等.珍稀濒危植物单性木兰传粉生态学研究[J].广西植物,2007,27(5):736-740.
[18]刘玉壶,罗献瑞,吴容芬,等.中国植物志[M].30卷.北京:科学出版社,1996:124.
[19]潘红丽,冯秋红,隆廷伦,等.四川省极小种群野生植物资源现状及其保护研究[J].四川林业科技,2014,35(6):41-46.
[20]中国科学院中国植物志编辑委员会.FOC.Flora of china[M].7卷.北京:科学出版社&密苏里州:密苏里植物园出版社,2013:67.
[21]林伟强,贾小容,陈北光,等.广州帽峰山次生林主要种群生态位宽度与重叠研究[J].华南农业大学学报,2006,27(1):84-87.
[22]颜廷芬,丛沛桐,刘兴华,等.环境因子对植物生态位宽度影响程度分析[J].东北林业大学学报,1999,27(1):35-38.
[23]吴明作,刘玉萃,杨玉珍,等.河南省栓皮栎林主要种群的生态位研究[J].西北植物学报,1999,19(3):511-518.
[24]王祥福,郭泉水,巴哈尔古丽,等.崖柏群落优势乔木种群生态位[J].林业科学,2008,44(4):6-13.
[25]毕晓丽.福建万木林珍稀植物群落学研究及其可持续经营[D].福州:福建农林大学,2002.
[26]杜峰,梁宗锁,胡莉娟.植物竞争研究综述[J].生态学杂志,2004,23(4):157-163.
[27]刘巍,曹伟.长白山云冷杉群落主要种群生态位特征[J].生态学杂志,2011,30(8):1766-1774.
Ecological Niche of Extremely Small Population of Wild Oyama sinensis
ZENG Hong1,CHEN Xiao-hong1*
(College of Forestry,Sichuan Agricultural University,Wenjiang 611130,Sichuan,China)
【Objective】The objective of the study was to explore ecological niche of extremely small population of wild O.sinensis and to provide a theoretical reference for wild resources protection and expansion of O.sinensis population.【Method】In the distribution area(Dachuan Town),21 plots(20 m×20 m)were set up in 7 elevations to investigate O.sinensis community composition.Ecologocial niche breadth andoverlapofmaintree.specieswereanalyzed.【Results】O.sinensiswasevenlydistributedateach altitude in the area.The niche breadth was relatively large for some species,including Fargesia adpressa(1.907),Oyama sinensis(1.870),Euptelea pleiospermum(1.778),Rubus setchuenensis(1.746)and Corylopsis sinensis(1.697),suggesting that these species had population advantage in the community.The niche overlap values were relatively high between O.sinensis and main associated tree species(e.g.,Euptelea pleiospermum,Fargesia adpressa,Rubus inopertus),with an range of 0.5-1.【Conclusion】Currently,the survival situation of O.sinensis was well.O.sinensis played an important role in the communities due to strong ecological adaptability.
Oyama sinensis;important value;niche breadth;niche overlap
Q948.1;S685.15 文献标志码:A 文章编号:1000-2650(2017)02-0220-07
10.16036/j.issn.1000-2650.2017.02.013
2016-12-09
国家林业局第二次全国重点野生植物资源调查项目(003Z0304)。
曾洪,硕士。*责任作者:陈小红,讲师,从事植物分类和野生植物资源保护与开发的研究,E-mail:493491274@qq.com。
(本文审稿:谢春平;责任编辑:巩艳红;英文编辑:徐振锋)
猜你喜欢
中外葡萄与葡萄酒(2022年1期)2022-01-21 09:29:10
散文诗(2021年22期)2022-01-12 06:14:04
红蜻蜓·中年级(2021年3期)2021-09-10 07:22:44
锦绣·上旬刊(2019年12期)2019-09-10 22:41:30
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
中老年健康(2016年6期)2016-06-30 10:50:16
茶叶学报(2016年4期)2016-03-29 03:24:48
河北遥感(2014年4期)2014-07-10 13:54:58
体育教学(2009年7期)2009-09-07 10:06:12
