
基于内生长土芯法探究不同林龄杉木人工林的营养限制因子
2017-12-25 06:59:31邱岭军胡欢甜王民煌葛露露林宝平何宗明王夏怡
福建农林大学学报(自然科学版) 2017年6期
邱岭军, 胡欢甜, 王民煌, 葛露露, 林宝平, 何宗明, 王夏怡
(1.福建农林大学林学院,福建 福州 350002;2.国家林业局杉木工程技术研究中心,福建 福州 350002;3.福建师范大学地理科学学院,福建 福州 350007)
基于内生长土芯法探究不同林龄杉木人工林的营养限制因子
邱岭军1,2, 胡欢甜1,2, 王民煌3, 葛露露1,2, 林宝平1,2, 何宗明1,2, 王夏怡1,2
(1.福建农林大学林学院,福建 福州 350002;2.国家林业局杉木工程技术研究中心,福建 福州 350002;3.福建师范大学地理科学学院,福建 福州 350007)
为探究不同林龄杉木林的营养限制因子,以3年生、14年生、21年生、46年生杉木为研究对象,运用内生长土芯法,监测杉木细根对不同施肥处理,即对照、+N溶液处理、+P溶液处理和+NPK混合溶液处理的响应.结果表明:试验进行12个月后,对照,+N、+P、+NPK处理的细根生物量变化范围依次为84.49~155.10、102.90~325.06、82.39~128.69、61.70~115.20 g·m-2,其中+N处理细根生物量变化最大.总生物量表现为对照>+N处理>+P处理>+NPK处理,杉木细根生物量随年龄的增大表现出先增大后减小的趋势,在速生期14 a达到最大值,呈单峰型.样地内3年生杉木人工林施加N、P元素更有利于杉木的生长,14年生杉木主要缺N元素,21年生杉木主要缺乏N、K元素,添加养分对46年生杉木的影响不明显.
杉木; 内生长圈; 养分限制; 蛭石; 细根
细根(直径≤2 mm)是植物地下生态系统的核心器官,也是最活跃的植物根系,因其生长和周转速率极快,对于整个丛林生态系统的物质循环和能量流动尤为重要[1-2].研究表明细根数量虽不到树木根系总量的30%,但每年消耗的净初级生产量(NPP)却占丛林生态系统总量的30%~80%[3],同时细根的死亡和分解过程向土壤归还的营养物质甚至超过地上凋落物[4].内生长法是指在设置内生长圈的过程中清除原有土壤,用蛭石填满内生长圈并添加特定比例的氮磷钾溶液,以保证12个月后野外试验取回的样品均是新近生长的细根[5],避免了测定过程中既有新根又有老根的问题[6].细根按照功能可以分为吸收根和先锋根两大类,吸收根(1级根)数量多、分布广,主要吸收养分和水分,而先锋根则起到构建根系结构的作用.
杉木(Cunninghamialanceolata(Lamb.) Hook.)是中国中亚热带地区重要的速生优良用材树种.我国杉木林面积为1.096×107hm2,占乔木总面积的6.7%[7].随着杉木连栽技术水平的提高,杉木人工林的林木产量不断提高,减轻对天然林依赖的同时,连栽导致的地力衰退和生产力下降现象日益突出[8].前人关于细根的研究主要集中在细根生产力[9]、生物量及分布[10]、细根水平分布[11].但在气候相同、土壤条件类似的区域内,通过内生长圈和添加不同养分方法相结合来探讨小生境中杉木养分限制因素还鲜有报道.传统的施肥方式不能揭示小范围内杉木林的养分限制因子.本研究以3年生、14年生、21年生2代杉木与46年生1代杉木林为研究对象,在整个林分受到人为干扰极小的状况下,通过细根生长量的变化判断不同林龄杉木人工林的养分限制因素,为解决杉木林连栽造成的地力衰退问题提供依据.
1 材料与方法
1.1 试验地概况
研究区位于福建省南平市峡阳国有林场 (东经117°59′,北纬26°48′),属武夷山南伸支脉,平均海拔238~650 m.该区为福建杉木主要产区,属中亚热带海洋季风气候,平均温度22.0 ℃(最高41.0 ℃,最低-5.8 ℃),3—9月为雨季,年均降水量为1 653 mm,平均湿度83%.土壤为绿泥片岩发育的红壤,土壤肥力较好,但均含有少量砾石.杉木人工林下植被有杨桐(Adinandramillettii)、芒萁骨(Dicranopterisdichotoma)、五节芒(Miscanthusfloridulus)、稀羽鳞毛蕨(Dryopterissparsa)、观音座莲(Angiopterisevecta)等.
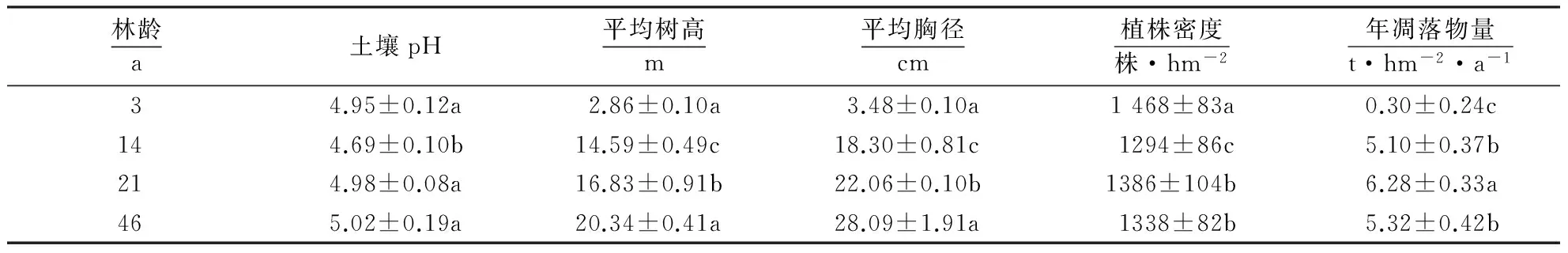
表1 不同林龄杉木人工林的生物量1)Table 1 Survey of the Chinese fir plantations with different stand ages
1)数据为平均值±标准误;不同小写字母表示差异显著(P<0.05).
1.2 标准地的设置
2013年1月在福建南平峡阳国有林场连栽杉木人工林,以2代3年生幼林、14年生近熟林、21年生成熟林和1代46年生过熟林为研究对象,在每个林龄随机选取4个立地条件相似且距离不大于100 m的25 m×25 m小区,共16个小区.在每小区内随机选取4株杉木,在根系宜生长周围均匀布置4个PVC生长圈,圈高10 cm,直径7 cm,表面共364个5 mm×1.7 mm的小孔.蛭石作为一种土壤改良剂,作为基质放入内生长圈,加入已配制好的4种不同营养溶液(对照:加入蛭石+100 mL去离子水.+N处理:蛭石+100 mL 6.76 g·L-1NH4Cl.+P处理:蛭石+100 mL 8.10 g·L-1Na2HPO4+NPK处理:蛭石+100 mL混合溶液 (2.25 g·L-1NH4Cl+2.7 g·L-1Na2HPO4+0.56 g·L-1KCl),用尼龙网(网眼3 mm)封住上下口.将生长圈埋入已挖好的深14 cm、直径8 cm的洞内,表层覆盖3 cm的枯枝落叶以及土壤,小区周围分别用红色绳子围起来,并用GPS记录每个样点的坐标以便后期采集.
1.3 样品的采集与分析
1.3.1 内生长圈的收集 本试验于12个月后,对收回的标准样地内64株标准木,设置4种不同溶液添加处理的内生长圈,按林龄(3、14、21、46 a)、小区号(1~4)、4种不同的施肥处理,分别装入已标记好的自封袋,当天带回室内进行处理.内生长圈取出后用周围的土壤填埋,以减轻人为对生态系统的干扰.
1.3.2 细根的处理 将野外收回的内生长圈倒入100目筛中,用自来水洗去蛭石等其他杂物,挑拣杉木细根,之后放置通风处自然风干.用Epson数字化扫描仪对细根形态进行扫描,然后用根系分析软件WinRHIZO对杉木根系长度和直径进行测量.比根长(SRL)=总根长/生物量.
1.3.3 土壤取样 去除每个林龄标准样地内的地表凋落物层,用直径为8 cm的土钻沿样地对角线分9个点,取0~10 cm土壤层样品,将同一土层的土样混和均匀后装在标记好的自封袋,带回备用.去除细根、杂物后进行风干,用自动球磨仪粉碎后过100目筛,采用全自动碳氮分析仪测定C、N含量.取一定量研磨好的土样,用浓硫酸—双氧水消煮法制备待测液.磷元素采用钼锑钪比色法测定,K、Ca、Mg以及交换性Ca、Mg含量均用原子吸收分光光度法测定.
1.4 数据处理
采用Excel 2003进行统计分析,使用方差分析(SPSS 17.0)和LSD检验分析不同林龄杉木细根的生物量之间的差异,相关图表采用Excel及Origin绘图软件完成.
2 结果与分析
2.1 蛭石与表层土壤的主要养分含量比较
对内生长圈中基质——蛭石相关元素进行测定,结果表明,蛭石含有的基本元素含量大体上与样地内土壤较为相近.土壤的氮含量表现为46 a>14 a>21 a>3 a,磷含量表现为14 a>3 a>21 a>46 a.在二代林中土壤氮、磷含量均表现为3 a<14 a,可能是3 a杉木林林下植被稀疏,地表生物量及枯枝落叶层较少.杉木人工林土壤钾含量表现为为3 a<14 a<21 a<46 a,随林龄的增大而升高.
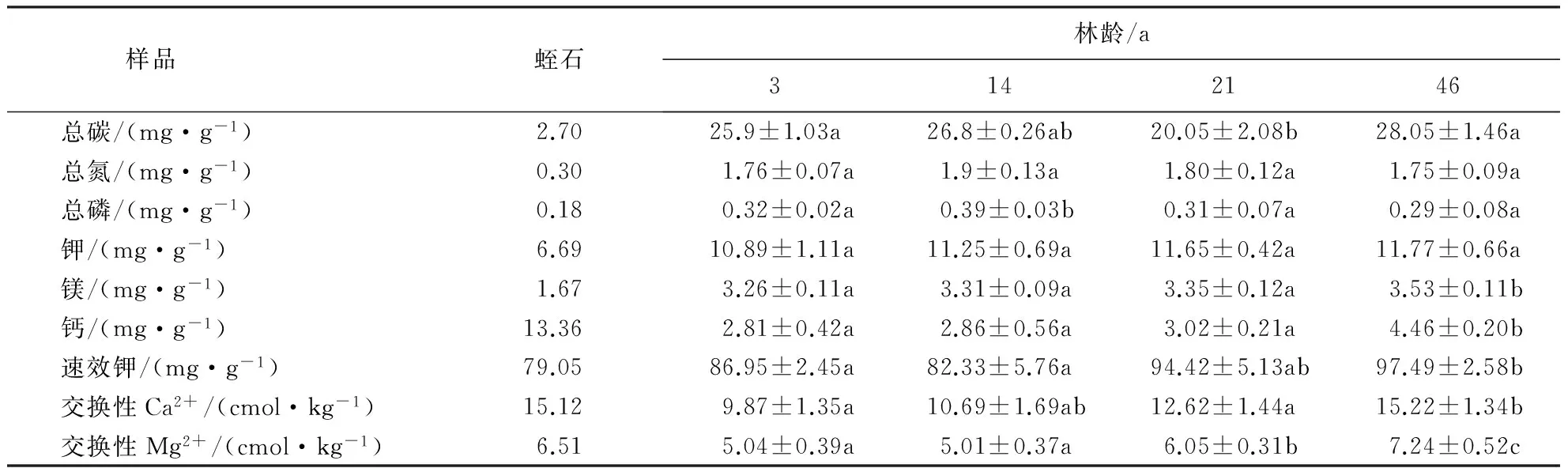
表2 不同林龄表层土壤与蛭石的养分含量1)Table 2 Nutrient contents of vermiculite and surface soils from plantations under different ages
1)数据为平均值±标准误;不同小写字母表示差异显著(P<0.05).
2.2 添加不同养分溶液对内生长圈中细根、草根生物量的影响
试验进行12个月后,不同林龄杉木林中内生长圈的细根总量表现为对照>+N处理>+P处理>+NPK处理,表明本试验设计合理.根据细根功能将其分为吸收根、先锋根.吸收根对照与+N处理均差异显著(P<0.05).研究表明,4种不同养分比例的草根生长量均呈U型,在21 a达到最低值;对照的草根生物量差异显著(P<0.05),表现为3 a>46 a>14 a>21 a.+N处理的草根生物量差异极显著(P<0.01),表现为46 a>3 a>14 a>21 a,呈U型,且在46 a达到最大值.+P处理的草根生物量差异显著(P<0.05),表现为3 a>46 a>14 a>21 a.+NPK处理的草根生物量差异显著(P<0.05),表现为3 a>46 a>14 a>21 a.
2.3 林龄和养分溶液添加处理对杉木细根生物量的影响
方差分析显示,在影响细根生物量的林龄以及4种不同养分溶液添加处理因子中,只有两两因子之间的交互作用对杉木细根有显著影响(P<0.05),4种不同养分溶液添加处理间对吸收根的影响差异显著(P<0.05),而不同林龄与交互作用间差异均不显著;4种不同养分溶液添加处理对先锋根的影响与不同年龄处理之间差异不显著.因此本试验主要根据吸收根的生物量变化来判断不同林龄连栽杉木林的养分限制因子.
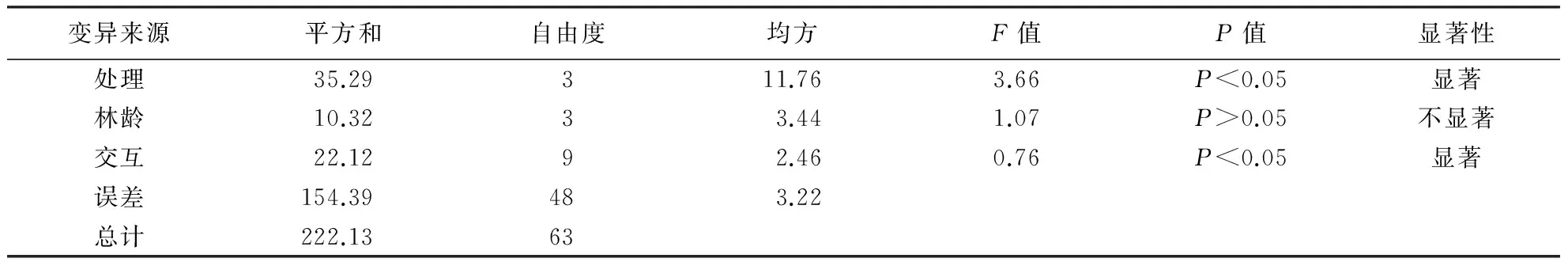
表3 4种处理对细根吸收根影响的显著性方差分析Table 3 Variance analysis of absorbing root under four treatments
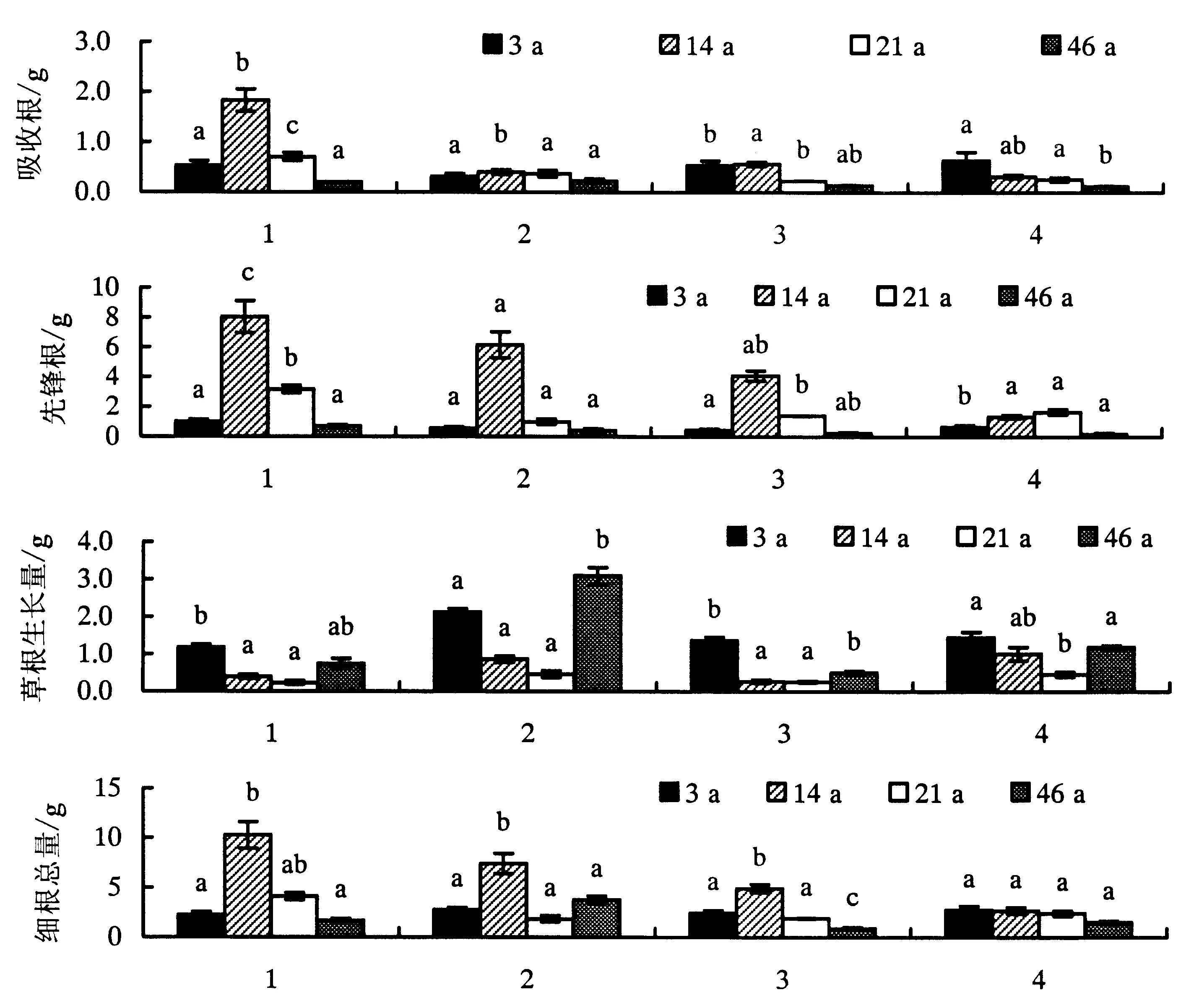
1.对照;2.+N溶液处理;3.+P溶液处理;4.+NPK处理.图1 不同处理措施下的内生长圈细根生长情况统计Fig.1 General growth stages of fine roots in inner growth rings under different treatments
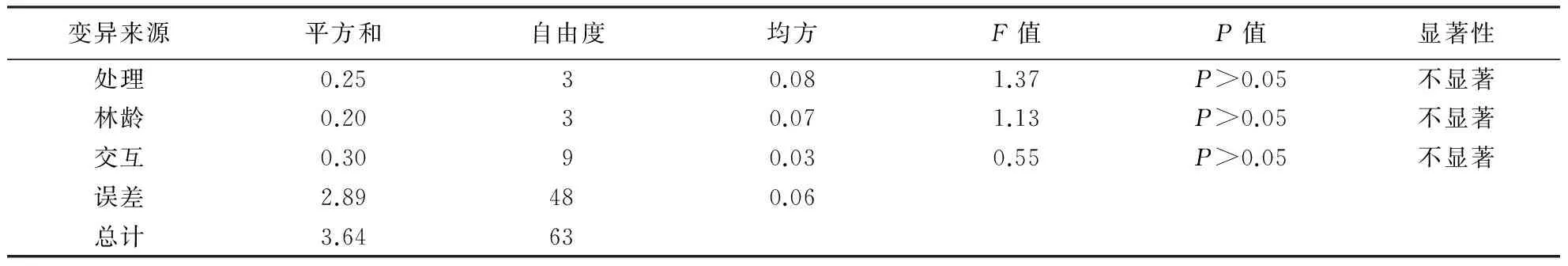
变异来源平方和自由度均方F值P值显著性处理0.2530.081.37P>0.05不显著林龄0.2030.071.13P>0.05不显著交互0.3090.030.55P>0.05不显著误差2.89480.06总计3.6463
3 小结与讨论
3.1 内生长圈中4种不同养分比例添加处理对细根生物量的影响
林木细根生物量与天气水文、土壤性质、林龄及人为干扰程度等有关[12-13].本研究杉木人工林细根生物量为444.56 g·m-2,与杉木生物量低于一般阔叶树种的细根生物量的研究结果[14]一致,与杉木生物量与同气候带的针叶林相似的研究结果[15]一致.Jackson et al[16]研究发现,细根每获取1个N元素须损耗40个单位的C元素.因此,土壤N的有效性直接影响到C向植物根系的分派,进而影响细根生长[17].不同树种施肥差异较大,既能导致林木细根生产量增加,又能导致其减少[18].对杨树(Populussimoniivar.przewalskii)施氮肥增加了细根生物量[19].但是,对花旗松(Pseudotsugamenziesii)施肥导致细根生物量减少40%[20].中亚热带地区是氮沉降最严重的区域,N含量已经不是该地区植物生长的限制性元素.杉木生长主要受P的限制,但内生长圈中+N处理细根产量>+P处理,表明试验区+N处理对细根生产量的影响还不确定.
内生长圈中添加不同营养元素对周围细根产生强烈的刺激作用,促进根系生长[21].本研究发现,杉木细根的总生物量表现为对照>+N处理>+P处理>+NPK处理,表明内生长圈设置合理,蛭石作为内生长圈的基质对细根具有明显的向导性,有利于根系向内生长圈延伸.蛭石因其良好的阳离子交换性和保水保肥性,有效地改善了细根生长土壤的物理环境.作为吸收养分的吸收根和存储养分的先锋根,+N、+P处理的产量都比+NPK处理的总量大,表明该区杉木缺乏N、P元素.而+NPK处理与对照相比较后发现先锋根占很大比例,说明该处理促进了根系的生长发育.研究发现,设置内生长圈后,不同林龄细根生物量明显增大,随着氮含量的提高,分配到细根中氮含量也升高,导致生物量增大.
3.2 通过对比细根生物量来判断不同林龄杉木林的限制因子
方差分析表明,内生长圈中营养元素的添加对不同林龄不同处理的吸收根的影响差异显著(P<0.05),对细根生长具有强烈的导向性;而对不同林龄不同处理的先锋根的影响差异不显著.杉木细根量总体表现为随年龄的增加逐渐降低,呈单峰型.研究发现14年生杉木细根量显著大于其他3个林龄的树木,14年生杉木林的吸收根和先锋根总量最大,处于快速生长发育期,养分添加的效果最明显.内生长圈设置后改善了小生境的土壤环境,透气性以及孔隙度等都有利于细根的生长.研究发现14年生杉木林表层土壤中的P含量明显高于其他林龄,可能是与林下枯枝落叶层较多及生物量丰富有关.而内生长圈中+N处理的细根总量最大,表明14年生杉木林主要受N元素限制;3年生杉木人工林处于幼龄期,组织发育不完善,需要大量的蛋白质和核酸维持生长,添加N、P元素更有利于其生长;21年生杉木林已经步入成熟阶段,但对比其他3种养分溶液处理的细根产量,添加+NPK混合溶液的细根产量最大,表明施加N和K元素有利于杉木生长.同时,由于46年生杉木已进入过熟阶段,细根和林木生长量均显著下降,表明内生长圈对其影响不明显.
3.3 不同林龄杉木土壤养分状况比较
通常C在大多数植物体内含量高且变异小,而N和P含量是最常见的限制性因子[22].土壤的N、P含量随林龄的变化而表现出有规律的增减,呈单峰型,14年生含量最高,46年生的则最低.14年生杉木处于快速生长阶段对养分需求量大,为了获取更多的养分和水分会把更多的营养分配至细根部分以适应环境[23].而幼林由于郁闭前水土肥流失较为严重,乔木层结构单一,46年生树木进入自然稀疏时期,林下植被增多带来的养分竞争导致土壤养分含量降低,这可能是导致土壤N、P含量下降的重要原因[24].不同林龄杉木土壤K、Mg、Ca、Ca2+、Mg2+等含量均随林龄的增加而提高,造成这种现象的原因可能是一代46年生杉木林土壤含量大于二代土壤含量[25-26].杨玉盛等[27]研究表明连栽致使某种养分缺失,循环速率降低,加上人为干扰破坏,使林地土壤的生物化学循环和土壤肥力向不利于林木生长的方向发展.因此,选育较大重吸收率、耐贫瘠的杉木树种是应对连栽导致地力衰退的有效措施.
[1] SONG X, LI Q, GU H. Effect of nitrogen deposition and management practices on fine root decomposition in Moso bamboo plantations[J]. Plant & Soil, 2017,410(1-2):1-9.
[2] CHEN H, HARMON M E, SEXTON J. Fine-root decomposition and N dynamics in coniferous forests of the Pacific Northwest, USA[J]. Can J For Res, 2002,32:320-331.
[3] GORDON W S, JACKSON R B. Nutrient concentrations in fine roots[J]. Ecology, 2000,81(1):275-280.
[4] JAKUB O, MAREK J K, MICHAEL B L, et al. Fine root production varies with climate in balsam fir (Abiesbalsamea)[J]. Canadian Journal of Forest Research, 2012,42:364-374.
[5] ZHANG Y H, NIU J Z, YU X X, et al. Effects of fine root length density and root biomass on soil preferential flow in forest ecosystems[J]. Forest Systems, 2015,24(1):12.
[6] 刘金梁,梅莉,谷加存,等.内生长法研究施氮肥对水曲柳和落叶松细根生物量和形态的影响[J].生态学杂志,2009,28(1):1-6.
[7] 李斌,方晰,项文化,等.湖南省杉木林植被碳贮量、碳密度及碳吸存潜力[J].林业科学,2013,49(3):1 001-7 488.
[8] 孙启武,杨承栋,焦如珍.江西大岗山连栽杉木人工林土壤性质的变化[J].林业科学,2003,39(3):1 001-7 488.
[9] 陈光水,杨玉盛,何宗明,等.福建柏和杉木人工林细根生产力、分布及周转的比较[J].林业科学,2004,40(4):15-21.
[10] 匡冬姣,雷丕锋.不同林龄杉木人工林细根生物量及分布特征[J].中南林业科技大学学报,2015,35(6):70-74.
[11] 陈光水,杨玉盛,何宗明,等.树木位置和胸径对人工林细根水平分布的影响[J].生态学报,2005,25(5):1 007-1 011.
[12] 胡芳,杜虎,曾馥平,等.广西不同林龄喀斯特森林生态系统碳储量及其分配格局[J].应用生态学报,2017,28(3):721-729.
[13] IDOL T W, POPE P E, FJR P. Fine root dynamics across a chronosequence of upland temperate deciduous forests[J]. Forest Ecology & Management, 2000,127(1-3):153-167.
[14] ZHANG X Q, WU K H. Fine-root production and turnover for forest ecosystems[J]. Scientia Silvae Sinicae, 2001,37(3):126-137.
[15] WANG S, WANG Z, GU J. Variation patterns of fine root biomass, production and turnover in Chinese forests[J]. Journal of Forestry Research, 2017,5:1-10.
[16] JACKSON R B, MOONEY H A, SCHULZE E D. A global budget for fine root biomass, surface area, and nutrient contents[J]. Proceedings National Academy Sciences of the United States of America, 1997,94:7 362-7 366.
[17] HENDRICKS J J, HENDRICK R L, WILSON C A, et al. Assessing the patterns and control soffine root dynamics: anempiricaltest and method logical review[J]. Journal of Ecology, 2006,94(14):40-57.
[18] 郭大立,范萍萍.关于氮有效性影响细根生产量和周转率的四个假说[J].应用生态学报,2007,18(10):2 354-2 360.
[19] KERN C C, FRIEND A L, JOHNSON J M F, et al. Fine root dynamics in a developingPopulusdeltoidesplantation[J]. Tree Physiology, 2004,24(6):651.
[20] GUNDERSON P, BERG B, CURRIE W S, et al. Carbon-nitrogen interactions in forest ecosystems; final report[J]. Open Research Online, 2015,71(7):1-23.
[21] 陈龙池,廖利平,汪思龙,等.酚类物质对杉木幼苗15N养分吸收、分配的影响[J].植物生态学报,2002,26(5):525-532.
[22] HENDRICKS J J, ABER J D, NADELHOFFER K J, et al. Nitrogen controls on fine root substrate quality in temperate forest ecosystems.[J]. Ecosystems, 2000,3(1):57-69.
[23] 曹娟,闫文德,项文化,等.湖南会同不同年龄杉木人工林土壤磷素特征[J].生态学报,2014,34(22):6 519-6 527.
[24] 邓浩俊,陈爱民,严思维,等.不同林龄新银合欢重吸收率及其C∶N∶P化学计量特征[J].应用与环境生物学报,2015,21(3):522-527.
[25] 曾德慧,陈广生,陈伏生,等.不同林龄樟子松叶片养分含量及其重吸收效率[J].林业科学,2005,41(5):21-27.
[26]李冬琴,谭益民,路宗岩,等.杉木炭疽病拮抗菌AM53菌剂的研制及其林间防效[J].福建农林大学学报(自然科学版),2015,44(2):154-158.
[27] 杨玉盛,何宗明,陈光水,等.杉木多代连栽后土壤肥力变化[J].生态环境学报,2001,10(1):33-38.
Applicationofroot-ingrowthcorestoassessnutrientlimitingfactorsofCunninghamialanceolataneedleleavesunderdifferentages
QIU Lingjun1,2, HU Huantian1,2, WANG Minhuang3, GE Lulu1,2, LIN Baoping1,2, HE Zongming1,2, WANG Xiayi1,2
(1.College of Forestry, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China;2.Chinese Fir Engineering Technology Research Center, State Forestry Administration, Fuzhou, Fujian 350002, China;3.College of Geographical Science, Fujian Normal University, Fuzhou, Fujian 350007, China)
To identify nutrient limiting factors ofCunninghamialanceolata, nitrogen (N), phosphors (P) and compound fertilizer of N, P and potassium (K) were applied toCunninghamialanceolataplantations of 3, 14, 21 and 46 years old. Then how root response to nutrients was monitored by root-in growth core. The results showed that biomass of fine root from N fertilizer varied the most among all treatments after 12-month application. The total biomass of fine root from different treatments in a descending order was deionized water as the control, +N, +P, and +NPK. Vermiculite proved to as a proper holding capacity of water holding capacity of the substrate and promotes the growth of fine root. Fertilizer efficacy firstly increased with stand age and then drop when the stand aged, with biomass peaking at 14-year-old stand. To summarize, N and P fertilization facilitate the growth of 3-year-old stand significantly. And 14-year-old stand is more likely to be deficient in N and 21-year-old stand will both need N and K fertilizers. While 46-year-old stand did not response to fertilization.
Cunninghamialanceolata; ingrowthcores; nutrition limitation; vermiculite; fine root
2017-03-01
2017-05-12
国家重点研发计划项目资助(2016YFD0600300) .
邱岭军(1991-),男,硕士研究生.研究方向:森林理水与保土功能.Email:447446583@qq.com.通讯作者何宗明(1965-),男,研究员.研究方向:水土保持、森林培育.Email:hezm2@126.com.
S714.8
A
1671-5470(2017)06-0648-06
10.13323/j.cnki.j.fafu(nat.sci.).2017.06.008
(责任编辑:叶济蓉)
猜你喜欢
矿产综合利用(2021年1期)2021-03-15 05:57:24
防护林科技(2020年10期)2020-12-22 11:46:42
水土保持研究(2020年2期)2020-03-19 09:03:26
乡村科技(2019年21期)2019-02-21 09:19:29
生态学报(2018年17期)2018-10-18 02:26:48
四川农业大学学报(2018年3期)2018-09-04 08:38:36
广州化工(2016年24期)2017-01-17 07:39:41
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
化工矿产地质(2015年3期)2015-12-04 02:09:40
