
火烧和砍伐对疏叶骆驼刺叶片生态化学计量特征的影响
2017-12-23 02:44赵生龙曾凡江李尝君高欢欢
水土保持研究 2017年1期
赵生龙,曾凡江,张 波,李尝君,刘 波,高欢欢
(1.中国科学院 新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室,乌鲁木齐830011;2.新疆策勒荒漠草地生态系统国家野外科学观测试验站,新疆 策勒848300;3.中国科学院干旱区生物地理与生物资源重点实验室,乌鲁木齐830011;4.中国科学院大学,北京100049)
火烧和砍伐对疏叶骆驼刺叶片生态化学计量特征的影响
赵生龙1,2,3,4,曾凡江1,2,3,张 波1,2,3,4,李尝君1,2,3,4,刘 波1,2,3,高欢欢1,2,3,4
(1.中国科学院 新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室,乌鲁木齐830011;2.新疆策勒荒漠草地生态系统国家野外科学观测试验站,新疆 策勒848300;3.中国科学院干旱区生物地理与生物资源重点实验室,乌鲁木齐830011;4.中国科学院大学,北京100049)
以塔克拉玛干沙漠南缘绿洲—沙漠过渡带的优势植物——疏叶骆驼刺为研究对象,设置春季砍伐、春季火烧、秋季砍伐和对照处理,研究了干扰对疏叶骆驼刺叶片N,P,K生态化学计量特征的影响。结果表明:春伐和火烧处理下,疏叶骆驼刺叶片N,P和K含量随季节变化显著降低,而N:P无明显差异;秋伐处理下,叶片N含量及N:P先降低后上升,K含量先升高后降低,P含量则无明显差异。不同干扰下疏叶骆驼刺叶片N,P,K含量及其N:P的季节变异分析显示,火烧处理下其N,P,K季节变异最大,分别为:22.32%,20.43%,28.27%;春伐处理下,N及N:P季节变异最小,分别为16.29%,8.67%;秋伐处理下P季节变异最小,为8.64%;对照处理下,K季节变异最小,为8.11%,N:P季节变异最大,为24.26%。生长季节与干扰处理对疏叶骆驼刺叶片N,P,K含量及其N:P影响的方差分析显示,疏叶骆驼刺叶片N及N:P主要受生长季节与干扰的交互作用影响,而P及K含量主要受生长季节的影响。
疏叶骆驼刺;砍伐;火烧;N,P,K;季节变化
氮(N)、磷(P)和钾(K)作为植物生长发育所必须的主要元素,它们对植物的生长和生理机能的调节起着非常重要的作用[1]。国外对N,P,K的研究及应用较多,而国内起步相对较晚,但发展较为迅速。国内多位学者从不同角度(包括不同空间尺度、不同植被类型以及不同演替阶段)对其进行了研究[2-6]。它不仅有助于研究植物养分平衡,而且在揭示植物通过改变养分利用策略,适应环境变化方面具有重要意义。
疏叶骆驼刺(Alhagi sparsifoliaShap.)是塔南沙漠—绿洲过渡带的主要优势植物种之一[7-8],分布面积广,在极端干旱环境下适应性较强。自然条件下生长的疏叶骆驼刺群落具有较大的生态防护效益和社会经济价值,是策勒绿洲前沿风沙灾害的主要生态屏障,其生态防护效益主要体现在防风固沙、抑制草场退化、改善周围环境等方面[8]。疏叶骆驼刺还是一种优良的牧草资源,可以减轻人畜争粮矛盾,具有较大的经济价值[9]。随着塔南地区人口的不断增加,以及当地居民的过度放牧和采伐,疏叶骆驼刺植被遭受大面积的破坏。砍伐和火烧作为当地主要的干扰方式,对绿洲—沙漠过渡带的自然植被带来很大影响。有研究表明,火烧和砍伐对疏叶骆驼刺的更新以及生长有着重要的影响[10-11]。但它对疏叶骆驼刺叶片N,P,K化学计量的研究较少。同时,国内外对于植物N,P,K虽展开了大量研究,但多集中于湿地、森林和草地,对荒漠生态系统研究较少。
鉴于此,针对塔克拉玛干南缘的疏叶骆驼刺优势植物群落的几种常见的干扰方式(春伐、秋伐、火烧),通过对不同干扰方式下疏叶骆驼刺叶片的N,P,K含量特征进行研究,进一步揭示疏叶骆驼刺在不同利用方式下的养分限制状况,以期为更加合理地开发与利用疏叶骆驼刺提供科学理论依据。
1 研究区域与研究方法
1.1 研究区概况
研究区位于塔克拉玛干沙漠南缘的策勒绿洲—沙漠过渡带,试验在新疆策勒荒漠草地生态系统国家野外科学观测试验站的长期监测样地内进行。策勒国家站位于策勒绿洲西北部,地理坐标80°03′24″—82°10′34″E,35°17′55″—39°30′00″N,平均海拔为1 371 m。策勒绿洲的东西两个方向被自然植被所包围,南部与流动沙丘和戈壁相连,北部与塔克拉玛干沙漠连接。该地区属于暖温带极端干旱荒漠气候,夏季炎热,干旱少雨,光照充足,日照时间长,昼夜温差大,风沙灾害尤为严重。年平均气温11.9℃,极端最高气温41.9℃,极端最低气温-23.9℃。年均降水量为35.1 mm,且主要集中在5月和7月。蒸发量高达2 595.3 mm,干燥度20.8。绿洲外围主要植物种有疏叶骆驼刺、多枝怪柳等草本和灌木。其前沿过渡带分布的5~10 km的自然植被,是当地牲畜饲料和薪柴的重要来源[12]。
1.2 试验设计
本研究选用策勒绿洲—荒漠过渡带间自然生长的多年生疏叶骆驼刺作为研究对象。于2013年在策勒绿洲—荒漠过渡带间选择地势平坦、地下水位一致的自然生长的疏叶骆驼刺群落,设置16个10 m×10 m长期固定的试验样地,并用围栏保护,防止外界人为或牲畜的干扰。群落内主要以疏叶骆驼刺为主,伴生有花花柴[Kareliniacaspia(Pall.)Less]、怪柳(Tamarix ramosissima)以及沙蓬[Agriophyllum squarrosum(L.)Moq.]等短命植物等。其中12块试验样地用3种不同的处理方法来代表当地主要的人为干扰方式:(1)春季砍伐(CS)。代表人类在绿洲—沙漠过渡带上开垦过程中的一种干扰方式,处理方法为:在2014年4月将样地内所有疏叶骆驼刺植株齐地面砍伐;(2)秋季砍伐(CF)。代表当地居民常规和传统的收获利用方式,砍伐方法和春季砍伐一样,砍伐时间为2013年10月;(3)春季火烧(BS)。代表绿洲—沙漠过渡带上人类在开垦过程中的另一种干扰方式,处理方法为:在2014年4月对样地内的疏叶骆驼刺植被地上部分进行人为火烧处理;(4)对照(CK)。自然生长的疏叶骆驼刺样地。
1.3 采样和指标测定
分别于2014年8月、9月、10月在每个处理下的4块样地中选择8~10株生长良好且没有遮阴的个体作为取样植株,选取样株时尽量保证同一处理下的植株大小一致,在选定的样株上采集上部位带健康、成熟叶片的小枝,带回实验室后将其茎、叶、刺分开,后装入自封袋。首先将植物样品置于105℃的烘箱中烘15 min进行杀青处理,然后降低烘箱温度至75℃烘干至恒量。对烘干后的疏叶骆驼刺叶片进行粉碎,过100目筛,装袋封存,用于N,P,K的测定。按照国家有关标准分析方法[13],N含量采用FOSS全自动定氮仪测定,P含量采用高氯酸、硫酸消化,钼锑抗比色法测定,K的测定采用火焰光度法,使用Cary 60紫外分光光度计测定,测定结果以单位质量的养分含量(g/kg)表示。
1.4 数据处理
数据的前期处理用Excel 2007软件完成,并使用Origin 8.0软件作图。不同干扰处理下的疏叶骆驼刺N,P,K含量差异显著性检验采用单因素方差分析方法,变异系数(coefficient of variation,CV)用公式CV=标准误差/平均值×100%计算得出。不同干扰处理和生长阶段对疏叶骆驼刺叶片N,P,K化学计量特征的影响结果采用两因素方差分析方法分析。所有数据均采用SPSS 19.0统计软件进行处理与分析,显著度水平为0.05。
2 结果与分析
2.1 不同干扰处理下疏叶骆驼刺叶片N,P,K以及N:P含量的动态变化特征
不同干扰下疏叶骆驼刺叶片N,P,K含量及其N:P从生长旺季到衰退期内的变化规律不一致,并且不同元素在不同干扰下差异显著(图1)。4种干扰下不同生长阶段的N含量变化范围为10.01~22.40 g/kg,平均值为(16.09±3.75)g/kg。N含量在对照中先升高后保持稳定,春伐干扰下是先升高后降低,秋伐则是先降低后上升,而火烧干扰下疏叶骆驼刺叶片N含量随着植物的生长逐渐降低(图1A)。P含量在不同生长阶段的变化范围为0.84~1.69 g/kg,平均值为(1.24±0.22)g/kg。P含量在火烧处理最高,并随着疏叶骆驼刺的衰老,P含量显著降低;对照则呈先上升后降低趋势,春伐和秋伐均降低(图1B)。K含量在不同生长阶段的变化范围在11.99~29.97 g/kg,平均值为(19.75±4.58)g/kg。K含量在火烧和春伐处理下,随着疏叶骆驼刺生长其显著降低,对照则是先下降后趋于稳定,而秋伐呈先上升后降低趋势(图1C)。N:P在不同生长阶段的变化范围在7.13~17.02,平均值为(13.00±2.25)。N:P中,秋伐为先降低后上升,而春伐则是先上升后趋于稳定,火烧处理下叶片N:P比呈缓慢下降趋势,至于对照则呈上升趋势(图1D)。
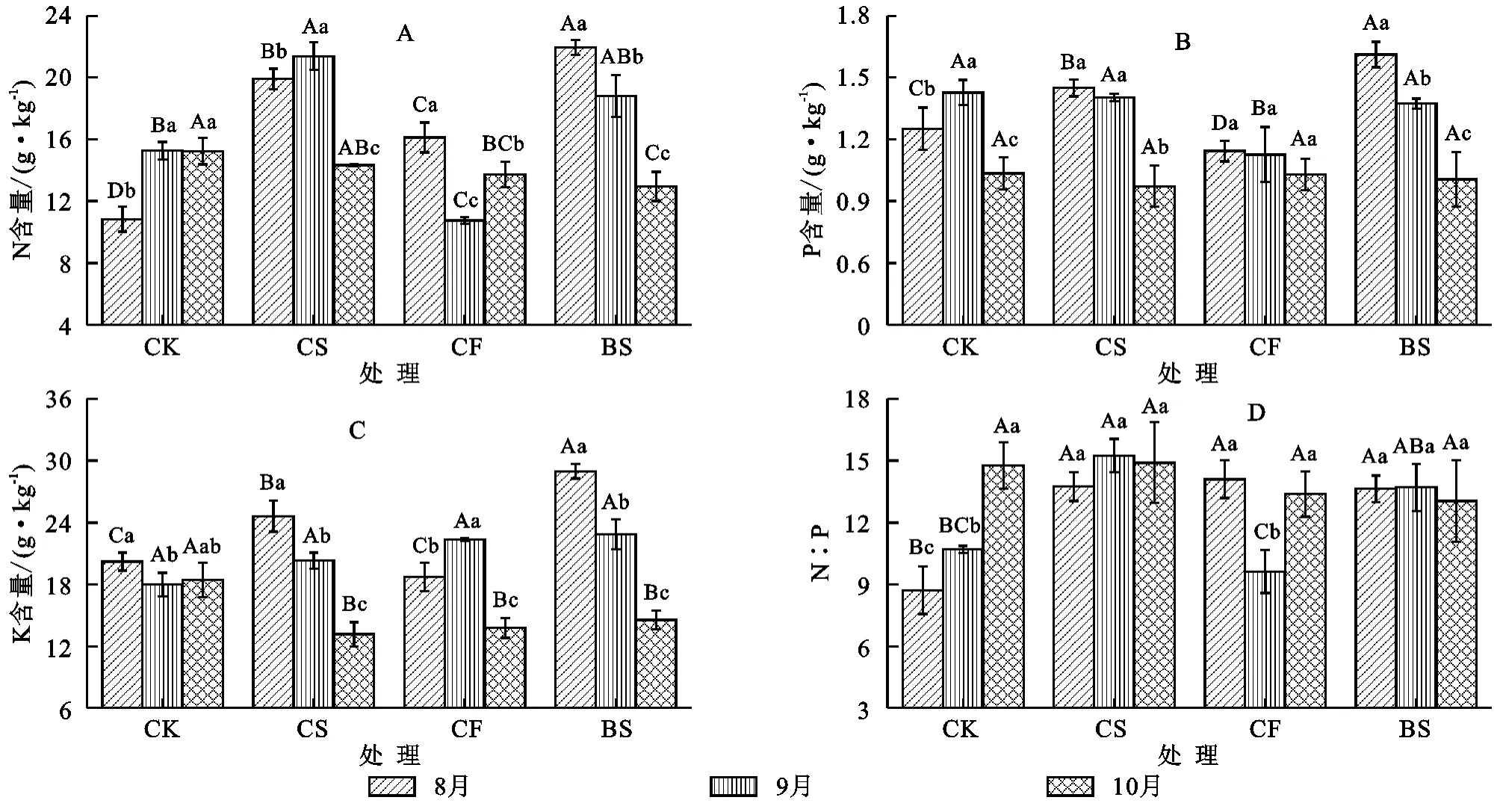
图1 不同干扰处理下疏叶骆驼刺叶片N,P,K及其N:P的动态变化特征(平均值±标准差)
2.2 不同干扰处理下疏叶骆驼刺叶片N,P,K含量及其计量比的变异特征
不同干扰下疏叶骆驼刺叶片N,P,K含量及其计量比的变异特征不同(表1)。疏叶骆驼刺叶片N含量的变异整体较大,其变异系数均大于10%,其中火烧处理下的变异系数最大,为22.32%。植物叶片P含量的变异,仅秋伐处理下的变异系数(8.64%)小于10%,其余3种处理均大于10%,火烧处理的变异系数最大,为20.43%。植物叶片K含量的变异,在对照下变异系数最小,为8.11%,而火烧的变异系数最大,为28.27%。疏叶骆驼刺叶片N:P含量的变异,春伐和火烧的变异系数均小于10%,分别为8.67%和9.47%,而对照的变异系数最大,为24.26%。
不同生长阶段疏叶骆驼刺叶片N,P,K含量及其计量比的变异特征也存在较大差异(表2)。植物叶片N含量中,仅10月份变异系数小于10%,其余月份均大于20%。而在P含量中,整体变异较小,仅8月份变异大于10%。植物叶片K及N:P中,不同月份变异系数均高于10%,K在8月份变异系数最大,为18.41%;而N:P在10月份变异系数最小,为11.58。
综合来看,4种干扰下疏叶骆驼刺叶片所有元素含量及其计量比变异系数均较高,变化范围为14.87%~20.4%,其中K含量变异系数最高为20.4%,N:P变异系数为14.87%。

表1 不同干扰处理下疏叶骆驼刺叶片N,P,K含量及其化学计量比的变化
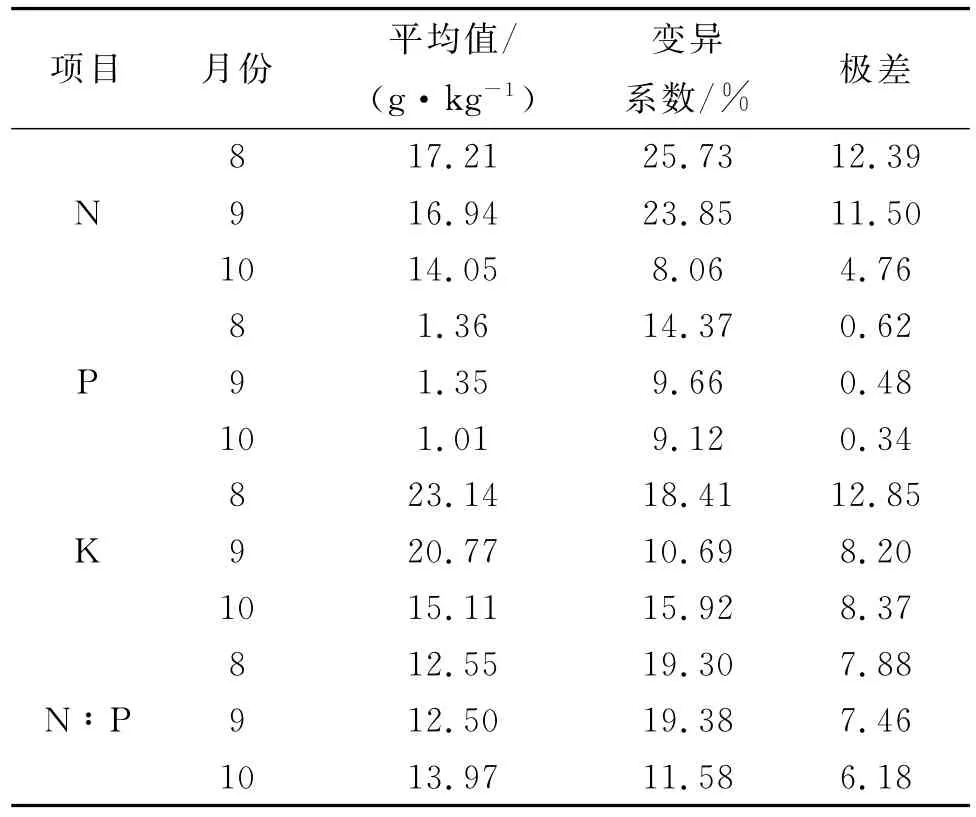
表2 不同生长阶段疏叶骆驼刺叶片N,P,K含量及其计量比的变化
2.3 不同干扰处理和季节变化对疏叶骆驼刺叶片N,P,K及其N:P的影响
疏叶骆驼刺叶片N,P,K及其N:P受不同的采样时间和不同干扰处理的单因素及其两因素的交互影响程度各不相同(表3)。疏叶骆驼刺叶片N含量主要受生长和处理的交互影响,其离差平方和达到376.96,但生长和处理对疏叶骆驼刺叶片N含量影响也达到显著性水平。疏叶骆驼刺叶片P含量主要受生长阶段的影响,其次为处理与生长的交互作用,但不同处理对叶片P含量的影响也达到显著性水平。与P含量相似,疏叶骆驼刺叶片K含量主要受生长阶段的影响,其离差平方和达到538.22,其次为处理与生长阶段的交互作用,但不同处理对叶片K含量的影响也达到显著性水平。疏叶骆驼刺叶片N:P则主要受生长与处理的交互作用,其次为处理,受生长阶段的影响最小,但它对叶片N:P的影响也达到显著性水平。
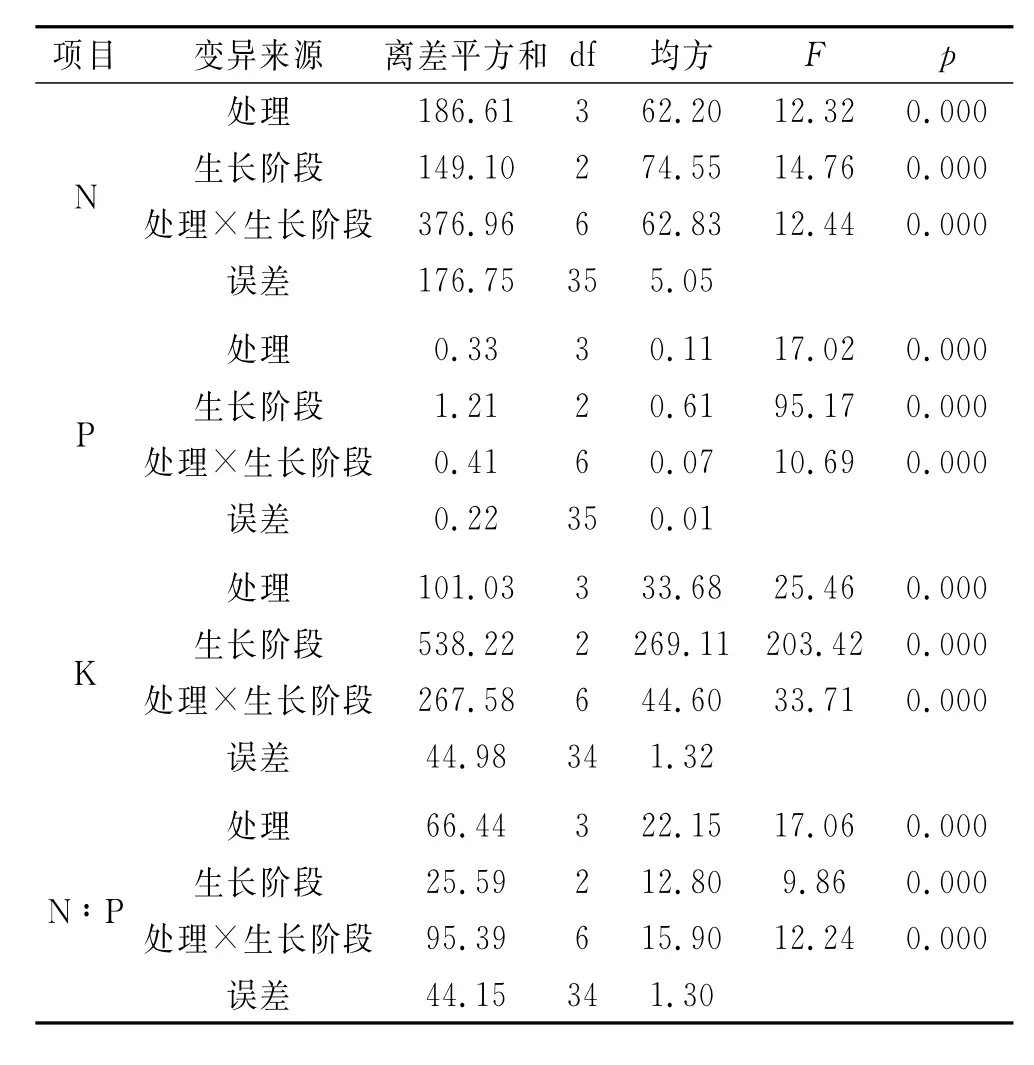
表3 不同干扰处理下疏叶骆驼刺叶片N,P,K含量及其化学计量比的变异来源分析
3 讨论
营养元素在植物体中的分布与动态变化特征,不仅反映了植物自身生物学特征,还受生长环境,如土壤、水分等方面的影响[14]。此外,植物对养分的需求量以及植物发育阶段等因素也对其有一定影响[15-16]。在本研究中,不同干扰处理下疏叶骆驼刺叶片在不同生长阶段的平均N含量为(16.09±3.75)g/kg,显著低于全球水平20.1 g/kg[17];平均 P含量为(1.24±0.22)g/kg,低于全球叶片P含量1.99 g/kg[18];而叶片平均N:P为13.00±2.25,也低于Han等[19]报道的结果。本文的研究结果中,除氮元素外,P及N:P的变化范围均落在李玉霖等[20]关于北方典型荒漠地区214种植物叶片氮磷分布特征的研究范围中。
有研究表明,植物叶片N,P,K等元素含量在不同的生长阶段差异很大[19,21]。本研究中,疏叶骆驼刺叶片N含量在对照条件下呈增加趋势,P含量是先升高后降低,K含量的变化幅度不大,而N:P比则是逐渐增大,这与Ågren[22]对陆地生态系统的N:P变化趋势的总结又略有不同,他认为在植物生长初期N:P逐渐降低,到生长旺季达到最低,生长末期又回升至初期,而后随叶片衰老,N:P逐渐降低。本研究结果显示,疏叶骆驼刺从生长旺季到衰老阶段叶片的N:P呈上升趋势,这可能与豆科植物特有的氮素吸收机制有关。
火烧和刈割作为生态系统管理的重要措施,可迅速改变养分循环[23]。本研究中,春伐、秋伐、火烧3种干扰处理下,叶片N含量整体呈降低趋势。尤其是火烧处理下,叶片N含量降低幅度最大。可能的原因是火烧影响了该区域土壤微生物群落[24],减少了土壤水分含量,尤其在荒漠地区,土壤有效养分的释放与转移受土壤水分影响较大,进而对骆驼刺植物养分含量的影响较大。也有研究发现火烧对植物对于光和养分的利用有一定影响[25]。但在本试验中,火烧是否对疏叶骆驼刺叶片的光合有影响,还需进一步研究。P含量整体呈下降趋势,这可能是因为,在8月份,温度较高,光照强烈,光合速率较高,而N,P作为蛋白质和RNA的组成元素[26-27],因而其含量较高。随着季节的变化,一方面,温度降低,导致其光合速率降低,因而N,P含量下降;另一方面,植物在生长季末的养分回收也是导致其降低的原因。
研究表明,N和P元素对植物的生长和发育起着非常重要的作用,而叶片N:P对群落结构常起到指示作用,它可作为判断环境对植物生长的养分供应状况的指标[28-32]。当叶片N:P<14时,植物生长主要受到N限制;当N:P>16时,植物生长主要受P限制;当14<N:P<16时,植物生长受N和P共同限制[33]。在本文中,研究区不同干扰下疏叶骆驼刺叶片N:P在不同生长阶段平均值分别为12.55,12.50,13.97,明显小于14,表明该区域疏叶骆驼刺生长可能主要受到N限制。这与一般植物生长多受P元素的限制,主要是因为环境为植物提供的可直接被吸收利用的P相对于N会更少有所不同[31]。但骆驼刺作为一种具有生物固氮能力的豆科植物,其生长的首要限制因素是否是N,还需要试验进一步验证。
我们可以通过植物叶片N,P含量以及N:P的变异来解释植物的遗传特性或对环境的适应结果。李玉霖等对北方荒漠地区植物叶片N,P含量及其计量比的研究中得出:植物叶片N含量的变化范围为6.8~52.1 mg/g,叶片P含量的变化范围为0.13~7.15 mg/g,N:P的变化范围为4.6~52.4[20]。综合分析本文4种干扰处理下疏叶骆驼刺叶片N,P,K含量及其计量比的季节变异发现,疏叶骆驼刺叶片N含量的变异系数最大,其次为K和P,叶片N:P的变异系数最小。本研究得出,不同干扰处理下的疏叶骆驼刺叶片N含量变化范围为10.01~22.40 g/kg,P含量变化范围为0.84~1.69 g/kg,N:P变化范围为7.13~17.02,均落在李玉霖等[20]对北方荒漠植物叶片氮磷化学计量的研究结果之内。其中P含量的变化范围又在牛得草等[34]在阿拉善荒漠区的研究结果之内,而N含量以及N:P的变化下限相对较低。针对单一处理下疏叶骆驼刺叶片N,P,K含量及其N:P计量比的季节变异分析显示,由于季节变异所计算的变异系数变化范围分别为16.29%~22.32%,8.64%~20.43%,8.11%~28.27%,8.67%~24.26%。作为细胞结构与功能最重要的生命元素,N,P,K的含量及其N:P随植物的生长发育而发生变化[35]。本研究中,P及K均主要受生长阶段的影响,其次为不同处理与生长季节的交互作用,受不同处理的影响最小;而N含量及N:P主要受处理与生长季节的交互作用影响,但不同处理和生长阶段对叶片N含量及N:P的影响也达到了显著性水平。
4 结论
疏叶骆驼刺作为策勒绿洲前沿风沙灾害的主要生态屏障,具有较大的生态防护效益和社会经济价值。砍伐和火烧作为当地疏叶骆驼刺主要的管理措施,对绿洲—过渡带的疏叶骆驼刺植被带来很大影响。虽然火烧和砍伐可以促进疏叶骆驼刺的更新以及生长,但是也会影响植物养分平衡,进而改变植物养分利用策略。本研究通过长期野外试验研究发现,春伐和火烧2种干扰处理下,不同季节叶片N,P和K含量整体呈降低趋势,尤其是火烧处理下,其含量降低幅度最大。相对于春伐而言,秋伐处理对疏叶骆驼刺叶片N,P和K含量的影响较小。此外,疏叶骆驼刺叶片N及N:P主要受生长季节与干扰的交互作用影响,而P及K含量主要受生长季节的影响。因此,为了更加合理地开发与利用疏叶骆驼刺,我们推荐秋伐作为疏叶骆驼刺植被主要的管理措施。
[1]Sardans J,Rivas-Ubach A,Penuelas J.The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function:a review and perspectives[J].Biogeochemistry,2012,111(1/3):1-39.
[2]张丽霞,白永飞,韩兴国.N:P化学计量学在生态学研究中的应用[J].植物学报:英文版,2003,45(9):1009-1018.
[3]高三平,李俊祥,徐明策,等.天童常绿阔叶林不同演替阶段常见种叶片N,P化学计量学特征[J].生态学报,2007,27(3):947-952.
[4]王绍强,于贵瑞.生态系统碳氮磷元素的生态化学计量学特征[J].生态学报,2008,28(8):3937-3947.
[5]马露莎,陈亚南,张向茹,等.黄土高原刺槐叶片生态化学计量学特征[J].水土保持研究,2014,21(3):57-61.
[6]王宁,张有利,王百田,等.山西省油松林生态系统碳氮磷化学计量特征[J].水土保持研究,2015,22(1):72-79.
[7]李小明,张希明.塔克拉玛干沙漠南缘自然植被的水分状况及其恢复[J].生态学报,2003,23(7):1449-1453.
[8]曾凡江,李向义,张希明,等.策勒绿洲多枝怪柳灌溉前后水分生理指标变化的初步研究[J].应用生态学报,2002,13(7):849-853.
[9]董志国,张苏江.干旱荒漠区骆驼刺资源利用[J].中国草地,2000(4):73-75.
[10]李海峰,曾凡江,桂东伟,等.不同干扰方式对疏叶骆驼刺形态特征及地上生物量的影响[J].应用生态学报,2012,23(1):23-28.
[11]高欢欢,曾凡江,鲁艳,等.不同干扰方式对疏叶骆驼刺生长及生理特征的影响[J].草业学报,2015,24(2):202-207.
[12]Bruelheide H,Vonlanthen B,Jandt U,et al.Life on the edge:to which degree does phreatic water sustain vegetation in the periphery of the Taklamakan Desert[J].Applied Vegetation Science,2010,13(1):56-71.
[13]鲍士旦.土壤农化分析[M].3版.北京:中国农业出版社,2005.
[14]刘长娥,杨永兴.九段沙芦苇湿地生态系统N,P,K的循环特征[J].生态学杂志,2008,27(3):418-424.
[15]He J S,Wang L,Flynn D F B,et al.Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes[J].Oecologia,2008,155(2):301-310.
[16]Coste S,Roggy J C,Imbert P,et al.Leaf photosynthetic traits of 14 tropical rain forest species in relation to leaf nitrogen concentration and shade tolerance[J].Tree Physiol,2005,25(9):1127-1137.
[17]Reich P B,Oleksyn J.Global patterns of plant leaf N and P in relation to temperature and latitude[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):11001-11006.
[18]Elser J J,Sterner R W,Gorokhova E,et al.Biological stoichiometry from genes to ecosystems[J].Ecology Letters,2000,3(6):540-550.
[19]Han W,Fang J,Guo D,et al.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J].New Phytologist,2005,168(2):377-385.
[20]李玉霖,毛伟,赵学勇,等.北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究[J].环境科学,2010,31(8):1716-1725.
[21]吴统贵,吴明,刘丽,等.杭州湾滨海湿地3种草本植物叶片N,P化学计量学的季节变化[J].植物生态学报,2010,34(1):23-28.
[22]Ågren G I.Stoichiometry and nutrition of plant growth in natural communities[J].Annual Review of Ecology,Evolution and Systematics,2008,39:153-170.
[23]刘超,王洋,王楠,等.陆地生态系统植被氮磷化学计量研究进展[J].植物生态学报,2012,36(11):1205-1216.
[24]杨文燕,宋长春,张金波.烧荒对土壤性质的影响[J].水土保持研究,2004,11(2):187-188.
[25]Cui Q,LüX T,Wang Q B,et al.Nitrogen fertilization and fire act independently on foliar stoichiometry in a temperate steppe[J].Plant and Soil,2010,334(1/2):209-219.
[26]Elser J J,Sterner R W,Gorokhova E,et al.Biological stoichiometry from genes to ecosystems[J].Ecology Letters,2000,3(6):540-550.
[27]Niklas K J,Owens T,Reich P B,et al.Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth[J].Ecology Letters,2005,8(6):636-642.
[28]Vitousek P.Nutrient cycling and nutrient use efficiency[J].American Naturalist,1982,119(4):553-572.
[29]Wassen M J,Olde Venterink H G M,Swart E O A M.Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems[J].Journal of Vegetation Science,1995,6(1):5-16.
[30]Aerts R,Chapin F S I.The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns[J].Advances in Ecological Research,1999,30(8):1-67.
[31]Güsewell S,Koerselman W.Variation in nitrogen and phosphorus concentrations of wetland plants[J].Perspectives in Plant Ecology Evolution and Systematics,2002,5(1):37-61.
[32]Güsewell S.N:P ratios in terrestrial plants:variation and functional significance[J].New Phytologist,2004,164(2):243-266.
[33]Aerts R.Interspecific competition in natural plant communities:mechanisms,trade-offs and plant-soil feedbacks[J].Journal of Experimental Botany,1999,50(330):29-37.
[34]牛得草,李茜,江世高,等.阿拉善荒漠区6种主要灌木植物叶片C:N:P化学计量比的季节变化[J].植物生态学报,2013,37(4):317-325.
[35]Kerkhoff A J,Enquist B J,Elser J J,et al.Plant allometry,stoichiometry and the temperature-dependence of primary productivity[J].Global Ecology and Biogeography,2005,14(6):585-598.
Effects of Burning and Cutting on Leaf Ecological Stoichiometry Characteristics of Alhagi sparsifolia
ZHAO Shenglong1,2,3,4,ZENG Fanjiang1,2,3,ZHANG Bo1,2,3,4,LI Changjun1,2,3,4,LIU Bo1,2,3,GAO Huanhuan1,2,3,4
(1.State Key Laboratory of Desert and Oasis Ecology,Xinjiang Institute of Ecology and Geography,Chinese Academy of Sciences,Urumqi830011,China;2.Cele National Station of Observation and Research for Desert-Grassland Ecosystems,Cele,Xinjiang848300,China;3.Key Laboratory of Biogeography and Bioresource in Arid Zone,Chinese Academy of Sciences,Urumqi830011,China;4.University of Chinese Academy of Sciences,Beijing100049,China)
Taking the dominant plantAlhagi sparsifoliaat the southern fringe of Taklimakan Desert as sample,we examined the effects of different disturbances treatments(cutting in spring,cutting in fall and burning in spring)on leaf N,P and K ecological stoichiometry characteristics.The results showed that N,P and K concentrations ofAlhagi sparsifolialeaf were significantly lower under cutting in spring and burning treatments along with the seasonal variation,while N:P mass ratio was no obvious variation.The N concentration and N:P mass ratio of leaf first decreased and then increased,and K concentration first increased then decreased under cutting in fall treatments,at the same time,P concentration was no obvious variation.Based on variation analysis among different seasons within different treatments ofAlhagisparsifolialeaf,N,P,K concentrations were more seasonal dynamic under the treatment of burning in spring than the other treatments,which were 22.32%,20.43%,28.27%,respectively,seasonal dynamics of N concentration and N:P mass ratio under treatment of cutting in spring were less than the other treatments,which were 16.29%,8.67%,respectively.P concentration under the treatment of cutting in fall was less seasonal dynamic than the other treatments,which was 8.64%.K concentration was less seasonal dynamic and in N:P mass ratio was more seasonal dynamics under control treatment than the other treatments,which were 8.11%,24.26%,respectively.With growth stage and treatments as independent factors,factorial analysis of variation for N,P and K concentrations and N:P mass ratio showed thatAlhagi sparsifolialeaf N concentration and N:P mass ratio were mainly determined by an interaction between growth stage and disturbance,while leaf P and K concentrations were mainly determined by growth stage of plant.
Alhagi sparsifolia;cutting;burning;N,P,K;dynamic variation
Q945.79
A
1005-3409(2017)01-0084-06
2015-11-05
2015-12-04
新疆自治区科技重大专项(201130106-1);国家基金委—新疆联合基金重点项目(U1203201);国家自然科学基金(41371516)
赵生龙(1989—),男,甘肃永登县人,硕士研究生,研究方向为荒漠植物生理生态。E-mail:13619398146@163.com
曾凡江(1966—),男,河南镇平人,研究员,博士生导师,主要从事荒漠植物生理生态研究。E-mail:zengfj@ms.xjb.ac.cn
猜你喜欢
计测技术(2020年6期)2020-06-09
阅读与作文(小学高年级版)(2020年2期)2020-05-21
南方周末(2019-06-27)2019-06-27
特别健康(2018年4期)2018-07-03
小学生作文(低年级适用)(2018年12期)2018-03-23
领导文萃(2018年3期)2018-03-12
北方人(2018年3期)2018-03-08
消费导刊(2017年24期)2018-01-31
莫愁(2017年15期)2017-05-11
红蜻蜓·低年级(2016年4期)2016-11-19
