
不同寄主对黄胸蓟马Thrips hawaiiensis生长发育及产卵的影响
2017-12-22 05:32:12曹宇尚保珍冉光梅赵燕群李灿张润志
中山大学学报(自然科学版)(中英文) 2017年6期
曹宇,尚保珍,冉光梅,赵燕群,李灿,张润志
(1. 贵阳学院生物与环境工程学院∥贵州省珍稀动物与经济昆虫重点实验室,贵州 贵阳 550005;2. 中国科学院动物研究所∥中国科学院动物进化与系统学重点实验室,北京 100101)
不同寄主对黄胸蓟马Thrips hawaiiensis生长发育及产卵的影响
曹宇1,尚保珍1,冉光梅1,赵燕群1,李灿1,张润志2
(1. 贵阳学院生物与环境工程学院∥贵州省珍稀动物与经济昆虫重点实验室,贵州 贵阳 550005;2. 中国科学院动物研究所∥中国科学院动物进化与系统学重点实验室,北京 100101)
黄胸蓟马Thripshawaiiensis是一种亚洲地区广泛分布的重要栖花害虫。研究了其在玫瑰、郁金香、康乃馨和百合等4种花卉寄主上的生长发育、存活及产卵差异,以期为探讨黄胸蓟马对不同寄主的为害差异及机制提供基础数据。结果表明,黄胸蓟马在玫瑰上发育最快,其完成一代需要10.07 d;在百合上发育最慢,完成一代需11.22 d。黄胸蓟马的世代存活率在郁金香上最高(0.76),在康乃馨上最低(0.64)。不同短时产卵处理表明,黄胸蓟马最喜好在郁金香上产卵,最不喜好在百合上产卵。因此,根据黄胸蓟马的生长发育、存活及产卵表现,4种寄主中,郁金香是黄胸蓟马的最适宜寄主,百合是其最不适宜寄主。
黄胸蓟马Thripshawaiiensis;花卉寄主;生长发育;存活;产卵
黄胸蓟马Thripshawaiiensiss(Morgan, 1913),隶属于缨翅目Thysanoptera锯尾亚目Terebrantia蓟马科Thripidae,是一种重要的栖花害虫。黄胸蓟马起源于环太平洋地区[1],主要分布于亚洲热带、亚热带和北美南部,是一种危险性害虫[2]。近年来,在法国[3]、西班牙[4]和土耳其[5]等欧洲地区也出现为害报道。在我国主要分布于海南、广西、广东、云南、福建和台湾等省区,是香蕉、蔬菜等植物花期的常见害虫[6-8]。
黄胸蓟马在贵州省蔬菜、花卉寄主花期均有为害[9-10],且表现出不同的嗜食性,但目前关于黄胸蓟马与寄主植物相互关系的研究较少。花卉产业在贵州省具有重要的经济地位,本文以玫瑰Rosarugosa、郁金香Tulipagesneriana、康乃馨Dianthuscaryophyllus和百合Liliumbrowniivar.viridulum等4种花卉寄主的花瓣为饲养材料,研究了黄胸蓟马在4种花卉寄主上的生长发育、存活及产卵差异,以期在一定程度上探讨其对不同寄主的为害差异,为花卉上黄胸蓟马的防治提供基础数据,保护花卉产业的健康发展。
1 材料与方法
1.1 寄主植物
玫瑰、郁金香、康乃馨和百合采自贵阳学院生物与环境工程学院园林专业的苗圃基地。寄主植物种植于温室大棚,生长期间无虫为害,且未喷洒农药,取其花蕾期的花瓣用于实验。
1.2 供试虫源
黄胸蓟马采自贵阳学院各类植物上,将其带回实验室置于塑料养虫盒内,分别以玫瑰、郁金香、康乃馨和百合4种寄主的花瓣为食料,在温度为(25 ± 1)℃、相对湿度(70 ± 5) %、光照L∶D=14∶10)的人工气候箱(RXZ系列多段可编程智能人工气候箱,RTOP-Y系列)中饲养3代后备用。
1.3 方 法
1.3.1 黄胸蓟马的生长发育及存活观察 将若干黄胸蓟马成虫(雌∶雄=1∶1)分别置于食料为玫瑰、郁金香、康乃馨和百合4种寄主的花的养虫盒内,每种寄主2盒,每盒放入黄胸蓟马200头(雌∶雄=1∶1),任其产卵,24 h 后移去成虫,将养虫盒置于人工培养箱中直到孵化出若虫,饲养条件同1.2。参考张治军等方法[11],挑取黄胸蓟马1龄若虫单头饲养于直径4.0 cm小塑料培养皿,皿内装有润湿滤纸片和直径约3.5 cm2的寄主花1片,每种寄主饲养100头,每头若虫为1个重复,每日8:00和19:00时观察 2 次,记录各虫态的发育和存活情况,直到若虫死亡或发育为成虫,卵的历期记录为从成虫产卵至卵孵出若虫的时间。每天更换新鲜植物的花瓣。
1.3.2 黄胸蓟马的产卵量观察 分别取玫瑰和康乃馨的花10~15片,由于郁金香和百合花瓣较大,取5~6片即可。将4种花瓣分别放入塑料瓶(规格: 高10 cm,直径为4 cm) 内,每个瓶各接入羽化3 d后的黄胸蓟马成虫20头(雌∶雄=1∶1),以200目纱布封住瓶口,防治蓟马逃逸,然后放入气候箱中任其产卵,在其分别产卵12,24,36和48 h后移除昆虫,各产卵处理重复4次,将含有虫卵的花瓣放入底部铺置湿润滤纸片的培养皿中(直径4 cm),皿口用Parafilm膜封住,并刺数小孔,保持通气,直至卵全部孵化成若虫[11-12],以一龄若虫作为黄胸蓟马产卵量的衡量[13]。实验条件同1.2。
1.4 数据统计与分析
采用SPSS18.0软件处理所测数据,采用Tukey’s多重比较检验,比较不同寄主间的差异显著性(P< 0.05)。
2 结果与分析
黄胸蓟马各虫态在不同花卉寄主上具有不同的差异显著性(表1)。卵期在4种花瓣上无显著差异。一龄若虫在玫瑰上发育最快(1.81 d),与郁金香无显著差异;在百合上发育最慢(2.05 d),与郁金香和康乃馨均无显著差异。二龄若虫在不同花瓣上的发育快慢与一龄若虫相似。预蛹在玫瑰上发育最快(1.46 d),与郁金香、康乃馨无显著差异;在百合上发育最慢(1.63 d),也与郁金香、康乃馨无显著差异。寄主对黄胸蓟马蛹和整个未成熟期(卵—成虫)的影响相似,其在玫瑰上的发育最快,分别为2.13 和 10.07 d;在百合上最慢,分别为2.55 和11.22 d;两者在郁金香和康乃馨之间均无显著差异。
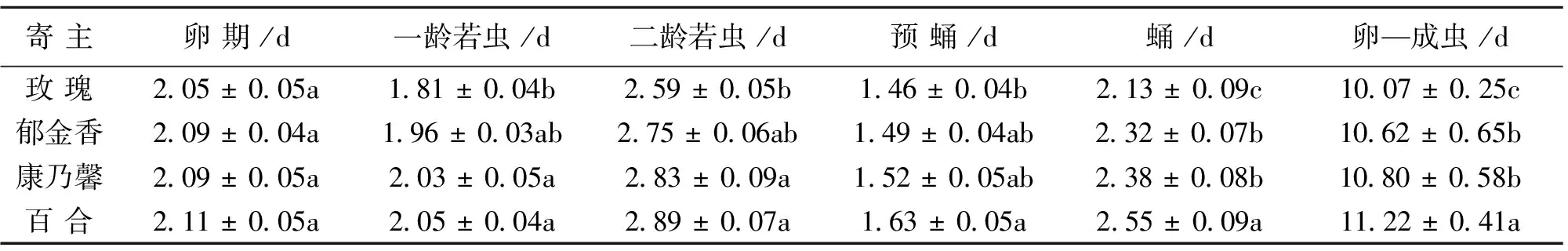
表1 不同虫态黄胸蓟马在不同寄主上的发育历期1)表中数据为平均数±标准误。同行不同小写字母表示不同寄主花上黄胸蓟马的生长发育差异达显著水平(P<0.05; Tukey’s多重比较检验)。Table 1 Development period of Thrips hawaiiensis at different stages on flowers of different hosts
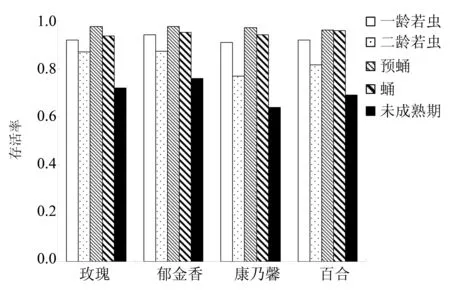
图1 不同虫态黄胸蓟马在不同寄主上的存活率Fig.1 Survival rate of Thrips hawaiiensis at different stages on flowers of different hosts
不同花瓣上,黄胸蓟马不同虫态的存活率同样呈现不同差异(图 1)。一龄若虫的存活率在康乃馨上最低,为0.91;在郁金香上最高,为0.94。二龄若虫的存活率在康乃馨上最低,为0.77;在玫瑰和郁金香上最高,均为0.87。预蛹及蛹期在4种花瓣上的存活率较高,均超过90%。整个未熟期的存活率在康乃馨上最低(0.64),在郁金香上最高(0.76)。另外,同一花瓣上,黄胸蓟马不同龄期的存活率也表现出不同差异。因此,黄胸蓟马相同龄期在不同寄主上的存活率、不同龄期在相同寄主上的存活率,均受寄主影响。
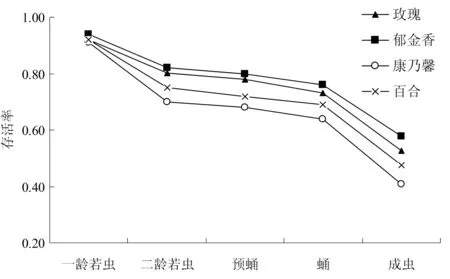
图2 不同虫态黄胸蓟马在不同寄主上的累积存活曲线Fig. 2 Cumulative survival curve of Thrips hawaiiensis at different stages on different flower hosts
由于黄胸蓟马产卵于花瓣组织中,无法统计其存活率,因此假设卵均能孵化为若虫,其孵化率为100%,以此计算黄胸蓟马整个未成熟期的累积存活情况[14]。如图2所示,不同花卉寄主上,黄胸蓟马各虫态的累积存活率均逐渐下降,其在康乃馨上表现最为剧烈,到羽化时累积存活率仅为0.41;在郁金香上各虫态的累积存活率最高,到羽化时为0.58。4种寄主上,黄胸蓟马在预蛹和蛹期阶段,曲线平缓,说明此时期死亡率较低。因此,寄主引起蓟马死亡的时期,主要集中于若虫和羽化阶段。
不同产卵时间处理下,黄胸蓟马在4种花卉寄主上的产卵量存在不同程度的差异(表 2)。产卵12 h处理下,黄胸蓟马的总产卵量在4种寄主之间无显著差异,但平均产卵量在郁金香上最高(4.25),在百合上最低(3.15)。产卵24 h处理下,其总产卵量在郁金香(47.50)和康乃馨上最高(43.75),两者显著高于玫瑰(40.50)和百合(37.50);平均产卵量则在郁金香上最高(4.75),在玫瑰上最低(3.75)。产卵36 h处理下,总产卵量同样在郁金香上最高(70.25),但与康乃馨(63.75)无显著差异;在玫瑰(55.00)上最低,但与百合(61.50)无显著差异;平均产卵量在4种寄主间的显著性差异与总产卵量类似。产卵48 h处理下,总产卵量及平均产卵量在4种寄主间的显著性差异与36 h处理下相似,其总产卵量、平均产卵量均在郁金香上最高,分别为94.50和4.73;在玫瑰上最低,分别为75.50和3.78。
相同寄主上,随着产卵时间的延长,其总产卵量显著增加,因此,在4种寄主上,不同处理下黄胸蓟马的总产卵量均为48 h > 36 h > 24 h > 12 h。相同寄主上,黄胸蓟马的平均产卵量受产卵时间的影响较小,如在玫瑰上,其平均产卵量在4种产卵时间处理之间无显著差异。而在郁金香、康乃馨和百合上,其平均产卵量均在24 h、36 h和48 h处理之间无显著差异,但三者处理下的平均产卵量显著高于12 h处理。
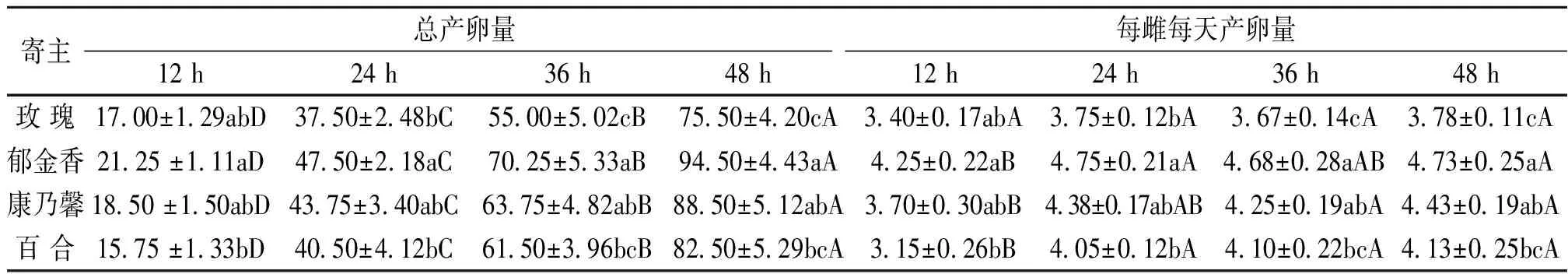
表2 不同处理下黄胸蓟马在不同寄主上的产卵量1)表中数据为平均数±标准误。同列中不同小写字母表示相同处理下,不同寄主之间差异显著;同行中不同大写字母表示相同寄主上,不同处理之间差异显著(P<0.05; Tukey’s多重比较检验)。Table 2 Oviposition of Thrips hawaiiensis on different flower hosts under different treatment
3 讨 论
昆虫在不同寄主上的生长发育、存活及产卵等差异,可从不同侧面反映其对寄主的偏好性[15-16]。本研究中,从未成熟期来看,黄胸蓟马对玫瑰的偏好性最强;从存活率来看,其对郁金香的偏好性最强;从产卵量来看,其对郁金香和康乃馨的偏好性最强。相关研究表明,相同实验温度下,在茶花粉上,黄胸蓟马的发育历期为8.90 d[2],低于本文中的所有结果;世代存活率为82.70%,高于本文中的所有寄主。同样,相同温度下的香蕉花上,其发育历期为10.61 d[17],与本文中郁金香上的发育历期相当,高于玫瑰,低于康乃馨和百合;存活率为63.16%,与康乃馨相当,低于其它3种花卉寄主。因此,寄主种类对黄胸蓟马的生长发育及存活存在显著影响。关于黄胸蓟马的产卵研究,报道较少。Cheng等的研究表明,25 ℃下,其一生在油桐花上的产卵量为41.9[18];相同温度下,其在茶花粉上的一生的产卵量为203.5,平均每天每雌产卵量为7.32[2],其平均产卵量显著高于本文中的研究结果。但本文仅研究了黄胸蓟马短时处理下的产卵比较,关于不同花卉寄主上其一生的繁殖力比较,需进一步探讨。综合生长发育、存活率及短时产卵量来看,本研究4种寄主中,郁金香是黄胸蓟马的最适宜寄主,而百合则是其最不适宜寄主。
除寄主外,温度对黄胸蓟马的生长发育、存活等也存在显著影响。如茶花粉上,在10~30 ℃之间,随着温度的升高,其发育历期显著缩短(52.1~ 7 d);20 ℃时,其世代存活率最高(96.7%),10 ℃下的存活率最低(25%)[2]。香蕉花上,在14~30 ℃之间,其发育历期最短为6.19 d,最长为28.54 d[17],存活率介于46.34%~63.16%之间;但卢辉等发现在20~36 ℃温度下,黄胸蓟马在香蕉花上的发育历期最短为9.68 d,最长为22.45 d,世代存活率最高为38.5%,最低为10.22%[19]。因此,同在香蕉花上的不同研究结果,可能与香蕉品种、蓟马地理种群差异有关,也可能与实验方法、实验条件等有关。而仅从温度对黄胸蓟马生长发育及存活等的影响来看,黄胸蓟马的适温范围较广,尤其在高温条件下,发育历期短,具备短时间内暴发成灾的特性[2, 17],需在生产和科研上加以重视。除寄主、温度外,湿度、天敌等因素都会影响蓟马的生存[20-21]。因此,要系统了解黄胸蓟马对不同花卉植物的偏好性及偏好机制,其需进一步研究自然条件下的生物与非生物因子作用下,黄胸蓟马在不同寄主上的生长发育、存活、繁殖等差异。
寄主理化性状对昆虫的寄主选择有着重要影响[22-23],如某些寄主特定的体表性状能够有效抵御美洲斑潜蝇的为害[24],而不同寄主的营养物质、次生物质差异等可显著影响斑潜蝇、蓟马等害虫的种群发展[25-26]。同样,取食不同寄主,可引起昆虫体内保护酶、解毒酶等发生不同变化,从而影响昆虫对寄主不同的适应性[27-28]。因此,要深入探讨黄胸蓟马对不同花卉寄主的偏好性,需进一步从寄主—昆虫双方面加强生理生化测定。
致谢:衷心感谢吉林大学王军博士对黄胸蓟马标本的鉴定。
[1] MOUND L A, KIBBY G. Thysanoptera: an identification guide[M]. 2nd ed. Oxon: CAB International,1998.
[2] MURAI T. Development and reproductive capacity ofThripshawaiiensis(Thysanoptera: Thripidae) and its potential as a major pest[J]. Bulletin of Entomological Research, 2001, 91: 193-198.
[3] REYNAUD P, BALMÉS V, PIZZOL J.Thripshawaiiensis(Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips now established in Europe[J]. Bulletin OEPP, 2008, 1: 155-160.
[4] GOLDARAZENA A. First record ofThripshawaiiensis(Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips in Spain[J]. Bulletin OEPP, 2011, 41: 170-173.
[5] ATAKAN E, ÖLÇÜLIÜ M, PEHLIVAN S, et al. A new thrips species recorded in Turkey:Thripshawaiiensis(Morgan, 1913) (Thysanoptera: Thripidae)[J]. Türkiye Entomoloji Bülteni, 2015, 2: 77-84.
[6] 林明光,刘福秀,彭正强,等. 海南省香蕉作物害虫调查与鉴定[J]. 西南农业学报, 2009, 22(6): 1619-1622.
LIN M G, LIU F X, PENG Z Q, et al. Survey and identification of pest insects on banana crop in Hainan[J]. Southwest China Journal of Agricultural Sciences, 2009, 22(6): 1619-1622.
[7] 郭志祥,曾莉,番华彩,等. 云南香蕉害虫种类及发生危害调查[J]. 热带农业科学, 2012, 32(10): 42-45.
GUO Z X, ZEN L, FAN H C, et al. Investigation on the species of banana insect pests and their damages in Yunnan Province[J]. Chinese Journal of Tropical Agriculture, 2012, 32(10): 42-45.
[8] 王慧,薛建平,刘浪,等. 菊花种苗蓟马种类和西花蓟马种群季节动态[J]. 云南农业大学学报, 2014, 29(4): 494-499.
WANG H, XUE J P, LIU L, et al. Species of thrips and seasonal population dynamics of western flower thrips on chrysanthemum seedling[J]. Journal of Yunnan Agricultural University, 2014, 29(4): 494-499.
[9] 袁成明,郅军锐,李景柱,等. 贵州省蔬菜蓟马种类调查研究[J]. 中国植保导刊,2008, 28(7): 8-10.
YUAN C M, ZHI J R, LI J Z, et al. Investigation on the species of thrips in fields of vegetable in Guizhou Province[J]. China Plant Protection, 2008, 28(7): 8-10.
[10] 黄宇,陈劲松,郅军锐,等. 贵阳地区花卉蓟马种类调查研究[J]. 河南农业科学, 2009, 38(6): 93-96.
HUANG Y, CHEN J S, ZHI J R, et al. Investigation on thrips species on flowers in Guiyang[J]. Journal of Henan Agricultual Sciences, 2009, 38(6): 93-96.
[11] 张治军,张友军,徐宝云,等. 温度对西花蓟马生长发育、繁殖和种群增长的影响[J]. 昆虫学报, 2012, 55(10): 1168-1177.
ZHANG Z J, ZHANG Y J, XU B Y, et al. Effects of temperature on development, reproduction and population growth of the western flower thrips,Frankliniellaoccidentalis[J]. Acta Entomologica Sinica, 2012, 55(10): 1168-1177.
[12] 曹宇,刘燕,熊正利,等. 西花蓟马对不同花卉寄主的产卵选择性[J]. 植物保护学报, 2015, 42(5): 741-748.
CAO Y, LIU Y, XIONG Z L, et al. Oviposition preference of western flower thripsFrankliniellaoccidentalisto different horticultural host plants[J]. Journal of Plant Protection, 2015, 42(5): 741-748.
[13] KOGEL W J D, BALKEMA-BOOMSTRA A, HOEK M V D, et al. Resistance to western flower thrips in greenhouse cucumber: effect of leaf position and plant age on thrips reproduction[J]. Euphytica, 1997, 94(1): 63-67.
[14] 曹宇,刘燕,梁文琴,等. 西花蓟马生长发育及其与寄主花化学物质的关系[J]. 中国生态农业学报, 2015, 23(5): 634-641.
CAO Y, LIU Y, LIANG W Q, et al. Development ofFrankliniellaoccidentalison flowers of different horticultural hosts and relationship with flower compounds[J]. Chinese Journal of Eco-Agriculture, 2015, 23(5): 634-641.
[15] van LENTEREN J C, NOLDUS L P J J. Whitefly-plant relationships: behavioural and ecological aspects. In D. Gerling (ed.), Whiteflies: their bionomics, pest status and management. Intercept, Andover, UK. 1990.
[16] 安新城,郭强,胡琼波,等. B型烟粉虱对23种寄主植物适应度的评估和聚类分析[J]. 生态学报, 2011, 31(11): 3150-3155.
AN X C, GUO Q, HU Q B, et al. Performance evaluation of B biotype whitefly,Bemisiatabacion 23 host plants [J]. Acta Ecologica Sinica, 2011, 31(11): 3150-3155.
[17] 张帆,付步礼,刘奎,等. 温度对香蕉花蓟马发育和存活的影响[J]. 生态学报, 2014, 34(14): 3895-3899.
ZHANG F, FU B L, LIU K, et al. The effect of temperature on the development and survival ofThripshamaiiensis[J]. Acta Ecologica Sinica, 2014, 34(14): 3895-3899.
[18] CHENG M F. Studies on morphology, life cycle and bionomics of the flower thrips,Thripshawaiiensis(Morgan), on citrus[J]. National Chia-I Agricultural Institute Bulletin, 1985, 12: 43-55.
[19] 卢辉,徐雪莲,卢芙萍,等. 温度对黄胸蓟马生长发育的影响[J]. 中国农学通报, 2011, 27(21): 296-300.
LU H, XU X L, LU F P, et al. Effects of temperature on development and reproduction ofThripshamaiiensis[J]. Chinese Agricultural Science Bulletin, 2011, 27(21): 296-300.
[20] CHAISUEKUL C, RILEY D G. Host plant, temperature, and photoperiod effects on ovipositional preference ofFrankliniellaoccidentalisandFrankliniellafusca(Thysanoptera: Thripidae)[J]. Journal of Economic Entomology, 2005, 6: 2107-2113.
[21] REITE S R, YEARBY E L, FUNDERBURK J J, et al. Integrated management tactics forFrankliniellathrips(Thysanoptera: Thripidae) in field-grown pepper[J]. Journal of Economic Entomology, 2003, 4: 1201-1214.
[22] 钦俊德,王琛柱. 论昆虫与植物的相互作用和进化的关系[J]. 昆虫学报, 2001, 44(3): 360-365.
QIN J D, WANG C Z. The relation of interaction between insects and plants to evolution[J]. Acta Ecologica Sinica, 2001, 44(3): 360-365.
[23] 庞保平,鲍祖胜,周晓榕,等. 寄主挥发物、叶色和表皮毛在美洲斑潜蝇寄主选择中的作用[J]. 生态学报, 2004, 24(3): 547-551.
PANG B P, BAO Z S, ZHOU X R, et al. Effects of host volatiles, leaf color, and cuticular trichomes on host selection byLiriomyzasativaeBlanchard[J]. Acta Ecologica Sinica, 2004, 24(3): 547-551.
[24] 成卫宁,仵均祥,李修炼,等. 美洲斑潜蝇寄主抗虫性与寄主叶片化学物质和物理结构的关系[J]. 中山大学学报(自然科学版), 2006, 45(5): 71-75.
CHENG W L, WU J X, LI X L, et al. Influence of chemicals and physical structure characteristics of host plant leaves on resistance toLiriomyzasativaeBlanchard[J]. Acta Scientlarum Naturalium Universitatis Sunyatseni, 2006, 45(5): 71-75.
[25] SCOTT B A S, SIMMONDS S J M, BLANEY W M. Relationship between nutritional composition of plant species and infestation levels of thrips[J]. Journal of Chemical Ecology, 2002, 28(12): 2399-2409.
[26] 闫丽英,庞保平,周晓榕,等. 南美斑潜蝇对不同菜豆品种的偏好性与寄主化学物质含量的关系[J]. 中国农业科学, 2008, 41(3): 713-719.
YAN L Y,PANG B P, ZHOU X R, et al. Relationship between host plant preference ofLiriomyzahuidobrensisfor differentPhaseolusvulgarisvarieties and plant compound contents[J]. Scientia Agricultura Sinica, 2008, 41(3): 713-719.
[27] 周福才,李传明,周桂生,等. 烟粉虱体内几种抗性酶对寄主转换的响应[J]. 生态学报, 2010, 30(7): 1806-1811.
ZHOU F C, LI C M, ZHOU G S, et al. Responses of detoxification enzymes inBemisiatabaci(Gennadius) to host shift[J]. Acta Ecologica Sinica, 2010, 30(7): 1806-1811.
[28] 张帆,马德英,周泓,等. 两种棉花防御反应次生代谢物质及其对烟粉虱羧酸酯酶活性的影响[J]. 应用昆虫学报, 2012, 49(2): 408-413.
ZHANG F, MA D Y, ZHOU H, et al. Two secondary defensive cotton metabolites and their effects on the specific activities of carboxylesterase inBemisiatabaci[J]. Chinese Journal of Applied Entomology, 2012, 49(2): 408-413.
InfluenceofdifferenthostsonthedevelopmentandovipositionofThripshawaiiensis
CAOYu,SHANGBaozhen,RANGuangmei,ZHAOYanqun,LICan,ZHANGRunzhi
(1. Department of Biology and Engineering of Environment, Key & Special Laboratory of Guizhou High College for Pest Control and Resource Utilization, Guiyang University, Guiyang 550005, China;2. Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China)
Thripshawaiiensisis an important flower inhabiting thrip pest, widely distributed in the Asian regions. The development, survival and oviposition ofT.hawaiiensison four flower hosts (Rosarugosa,Tulipagesneriana,DianthuscaryophyllusandLiliumbrowniivar.viridulum) were studied in this paper, in order to discuss the damage difference and damage mechanism to different plant hosts. The results showed that the shortest developmental period ofT.hawaiiensiswas onRosarugosaflower, which needed 10.07 d to complete one generation. The longest was onLiliumbrowniivar.viridulumflower, which needed 11.22 d to complete one generation. The highest survival rate was onT.gesnerianaflower, while the lowest was onD.caryophyllusflower, with the values of 0.76 and 0.64, respectively. According to different treatments for oviposition in short time, we found thatT.gesnerianaflower was the most preferred host for oviposition byT.hawaiiensis, whileLiliumbrowniivar.viridulumflower was the least preferred host. In summary, according to the performance of development, survival and oviposition ofT.hawaiiensison the four flower hosts,T.gesnerianawas the most suitable host forT.hawaiiensis, whileL.browniivar.viridulumwas the least.
Thripshawaiiensis;flower plant host;development;survival;oviposition
10.13471/j.cnki.acta.snus.2017.06.022
2016-12-14
贵州省省级特色重点学科项目(ZDXK[2015]11号); 贵州省高层次创新人才培养项目 (黔科合人才 (4020)号); 2016贵州省大学生创新创业计划训练项目(201610976010)
曹宇 (1984年生),男;研究方向昆虫生态与综合治理; E-mail:yucaosuccess@126.com
李灿(1979年生),男;研究方向有害生物控制与资源利用;E-mail: lican790108@163.com
张润志(1965年生),男;研究方向:外来入侵昆虫的鉴定、预警与综合治理;E-mail:zhangrz@ioz.ac.cn
Q965
A
0529-6579(2017)06-0141-06
猜你喜欢
今日农业(2021年11期)2021-11-27 10:47:17
中老年保健(2021年4期)2021-08-22 07:10:42
时代英语·高一(2019年5期)2019-09-03 02:09:34
娃娃乐园·综合智能(2018年9期)2018-06-08 05:49:20
现代园艺(2017年21期)2018-01-03 06:42:17
快乐语文(2016年15期)2016-11-07 09:46:34
好孩子画报(2014年11期)2014-11-12 03:26:21
高中生学习·高三版(2014年3期)2014-04-29 06:13:52
新疆农垦科技(2014年9期)2014-02-28 19:20:58
植物保护(2010年4期)2010-06-12 02:44:04
