
两种弧菌对菲律宾蛤仔代谢途径影响的比较研究
2017-12-18 08:39:58刘小莉侯承宗来涌凯尚秋爽邓小玉邱文彬孙虎山
水产科学 2017年2期
刘小莉,侯承宗,来涌凯,高 军,李 洁,尚秋爽,龙 静,邓小玉,邱文彬,孙虎山
( 鲁东大学 生命科学学院,山东 烟台 264025 )
两种弧菌对菲律宾蛤仔代谢途径影响的比较研究
刘小莉,侯承宗,来涌凯,高 军,李 洁,尚秋爽,龙 静,邓小玉,邱文彬,孙虎山
( 鲁东大学 生命科学学院,山东 烟台 264025 )
采用基于核磁共振波谱技术的代谢组学技术,探索菲律宾蛤仔在受到鳗弧菌和灿烂弧菌感染的代谢物变化特征,构建菲律宾蛤仔对弧菌感染后的代谢网络调控图谱,并比较两种弧菌毒性效应的差异。试验结果表明,3种代谢物葡萄糖、谷氨酸、苏氨酸在两种弧菌感染时均发生了变化,表征鳗弧菌感染的代谢物为牛磺酸、精氨酸、酪氨酸、苯丙氨酸、赖氨酸、亮氨酸、异亮氨酸、缬氨酸;而表征灿烂弧菌污染的代谢物为甜菜碱、二甲基甘氨酸、胆碱、谷氨酸、亚牛磺酸。
鳗弧菌;灿烂弧菌;菲律宾蛤仔;代谢物;核磁共振波谱技术
随着我国海洋高密度养殖技术的不断发展,各种海洋致病菌日益频繁出现,海水养殖动物弧菌(Vibrio)病的爆发成为威胁养殖业的主要瓶颈之一[1]。弧菌病是由弧菌属细菌引起的一类细菌性疾病,弧菌分泌的外毒素等物质,可导致多种海洋生物出现出血性败血症,并伴随大面积组织损伤而死亡[2]。其中,鳗弧菌(V.anguillarum)和灿烂弧菌(V.splendidus)是我国海水养殖动物所面临的重要条件致病菌,由此引起的水产养殖动物疾病较为严重,威胁着经济贝类的养殖安全[3-4]。
鳗弧菌和灿烂弧菌是常见的革兰氏阴性细菌,广泛存在于海水及近岸河口海区、沉积物、海洋生物体上等,可感染多种海水经济养殖动物[5]。无脊椎动物由于缺少真正的淋巴细胞和功能性抗体,主要依靠非特异性免疫功能来对抗病原菌[6-7]。鳗弧菌可引起贝类、虾类、鱼类等体内的免疫功能发生变化,多种基因表达异常,引起代谢紊乱[8-11]。灿烂弧菌引起水产动物体内的蛋白质发生变化,其中33%与免疫反应有关[12-15]。加强海洋贝类(宿主)与弧菌之间相互作用的分子应答机制研究,对预防与控制养殖贝类弧菌病的发生,具有重要的意义。
菲律宾蛤仔(Ruditapesphilippinarum)广泛分布于沿海区域,在渤海尤其是胶州湾水域分布甚广[16-17]。在菲律宾蛤仔的生长过程中,由于受环境因素及自身遗传因素的影响,同一群体中产生了壳色多态性,经过前期调研,在渤海地区菲律宾蛤仔的优势品系主要包括斑马蛤、白蛤及两道红等。已有的研究表明,不同壳色菲律宾蛤仔的生长和抗逆性状差异显著,免疫指标表现出较大差别[18];不同品系菲律宾蛤仔(白蛤及斑马蛤)对致病菌的响应研究中发现,白蛤反应更为敏感[19]。
代谢组学能在特定时间及环境中,对生物组织细胞所有低分子量代谢产物进行分析,其目的在于定性和定量生物体内各种代谢物,阐明代谢物的变化规律,从本质上揭示生命现象的本质和过程[20-21]。代谢组学分析技术因能从全局的角度评价生物体系对外界刺激的应答,故可应用于评价生物体对致病菌响应后代谢物及代谢通路的变化。
本试验以渤海区域常见的海洋养殖经济贝类菲律宾蛤仔(白蛤品系)为研究对象,采用基于核磁共振波谱技术的代谢组学技术,探索菲律宾蛤仔在受到鳗弧菌及灿烂弧菌感染后的代谢物变化特征,比较并构建菲律宾蛤仔对弧菌感染后的代谢网络调控图谱,从代谢水平解析菲律宾蛤仔对弧菌特异性应答效应及其分子机制,以期对菲律宾蛤仔抗弧菌感染的分子机理提供一定的理论依据。
1 材料与方法
1.1 染菌试验
菲律宾蛤仔常规适应性养殖9 d后,选取壳长相近的个体进行正式试验。试验分为正常对照组、鳗弧菌染菌组、灿烂弧菌染菌组,每组10枚。参照文献并在充分预试验的基础上进行菌液密度设定,鳗弧菌和灿烂弧菌的菌液密度皆调至1×107cfu/mL[22]。染菌24 h后迅速取消化腺组织冻存于液氮中以备后续试验。
1.2 试验所需主要试剂
重水(D2O,Sigma公司,151882),3-trimethylsilyl-2,2,3,3-d4-propionate(TSP,NORELL公司,5715),甲醇(Buidick & Jackson公司,13CG4H),氯仿(国药集团化学试剂有限公司,10006828),乙腈(国药集团化学试剂有限公司,40064160),重水磷酸盐缓冲液(100 mmol/L Na2HPO4和NaH2PO4,含0.5 mmol/L TSP,pH 7.0)。
1.3 试验所需主要仪器
核磁共振波谱仪(Bruker BioSpin公司,AV 500型),台式高速冷冻离心机(Eppendorf公司,5804型),显微镜成像提升装置(烟台显微生物科技有限公司,201620092090.6),干燥器(Thermo公司,UVS 400A型及SPD111V型)。
1.4 代谢物提取
取菲律宾蛤仔消化腺组织约100 mg于低温条件下研磨成细粉,加入甲醇/氯仿/水(体积比为1∶2∶1)溶剂体系对代谢物进行提取,充分涡旋混匀,然后离心(10 min,2800 r/min,4 ℃),离心后转移上层水相至新的1.5 mL离心管中,放入干燥器中充分干燥后以备核磁检测。
1.5 核磁共振波谱仪检测、数据分析及谱图解析
将干燥的样品粉末加入600 μL 100 mmol/L的重水磷酸盐缓冲液(pH=7.0),充分混匀溶解后离心(5 min,2800 r/min,4 ℃),取出上层550 μL液体转移至5 mm核磁检测管中进行核磁共振波谱仪检测。在Bruker AV 500氢核磁共振波谱仪上对白蛤消化腺样品进行分析,检测的具体参数如下:一维氢谱观察频率:500.18 MHz,测试温度:298 K,谱宽:6009.6 Hz,混合时间:0.1 s,弛豫延迟时间:0.3 s,在弛豫延迟和混合期间采用预饱和方式压制水峰。累加次数:128次,每个样本核磁扫描均收集到16384个数据点,采用线宽为0.3 Hz的指数窗函数进行傅里叶变换,获得氢核磁共振谱图载入TopSpin 软件(version 2.1,Bruker BioSpin,Canada),对所有的氢核磁共振谱图进行相位校正、基线调整,并以TSP内标为化学位移参考峰(TSP=0.0 ppm)。利用Matlab ProMetab软件(ver 7.0;MathsWorks,Natick,MA,USA),将δ 0.2~10.0内的谱图以0.005谱宽进行分段并积分,去除水峰所在的化学位移区域(4.72~4.96)。归一化剩余总的谱峰面积以便比较谱峰之间的强度差异。为了增加低强度峰的权重,在多元统计分析之前,先将所有的核磁谱进行对数化处理,进行log转换(转换因子,λ=6.36×108)。通过Chenomx Suite软件(评估版,Chenomx公司,加拿大)分析代谢物的化学位移,来完成主要代谢产物的鉴定。
2 结 果
2.1 菲律宾蛤仔代谢图谱解析
菲律宾蛤仔消化腺代谢物核磁共振一维氢谱见图1。由图1可知,共得到32种代谢物。
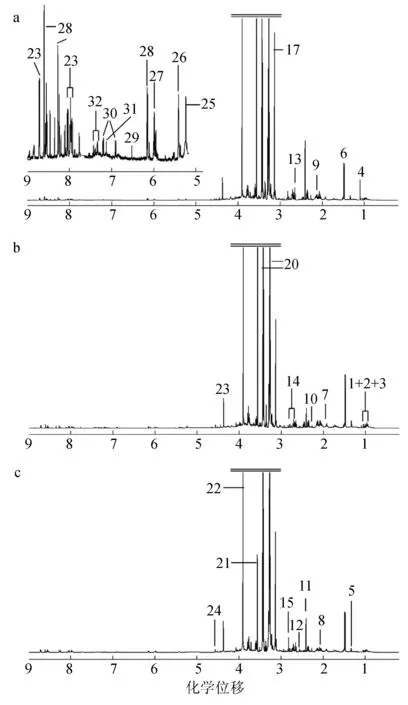
图1 正常对照组、鳗弧菌感染组、灿烂弧菌感染组菲律宾蛤仔消化腺代谢物核磁共振一维氢谱图
a.正常对照组图谱;b. 鳗弧菌感染后图谱;c. 灿烂弧菌感染后图谱;解析的代谢物分别为:1. 亮氨酸,2. 异亮氨酸,3. 缬氨酸,4. 未知代谢物(1.12×10-6),5. 苏氨酸,6. 丙氨酸,7. 精氨酸,8. 谷氨酸,9. 谷氨酰胺,10. 乙酰乙酸,11. 琥珀酸,12. β-丙氨酸,13. 亚牛磺酸,14. 天冬氨酸,15. 二甲基甘氨酸,16. 赖氨酸,17. 丙二酸,18. 胆碱,19. 磷酸胆碱,20. 牛磺酸,21. 甘氨酸,22. 甜菜碱,23. 龙虾肌碱,24. α-葡萄糖,25. β-葡萄糖,26. 糖原,27. 未知代谢物(5.98×10-6),28. ATP,29. 延胡索酸,30. 络氨酸,31. 组氨酸,32. 苯丙氨酸.
2.2 鳗弧菌和灿烂弧菌刺激后菲律宾蛤仔代谢物变化情况
菲律宾蛤仔感染两种弧菌后,代谢物变化不同,鳗弧菌胁迫组有11个代谢物发生变化,而灿烂弧菌胁迫组有8个代谢物发生变化,有3个代谢物葡萄糖、谷氨酸、苏氨酸在两种菌胁迫后都发生了变化(图2)。
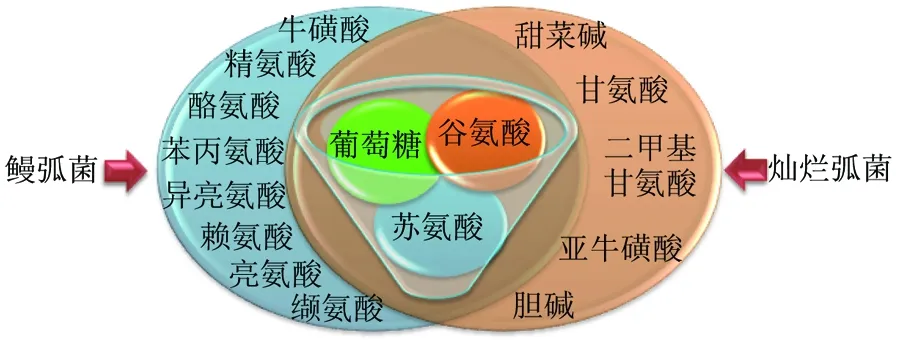
图2 菲律宾蛤仔受到鳗弧菌和灿烂弧菌感染后代谢物变化情况
2.3 鳗弧菌刺激后菲律宾蛤仔消化腺代谢途径变化
菲律宾蛤仔感染鳗弧菌后,代谢图谱变化见图3。由图3可知,降低的代谢物只有牛磺酸,升高的代谢物包括精氨酸、酪氨酸、苯丙氨酸、谷氨酸、葡萄糖、赖氨酸、亮氨酸、异亮氨酸、缬氨酸、苏氨酸。
2.4 灿烂弧菌刺激后菲律宾蛤仔消化腺代谢途径变化
菲律宾蛤仔感染灿烂弧菌后,代谢图谱变化见图4。由图4可知,升高的代谢物只有甜菜碱,降低的代谢物包括二甲基甘氨酸、甘氨酸、葡萄糖、胆碱、苏氨酸、谷氨酸、亚牛磺酸。
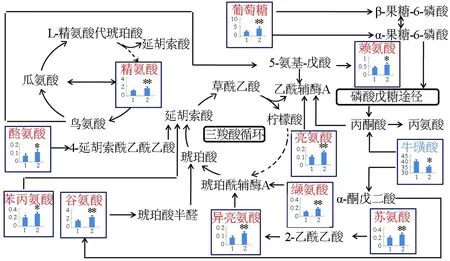
图3 鳗弧菌刺激后菲律宾蛤仔消化腺代谢图谱变化
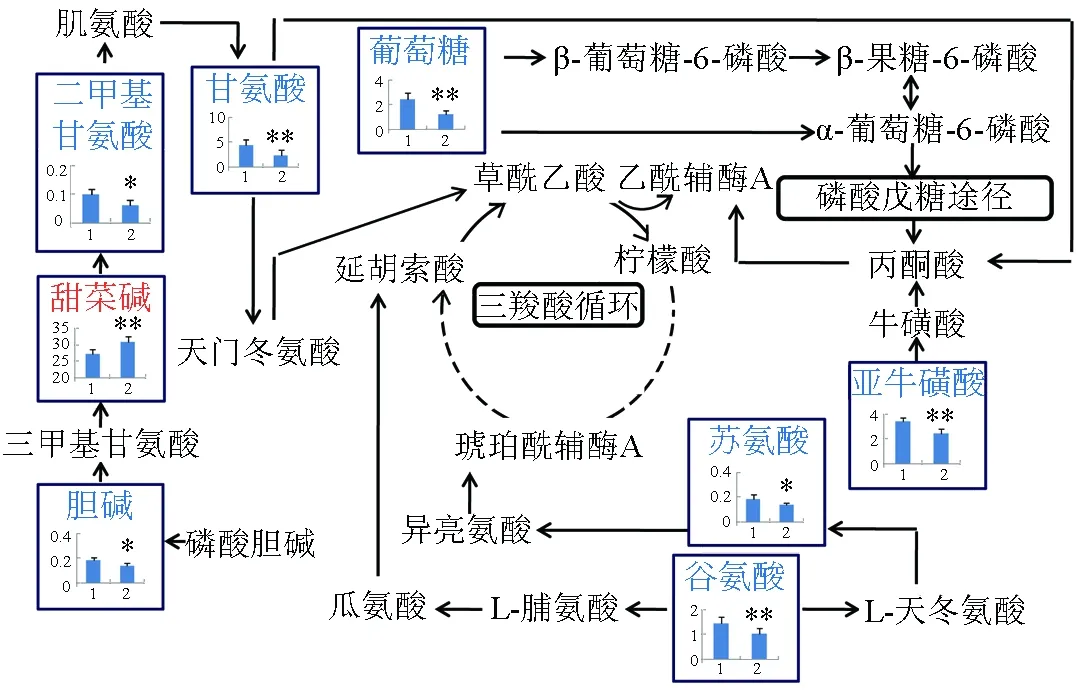
图4 灿烂弧菌刺激后菲律宾蛤仔消化腺代谢图谱变化
3 讨 论
从正常对照组、鳗弧菌感染组、灿烂弧菌感染组菲律宾蛤仔消化腺代谢物核磁共振一维氢谱图解析的代谢图谱可以看出,甜菜碱丰度最高(图1),甜菜碱广泛存在与动植物中,主要在渗透调节方面发挥着重要作用[23],这与菲律宾蛤仔生活在高盐环境中需要进行渗透压调节相符合。
在菲律宾蛤仔受到鳗弧菌和灿烂弧菌感染后,代谢物葡萄糖、苏氨酸、谷氨酸在两种菌刺激后均发生变化。通常贝类通过体内氨基酸含量的改变来调节细胞内渗透性,氨基酸及糖原直接参与体内的能量代谢过程[24],葡萄糖的改变证明弧菌可引起菲律宾蛤仔能量代谢发生变化。谷氨酸是中枢神经系统最重要的兴奋性氨基酸类神经递质,谷氨酸的改变推测弧菌可引起贝类神经系统毒性。苏氨酸直接参与体内蛋白质的合成,并且具有一定的免疫作用[25]。综上所述,两种弧菌刺激后共同发生的代谢变化包括能量代谢、谷氨酸代谢及蛋白质合成等。
鳗弧菌和灿烂弧菌均属于革兰氏阴性致病菌,寻找他们对水产养殖生物产生毒性的差异代谢物,对区分两种弧菌的毒性效应具有重要的意义,因此笔者构建了不同弧菌胁迫后菲律宾蛤仔代谢图谱。鳗弧菌暴露24 h后,代谢物牛磺酸降低,表明鳗弧菌引起菲律宾蛤仔低渗胁迫。亮氨酸、异亮氨酸、缬氨酸属于支链氨基酸,在免疫反应中主要涉及到氨基酸生成蛋白质的过程。精氨酸能提高鱼类抗氧化酶活性,抑制鱼类炎症反应,保护鱼体在免疫应答时免受自我损伤[26]。这些氨基酸的升高可补充牛磺酸的降低,从而对抗菲律宾蛤仔的低渗胁迫反应。苯丙氨酸对乙酰胆碱酯酶有一定的抑制活性[27]。赖氨酸是具有促进动物生长、发育以及神经系统机能完善等作用[28]。苯丙氨酸及赖氨酸升高与神经系统作用有关,鳗弧菌可导致贝类神经系统毒性反应,其具体机制有待于进一步研究。灿烂弧菌暴露24 h后,有关胆碱合成的代谢途径发生改变,主要包括二甲基甘氨酸、甜菜碱和胆碱。亚牛磺酸为牛磺酸的前体物质,其化学结构与牛磺酸相似,且还原性更强,本身具有强大的抗氧化作用[29],表明灿烂弧菌可引起菲律宾蛤仔氧化应激反应。
由此可见,鳗弧菌和灿烂弧菌引起菲律宾蛤仔代谢途径变化主要包括能量代谢、渗透调节、蛋白质合成及氧化应激等。表征鳗弧菌感染的代谢物包括牛磺酸、精氨酸、酪氨酸、苯丙氨酸、赖氨酸、亮氨酸、异亮氨酸、缬氨酸。而表征灿烂弧菌污染的代谢物包括甜菜碱、二甲基甘氨酸、胆碱、谷氨酸、亚牛磺酸。随着现代分子生物学技术的迅猛发展,尤其是“组学”技术(包括蛋白质组学、代谢组学、转录组学及基因组学)的逐渐兴起,在整体的生物学水平系统阐明贝类在受到弧菌感染下的响应特征指日可待。
[1] 吴后波,潘金培.弧菌属细菌及其所致海水养殖动物疾病[J].中国水产科学,2001,8(1):89-93.
[2] 杨少丽,王印庚,董树刚.海水养殖鱼类弧菌病的研究进展[J].海洋水产研究,2005,26(4):75-83.
[3] 张新中,张世秀,李海平,等.海水养殖动物致病性(Pathogenicity)鳗弧菌(Vibroanguillarum)的研究综述[J].现代渔业信息,2007,22(1):16-19.
[4] 李华,王扬,李强,等.灿烂弧菌的疏水性和生物被膜形成能力[J].中国水产科学,2011,18(5):1084-1091.
[5] 于永翔,王印庚,刘智超,等.基于gyrB基因的SYBR Green I 实时定量PCR检测病原灿烂弧菌[J].渔业科学进展,2014,35(3):134-142.
[6] 樊甄姣,杨爱国,吕振明,等.鳗弧菌注射对栉孔扇贝免疫活性的影响[J].南方水产,2007,3(6):52-55.
[7] Wang J,Wang L,Yang C,et al.The response of mRNA expression upon secondary challenge withVibrioanguillarumsuggests the involvement of C-lectins in the immune priming of scallopChlamysfarreri[J].Developmental and Comparative Immunology,2013,40(2):142-147.
[8] Wang X,Wang L,Zhang H,et al.Immune response and energy metabolism ofChlamysfarreriunderVibrioanguillarumchallenge and high temperature exposure[J]. Fish & Shellfish Immunology, 2012, 33(4):1016-1026.
[9] Li C, Li Y, Li T,et al. Two classes of glutathione S-transferase genes with different response profiles to bacterial challenge inVenerupisphilippinarum[J]. Fish & Shellfish Immunology,2012, 32(1):219-222.
[10] Duan Y, Liu P, Li J, et al. Molecular responses of calreticulin gene toVibrioanguillarumand WSSV challenge in the ridgetail white prawnExopalaemoncarinicauda[J].Fish & Shellfish Immunology,2014,36(1):164-171.
[11] 罗凯娅,刘欣欣,葛端阳,等.鳗弧菌(Vibrioanguillarum)侵染对青蛤(Cyclinasinensis)谷胱甘肽硫转移酶及其基因表达的影响[J].海洋与湖沼,2012,43(4):735-740.
[12] Zhang P,Li C,Li Y,et al.Proteomic identification of differentially expressed proteins in sea cucumberApostichopusjaponicuscoelomocytes afterVibriosplendidusinfection[J].Developmental and Comparative Immunology,2014,44(2):370-377.
[13] Yan F,Tian X,Dong S,et al.Growth performance,immune response,and disease resistance againstVibriosplendidusinfection in juvenile sea cucumberApostichopusjaponicusfed a supplementary diet of the potential probioticParacoccusmarcusiiDB11[J].Aquaculture,2014,420/421(2):105-111.
[14] Sun Y, Du X, Li S, et al. DietaryCordycepsmilitarisprotects againstVibriosplendidusinfection in sea cucumberApostichopusjaponicus[J].Fish & Shellfish Immunology,2015,45 (2):964-971.
[15] Li X, Jing K, Wang X, et al. Protective effects of chicken egg yolk antibody (IgY) against experimentalVibriosplendidusinfection in the sea cucumber (Apostichopusjaponicus)[J].Fish and Shellfish Immunology,2016(48):105-111.
[16] 刘青,张越,付鑫,等.菲律宾蛤仔的研究进展[J].河北渔业,2011(1):56-59.
[17] 郭永禄,任一平,杨汉斌.胶州湾菲律宾蛤仔生长特征研究[J].中国海洋大学学报:自然科学版,2005,35(5):779-784.
[18] 丁鉴锋,杨霏,闫喜武,等.不同壳色菲律宾蛤仔免疫机能的比较研究[J].大连海洋大学学报,2012,27(5):411-416.
[19] Liu X,Zhao J,Wu H,et al.Metabolomic analysis revealed the differential responses in two pedigrees of clamRuditapesphilippinarumtowardsVibrioharveyichallenge[J].Fish and Shellfish Immunology,2013,35(6):1969-1975.
[20] Nicholson J K,Lindon J C,Holmes E.“Metabonomics”:understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data[J].Xenobiotica,1999,29(11):1181-1189.
[21] Nicholson J K,Lindon J C.Systems biology:metabonomics[J].Nature,2008,455(7216):1054-1056.
[22] 葛端阳,赵婷,潘宝平.鳗弧菌(Vibrioanguillarum)对青蛤(Cyclinasinensis)的毒性及半致死浓度研究[J].海洋与湖沼,2012,43(6):1192-1195.
[23] Ferrino L A,Pierce S K.Betaine aldehyde dehydrogenase kinetics partially account for oyster population differences in glycine betaine synthesis[J].Journal of Experimental Zoology,2000,286(3):238-249.
[24] Calder P C. Branched chain amino acids and immunity[J].Journal of Nutrition,2006,136(Suppl 1):288-293.
[25] 王伟,叶金云,杨霞,等.中华绒螯蟹幼蟹对苏氨酸需求量的研究[J].动物营养学报,2015,27(2):476-484.
[26] Wang B,Feng L,Jiang W D,et al.Copper-induced tight junction mRNA expression changes, apoptosis and antioxidant responses via NF-kB, TOR and Nrf2 signaling molecules in the gills of fish:preventive role of arginine[J].Aquatic Toxicology,2015,158(19):125-137.
[27] 陈俊涛,曹婷婷,田云锋,等.新型L-苯丙氨酸类化合物的设计、合成及其对乙酰胆碱酯酶的抑制活性研究[J].中国药学杂志,2015,50(14):1180-1185.
[28] 邱红,黄文文,侯迎梅,等.黄颡鱼幼鱼的赖氨酸需要量[J].动物营养学报,2015,27(10):2057-2066.
[29] Nishimura T,Duereh M,Sugita Y,et al.Protective effect of hypotaurine against oxidative stress-induced cytotoxicity in rat placental trophoblasts[J].Placenta,2015,36(6):693-698.
MetabolicResponsesofManilaClamRuditapesphilippinarumtoVibrioanguillarumandVibriosplendidusChallenges
LIU Xiaoli, HOU Chengzong, LAI Yongkai, GAO Jun, LI Jie, SHANG Qiushuang, LONG Jing, DENG Xiaoyu, QIU Wenbin, SUN Hushan
( School of Life Sciences, Ludong University, Yantai 264025, China )
In this work, we characterized the metabolic responses induced byVibrioanguillarumandV.splendiduschallenges towards hepatopancreas of Manila clamRuditapesphilippinarumusing nuclear magnetic resonance (NMR)-based metabolomics. Metabolic responses showed that bothV.anguillarumandV.splendidusinduced disturbances in energy metabolism, osmotic regulation, protein biosynthesis and oxidative stress with different mechanisms. Three metabolites (glucose, glutamate and threonine) were altered in bothV.anguillarumandV.splendidusexposed groups. The metabolic biomarkers inV.anguillarum-treated groups included leucine, isoleucine, valine, arginine, lysine, taurine, tyrosine, and phenylalanine. And the metabolic biomarkers inV.splendidus-treated groups included hypotaurine, dimethylglycine, choline, betaine, and glycine. The findings indicate that metabolomics could be used to elucidate the biological effects of marine pathogens to the clam.
Vibrioanguillarum;Vibriosplendidus;Ruditapesphilippinarum; nuclear magnetic resonance (NMR); metabolites
10.16378/j.cnki.1003-1111.2017.02.004
2016-02-23;
2016-05-12.
国家自然科学基金资助项目(41506190);山东省高等学校科技计划项目(J14LE08);鲁东大学基金资助项目(LY2014018).
刘小莉(1979—),女,讲师,博士;研究方向:海洋生物学. E-mail:lxlshz2006@163.com.
S917.1
A
1003-1111(2017)02-0143-05
猜你喜欢
渔业现代化(2023年2期)2023-04-23 01:10:18
当代水产(2022年8期)2022-09-20 06:45:40
电脑报(2022年25期)2022-07-05 23:54:27
新农业(2022年6期)2022-04-13 13:37:47
食品安全导刊(2021年20期)2021-08-30 06:39:56
科普童话·学霸日记(2021年11期)2021-05-30 10:48:04
当代水产(2019年5期)2019-07-25 07:50:56
广东海洋大学学报(2015年3期)2015-12-22 10:05:26
中国当代医药(2015年23期)2015-03-01 02:05:50
河北渔业(2014年5期)2014-07-08 21:15:18
