
中国小型底栖生物研究的40年
——进展与展望*
2017-12-14 05:34张志南慕芳红刘晓收于子山
海洋与湖沼 2017年4期
张志南 周 红 华 尔 慕芳红 刘晓收 于子山
(中国海洋大学海洋生命学院 青岛 266003)
中国小型底栖生物研究的40年
——进展与展望*
张志南 周 红 华 尔 慕芳红 刘晓收 于子山
(中国海洋大学海洋生命学院 青岛 266003)
基于2016年希腊第16届国际小型底栖生物会议资料及我国已有的信息, 本文论述了国内外小型底栖生物的研究概况, 着重介绍了我国该领域的主要进展, 包括小型底栖生物的方法学、丰度与生物量的分布、海洋线虫和底栖桡足类的分类学、入侵植物种对线虫群落的影响、粒径谱和次级生产力、摄食生态学、实验室培养和生活史、环境检测、海洋线虫的分子生物学与系统演化、生态动力学-水层底栖耦合模型以及人才培养。依据国内外研究现状提出了今后的发展目标和应着重加强的研究方向。
小型底栖生物; 海洋线虫; 生物多样性; 水层-底栖耦合; 全球变化
Q958.8
10.11693/hyhz20170100022
海洋底栖生物(marine benthos)是海洋生物中栖息范围最广、生物多样性最高的生态类群, 其中的小型底栖生物(meiobenthos)是指分选时能通过 0.5 mm(或1.0 mm)孔径的网筛, 而被0.042 mm孔径的网筛(深海生物研究者建议用0.031 mm作为下限, 以阻挡更多细小的线虫)所阻留的一类底栖生物, 主要成分是多细胞后生动物(Metazoa), 也包括一部分原生动物, 如有孔虫、纤毛虫等。这类生物主要是指动物, 有时meiofauna与meiobenthos混用, 但后者范围更广一些, 如果研究对象是动物, 应使用meiofauna (McIntyre,1969; Heipet al, 1985; Higginset al, 1988; Giere,2009)。
小型底栖生物和渔业生物关系密切, 它是许多经济鱼类、虾类和贝类幼体阶段的优质饵料, 因而是底栖食物网中十分重要的环节。小型底栖生物的丰度和生物量波动直接控制着大型经济无脊椎动物幼体的定着和补充。小型底栖生物的生物量一般不超过大型底栖动物的20%左右, 但在近岸、河口、海湾和某些深海海底, 其生物量大体与大型底栖动物相当, 考虑到小型底栖生物极高的世代周转率, 若按年生产力计算则更十分可观。小型底栖生物是沉积物中异养细菌和碎屑的主要摄食者。不仅是水层-底栖生态系统耦合过程中关键的一环, 且对微生物摄食和调控具有全球尺度的效应(McIntyre, 1969; Heipet al, 1985;Montagnaet al, 1991; Coullet al, 1992)。国际最新的研究证明, 即使在大型动物很少存在的情况下, 小型底栖生物的筑管、摄食活动仍可改变沉积物的物理、化学和生物性质, 如颗粒输运、有机物质转移和微生物搬运, 这些活动极大地影响着底栖生态系统的服务过程, 如沉积物稳定性、生物地化循环、废物排放,也直接或间接地正面或负面地影响食物网动力学(Schratzbergeret al, 2016)。此外, 小型底栖生物野外采样简便, 物种丰富度极大(比大型动物高一个数量级), 生活周期短, 每年平均 3—5代, 生活史无浮游阶段等, 使这类生物成为海洋环境质量重要的指示生物。近几十年它们作为海洋生态监测和生态系统健康评估体系的一个指标, 已被广泛应用于海洋环境监测(McIntyre, 1969; Heipet al, 1985; Higginset al,1988; Coullet al, 1992; Giere, 2009; Schratzbergeret al, 2016)。
1 国际小型底栖生物的研究历史和现状
关于国际小型底栖生物研究的历史在本作者的评述中已有详尽的描述(张志南等, 2004a), 本文仅就几个重要的发展阶段作一评述。
1.1 19世纪中至20世纪50年代末, 是小型底栖生物研究的起步阶段(Higgins et al, 1988; Giere, 2009)
不同类群大量新种的鉴定和区系描述, 以及基于环境(主要是基于沉积物结构)定义不同的“群落”是其中心研究内容。抓斗式采泥器和底拖网的使用极大地扩展了小型底栖生物的取样和研究的空间(Nicholls, 1935), meiofauna一词被正式创立和确认,极大地推动了小型底栖生物的研究(Mare, 1942)。本阶段以德国学者 Remane(1933)和他的继承者们在波罗的海基尔湾和德国北海沿岸的工作最为经典和具有代表性。特别值得一提的是, 一大批欧美学者, 如Cobb、Filip’ev、Chitwood、Gerlach、Rieman和 Wieser等对小型底栖生物最优势的类群, 自由生活海洋线虫的分类学、系统学和生态学, 做出了重要贡献, 为后续小型底栖生物研究奠定了基础(Heipet al, 1985;张志南等, 2004a)。
1.2 20世纪60年代到80年代初, 是描述生态学和实验生态学起步阶段
小型底栖生物的类群组成、丰度和生物量分布及与栖息生境的关系, 小型底栖生物对颗粒沉积物粒径选择的实验, 开启了实验生态学研究(Warwick,2016)。氧-硫复合体对数量分布的影响, 捕食与被捕食者相互作用, 亚显微结构用于系统演化研究(Giere,2009)。1969年, 国际小型底栖生物学家协会在突尼斯的成立, 和第一届会议论文集与小型底栖生物研究手册的出版, 是该领域研究的一个重要的里程碑(Hulings, 1971; Hulingset al, 1971)。
1.3 20世纪80年代到90年代, 是功能生态学阶段
主流研究是借助实验室操作、野外现场实验和其他新技术, 了解小型底栖生物在生态系统中的作用。研究内容还包括生产力和能流的计算, 特定生境下的小型生物物种、群落分布适应, 如深海、两极和低氧缺氧带; 小型底栖动物的定着和扩散机制; 种群动态; 由人类活动和气候变化所导致的灾害事件对小型底栖动物的影响。小型底栖生物研究手册、小型底栖生物学、海洋底栖生物研究方法的出版以及互联网数据库NeMys和WoRMS小型底栖生物种名录的发布, 标志着小型底栖生物作为一门新兴学科已经成熟(Eleftheriouet al, 2005; Guiliniet al, 2016)。
1.4 20世纪90年代末至今, 全球变化与生物多样性保护阶段
变化中的海洋和人类活动对个体、种群、群落和生态系统的影响, 探讨生物在分子水平上的适应和响应机制, 生物分布格局和它的遗传背景, 极端条件下的遗传变化, 由污染等人类活动诱发的遗传改变、系统演化分析等。
“Psammonalia”是一个小型底栖生物学会刊物,始于 1966年, 当时每年发行两页, 仅作为美国同行通讯刊物。1974年扩展为国际刊物; 到20世纪末, 每年发行 4期, 年均 50页, 含有 400篇文献; 进入 21世纪以来呈逐渐下降的趋势。但这并不意味小型底栖生物研究的减少, 互联网搜索结果表明, 小型底栖生物文献呈线性增加, 这说明小型底栖生物学正逐渐融入普通生态学和生物海洋学的范畴。波罗的海和切莎皮克湾的网络模型、欧洲区域性模型, 以及更多的小型底栖生物文献, 出现在生态学和生物海洋学的主流期刊上, 就是一个有利的佐证(Warwick, 2016)。
2 我国小型底栖生物研究现状
2.1 启动和准备阶段
国家海洋局和中国海洋大学于1973—1975年主持了我国第一次海洋调查规范修改。海洋生物卷增加了小型底栖生物的调查内容, Smith-McIntyre弹簧采泥器、小型底栖生物取样管和微型底拖网的研制, 以及一整套海上和潮间带取样方法、室内筛选分离和资料分析方法的成功运用, 开启了我国小型底栖生物研究的准备阶段(国家海洋局, 1975; 张志南等, 2004a)。
2.2 描述生态学阶段——由“中-美”合作带动的大、小型底栖动物同步观测、取样和研究阶段
自20世纪70年代末和80年代初的改革开放以来, 我国许多学者出国进修并开展国际合作, 在国际第一届海洋线虫研讨会上(Plymouth, 1981年9月)展示了我国自由生活海洋线虫分类学和小型底栖生物生态学研究的成果(Plattet al, 1982; Zhang, 1983;Zhanget al, 1983)。80年代初, 中美长江口和黄河口沉积动力学联合调查(1980—1982年, 1985—1987年)先后开展, 属我国首批大型底栖动物和小型底栖动物的多航次同步观测、取样的综合研究, 引进了多种大型箱式采泥器, 并结合对沉积物的现场 X光剖面摄影, 研究大型和小型生物的扰动图形。1986年, 国家自然科学基金设立, “我国北方海域小型底栖生物的分类学、生态学和方法学研究”获得首批资助(3860804号)。与此同时, 20世纪80年代末, 中国海洋大学开始了第一批硕士生的培养并相应地开设了多门有关小型底栖生物学的分类学、生态学课程, 包括实验课和海上调查实习课。
2.3 多学科交叉的海洋生态动力学阶段
20世纪90年代国家教委和国家基金委先后在胶州湾和渤海启动了海洋生态动力学胶州湾预研究(1995—1997年)和渤海国家自然科学基金重大项目:渤海生态动力学(1997—2001年), 自此我国的小型底栖生物研究开启了多学科交叉的时代。小型底栖生物作为底栖生态系统的一个重要环节, 正式进入水层-底栖耦合模式, 充当了联结水层和底栖过程的一个关键变量。与此相配合的两次“中-英”国际合作(1995—1997年, 1997—2000年), 开展了海洋生态动力学和渤海小型底栖生物多样性(海洋线虫和底栖猛水蚤),后者同时获得了英国环境部“Darwin Initiative”项目的资助, 这是我国最早获得的国际生物多样性项目之一。我国首批小型底栖生物博士生的中-英联合培养, 一系列博士生课程的开设以及大型统计软件PRIMER的引进和应用, 开启了高层次人才培养及我国大小型底栖动物同步观察研究的生态动力学研究新阶段(张志南等, 2017)。
2.4 开启全国性调查——向实验生态和功能生态迈进的阶段
进入21世纪, 小型底栖生物作为水层-底栖耦合过程中的一个关键度量, 被正式纳入国家重点基础研究发展计划(973)项目, 开展了区域性尺度(黄、东海)的调查。我国首部生物扰动实验系统(AFS)和底栖群落生产力的测量系统(BCSPM)的建立, 以及底栖动物粒径谱和次级生产力的测定, 标志着我国的小型生物研究由描述性生态学逐渐向实验生态和功能生态学过渡(张志南等, 1999, 2000a, 2005, 2017)。
3 主要进展
3.1 小型底栖生物的方法学研究
3.1.1 三次海洋调查规范修改编写(1973—1975年,1986—1988年, 2005—2007年)
借助国际合作和国内重大项目的执行, 试制了国外先进的取样仪器, 如 Smith-Mclntyre弹簧采泥器、改进型Gray-O’Hara箱式采泥器, 多种取样管和雪橇式小型底拖网, 并引进了数据处理大型软件包PRIMER 5.0及6.0, 总结编写了从潮间带至陆架浅海的取样、室内分选、数据处理和集成的一整套符合我国国情的小型底栖生物研究方法和操作流程, 特别是最近的海洋调查规范(GB-国家标准 2007), 全部内容被列入国家“908”专项项目的技术操作规程, 开展了全国性的综合性调查, 这是我国小型底栖生物研究发展的一个里程碑(张志南等, 2007)。
3.1.2 小型底栖生物群落集群分布的测定和微尺度(cm级)的发现, 为陆架浅海取样设计提供了依据1987年 10月“中-美”黄河口联合调查, 使用美国Gray-O’Hara大型箱式采泥器, 结合内径2.2cm, 管内距为3cm的4×4复式取样管, 对16个芯样的小型底栖动物的丰度和线虫进行了种类鉴定和空间分布型的测定、扩散卡方检验。结果表明, 海洋线虫主要种群的分布为集群分布, 拟合负二项分布模型、异质性卡方值指明, 在所研究的样品分离尺度上可作出微生境(cm级)的划分。线虫群落的种类组成和主要种类取食类型的分析表明, 食物的可获性和幼体大量补充的机制是特定条件下影响线虫微尺度空间分布的主要机制(张志南等, 1989; 张志南等, 1990a)。
3.1.3 不同内径取样管取样效率及重复芯样计数变异性的测定 1984年在秦皇岛砂质滩分别对1.8、2.2和 3.4cm内径取样管取样计数, 用单因子方差分析和 LSR多重比较法检验结果, 这三种内径取样管所得海洋线虫计数的平均数之间没有显著差异(α=0.05)。
依据对秦皇岛砂质滩和黄河口水下三角洲E2站位 16个连续芯样的计数测试, 给出了海洋线虫、底栖桡足类和介形类一个芯样计数和2、3个芯样计数平均数的 95%置信区间, 论证了潮间带和浅海软底小型底栖生物 2.2cm内径取样管及不同取样次数的可行性和可信度, 为海洋调查规范编写提供了统计学依据(张志南等, 1989, 1990a、b, 2007)。
3.1.4 小型底栖动物样品分选方法探索 从沉积物中分离小型底栖生物样品耗时费力, 基于小型底栖生物的荧光特性, 设计了一种激光诱导荧光检测技术, 应用于小型底栖动物的分离。该技术已应用在深海样品的分离, 试验表明, 分离准确率高达95%以上, 展现了良好的应用前景(叶树明等, 2005, 2006)。
3.2 主要类群的分类学研究
3.2.1 自由生活海洋线虫(free-living marine nematodes, 以下简称海洋线虫)分类学研究 海洋线虫通常是小型底栖生物中最优势的类群, 在陆架浅海的丰度占小型生物总丰度的60%—90%, 在河口和有机质较丰富的生境可达95%以上。按生物量排序,海洋线虫、小型多毛类(包括幼龄个体)和底栖猛水蚤通常占据小型底栖动物的前三位(张志南等, 1989,1990a; 张志南, 1991)。1983年, 青岛湾有机质污染带三个新种的描述, 开启了我国海洋线虫分类学的研究(Zhanget al, 1983)。自20世纪90年代开始, 特别是进入 21世纪, 伴随着国际合作和一系列国际自然科学基金项目的启动, 海洋线虫分类学有了较快的发展。至今, 在渤海、胶州湾潮下带、岩礁海藻、砂质滩等已鉴定海洋线虫174种, 包括24个新种(Zhang,1990, 1991, 1992, 2005; Zhanget al, 1994, 2005, 2006b,2012; Hopeet al, 1995; Huaet al, 2007; Huanget al,2005, 2006a、b、c、d, 2007, 2009, 2010, 2012, 2013)。初步建立了我国黄海和东海海洋线虫的种名录, 鉴定黄海线虫170种, 其中新属3个, 40余个新种。鉴定东海线虫近300种, 发现新属5个, 新种20余个(蔡立哲等, 2000a, b; Guoet al, 2001; Huanget al, 2006a,b, c, d, 2007, 2009, 2010, 2012, 2013, 2015, 2016; Fuet al, 2013; Chenet al, 2014; 2015; Jianget al, 2015; 蔡立哲, 2015; Guoet al, 2015; 2016; 史本泽, 2016; Liet al, 2016a, b; Sunet al, 2016)。在东海红树林湿地和沙滩潮间带共发现新种 20个和一批新记录, 其中包括在极地楚奇科海发现的 1个新种(Guoet al, 2001,2015, 2016; Chenet al, 2014; 2015; Liet al, 2016a, b)。南海海洋线虫分类学研究也已起步, 已记录和描述台湾海峡和北部湾自由生活海洋线虫300多种, 已有少数新属新种的记录(Fuet al, 2013; 蔡立哲, 2015)。基于传统分类学(光镜和扫描电镜)和18S rRNA基因序列分析发现并描述的一新种——太平湾嘴刺线虫(Enoplus taipingensisZhang & Zhou, 2012)标志着我国海洋线虫集成分类学(integrative taxonomy)的开端(Zhanget al, 2012)。
据初步估计, 我国海洋线虫常见种约有 1000种(张志南等, 2003), 已有记录约500种, 世界已有记录约7000种(Appeltanset al, 2012)。总体说来, 我国海洋线虫分类学研究还处在正在加速的初级阶段, 系统演化和动物地理学分析亟待加强(Andrássy, 1976;张志南等, 2003, 2004a)。
3.2.2 底栖猛水蚤桡足类 我国猛水蚤的研究始自20世纪50年代至70年代。在20世纪90年代, 受国家自然科学基金和中-英Darwin Initiative项目推动和支持, 猛水蚤的分类学研究重新起步并获得一定进展。至今, 已记录 213种; 其中, 在渤海鉴定 116种(或分类实体), 包括新属2个, 新种8个, 对一些优势种和常见种类进行了重新描述, 对部分属进行了系统发育分析和修订(Geeet al, 2000; Muet al, 2000a,b, 2004; Huyset al, 2008)。黄海及邻近海域鉴定了24种包括 1个新属, 17个新种(马林, 2011; Maet al,2011), 台湾海峡鉴定了 63种和分类实体(王彦国,2008)。
至今国际上已发现海洋猛水蚤 56科, 589属,4300种(Wells, 2007), 仍有很大一部分有待发现(Huyset al, 1996), 我国猛水蚤的分类学研究亟待加强。
3.2.3 其他门类 动吻动物门 Kinorhyncha, 已记录1目1科6属10种, 仅1种鉴定到种, 烟台棘皮虫(黄宗国, 2008; 黄宗国等, 2012)。
曳鳃动物门Priapulida, 已记录曳鳃虫科2种(黄宗国, 2008; 黄宗国等, 2012)
环节动物门寡毛亚纲 Oligochaeta, 在青岛胶州湾水域和香港水域记录4科, 27属69种(黄宗国, 2008;黄宗国等, 2012)
节肢动物门海蜘蛛纲 Pycnogonida, 皆足目Pantopoda已记录4属4种(黄宗国, 2008; 黄宗国等,2012)
节肢动物门蛛形纲Arachnoida蜱螨亚目Acari已记录9科23属49种(黄宗国, 2008; 黄宗国等, 2012)
3.3 丰度和生物量分布
3.3.1 浅海陆架 我国陆架浅海(100m 以浅)小型底栖生物的年均丰度为 377—1971ind/(10cm2), 生物量值为 468—1393 μg dwt/(10cm2), 这与国际上陆架浅海的量值十分接近和吻合[丰度为1000ind/(10cm2),生物量为1000—2000μg dwt/(10cm2)] (Higginset al,1988; 张志南等, 2000b, 2001a、b, 2002, 2004b; 孙松,2012), 北黄海冷水团夏初的小型底栖动物丰度和生物量分别达到(3410±1578)ind/(10cm2)和(2766±1344)μg dwt/(10cm2)是我国浅海水域的最高值(王家栋等, 2011)。东海长江口及陆架浅海丰度和生物量值次高, 海洋线虫的丰度值占 91%, 生物量值占 51%,由近岸向离岸优势度下降但多样性增高(华尔等,2005)。南海小型底栖生物丰度最低, 约为东海的1/5,生物量约为东海的 1/3。两个量值由沿岸向离岸逐渐下降。我国四大海域小型底栖生物的丰度和生物量见表 1(孙松, 2012)。
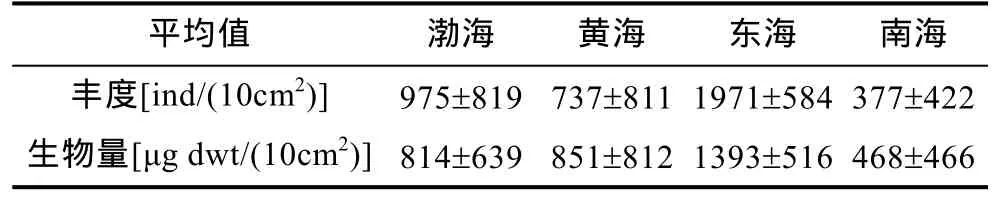
表1 我国主要海区小型底栖生物的丰度和生物量
南黄海、渤海的丰度和生物量居中。但水平分布呈现不同的格局, 黄海呈现明显的镶嵌式斑块分布(Liuet al, 2005, 2007), 这与黑潮暖水的入侵、长江口冲淡水逆流北上支流的涌入以及冷水团的存在, 所导致的沉积生境的多样性密切相关。南黄海冬季航次共鉴定线虫 233种(分类实体), 沉积食性者(1A+1B)占优势, 幼龄个体占个体总数的60%以上, 雌雄比例平均为 1∶0.79。南黄海线虫群落可划分为受沿岸水影响的沿岸群落、冷水团群落和两者之间的过渡型群落(Liuet al, 2007)。渤海小型生物的丰度和生物量值呈现中北部高而西南部低。四个海域的丰度和生物量值均表现为50%—70%的量值分布在0—2cm表层(黄勇, 2005; Liuet al, 2005, 2007; 张艳, 2006)。
3.3.2 陆坡和深海 南海北部陆坡两个站位的小型底栖生物丰度分别为(236±72)ind/(10cm2)和(725±236)ind/(10cm2), 线虫优势度达95%。太平洋深海小型底栖生物的丰度为 26—313ind/(10cm2), 线虫优势度达91%—97%, 这与国际上的报道相接近(Higginset al,1988; Giere, 2009)。目前陆坡深海取样的仪器包括0.1m2大洋型采泥器, 0.25m2箱式采泥器和 MCS-1多管取样器(见表2)。深海取样分选尚需规范化(王小谷等, 2005;杨俊毅等, 2005; 刘晓收等, 2014; 史本泽, 2016)。
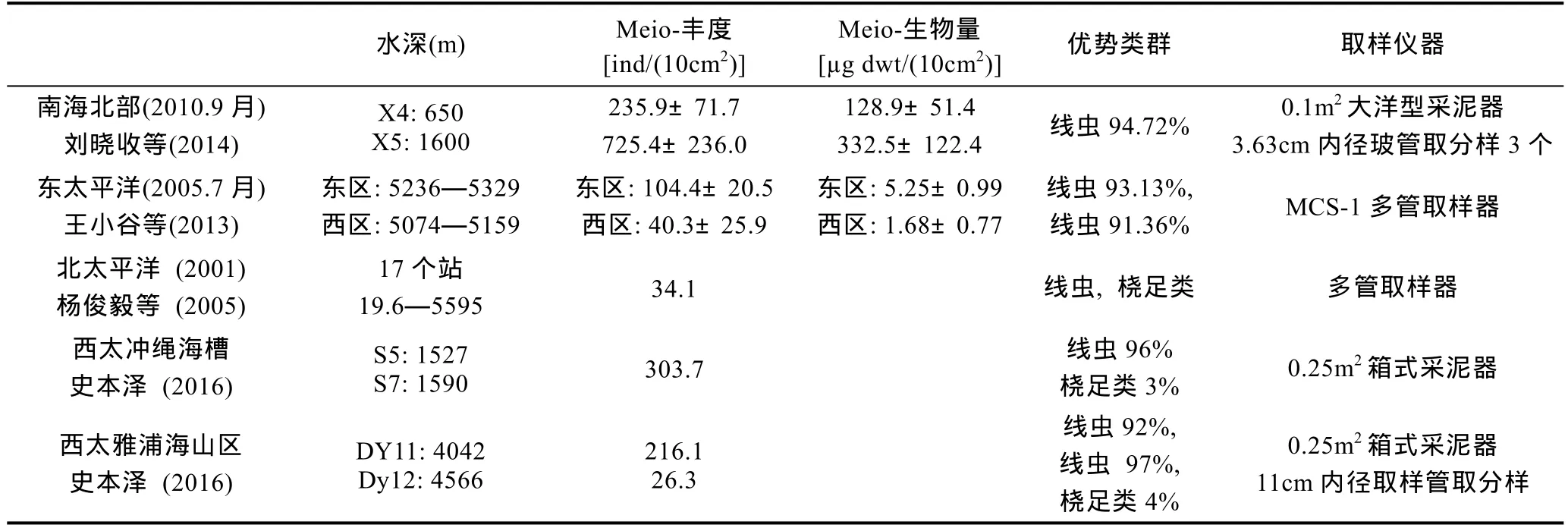
表2 深海小型底栖生物丰度和生物量
3.3.3 局域尺度分析——物理扰动与生物扰动的交替出现 在黄河口和长江口水下三角洲及邻近海域发现了不同扰动梯度。如黄河口等深线 10m内属于高沉积速率区, 缺乏大型底栖动物, 但存在小型底栖动物, 特别是海洋线虫; 等深线10—20m之间的莱州湾为生物扰动区, 栖息着大型底栖动物, 如心形海胆、筑穴蛤类、筑管多毛类以及典型的以线虫和猛水蚤为代表的小型底栖动物群落; 等深线 20m以外则属于典型的浅海陆架水域, 生物群落由水动力(渤海环流)所控制。由黄河口至渤海中部线虫多样性依次递增分别为H′=3.60±0.24,H′=3.95±0.30 和H′=4.94±0.23。线虫生物多样性与海水盐度和沉积物异质性指数(h)呈显著负相关(P≤0.01)(张志南等, 2000b,2001a, b)。
胖子不耐烦地给了卡车司机五百元钱,把他打发走了,偷运树木与卡车司机是没有什么关系的,卡车司机只嘀咕说格老子的亏大了,油钱都不够。胖子说,算了算了,快走快走,我亏得还大呢!
3.3.4 潮间带线虫大尺度空间分布的研究 沿中国沿海(20.54°—44.88°N)在湿地和红树林设 16个站点, 共获得53个样品。结果表明, 沿中国沿海湿地线虫的分类学多样性随纬度的增加而下降, 而系统演化多样性指数(AvTD和 AvPD)随纬度增加而显著增大。这表明, 在较低的纬度, 线虫群落的物种十分相近, 随着纬度的增加, 选择沉积食性者的物种丰度和个体数量相对减少, 而非选择食性者增加。营养结构分析显示, 低纬度线虫的小生境宽度因摄食的特化相对较窄, 高纬度气候变异大, 更有利于机会种的生存, 由此, 低纬度线虫群落含有更加密切相关的种(Wuet al, 2016)。
3.3.5 岩礁附植小型底栖动物 对大连石槽14种海藻(向浪和背浪面)上共鉴定出小型底栖动物 11个类群, 大型底栖动物4个类群。海洋线虫和底栖桡足类的相对丰度分别是41%和28%, 按每克干重海藻求得的平均丰度值是 1021ind/(g·d·w)[3.1—10107ind/(g·d·w)]。共鉴定附植线虫 46种, 其中, 平均相对丰度≥1%的优势种和见习种有16种。不同藻类, 不同位置(向浪和背浪)种类组成差异显著。如多管藻上共发现线虫26种, 在背浪面单宫类线虫Theristus aier占70%, 而在向浪面退居第二位, 第一优势种为色矛类线虫Monopothia costata, 这种差异归因于叶面上沉积的颗粒碎屑种类和数量(张志南等, 1994)。
此外, 附植动物区系的工作还包括对中国香港自然保护区马尾藻, 青岛6种海藻连续周年逐月的调查及附植小型底栖动物 ATP的测定。该研究测得单个线虫 ATP含量平均为 1.18—7.01ng/ind, 底栖桡足类为 1.42—19.64 ng/ind, 线虫个体干重与 ATP的比值为43∶1—99∶1, 该值与Sikora等的结果相近, 但略偏低, 该研究为进一步开展 ATP用作指示底栖生物生物量的生理生化指标提供了依据(Zhanget al,1997; 林岿璇等, 2003; 张艳等, 2005; 陈海燕等,2011)。
3.3.6 红树林湿地小型底栖动物 蔡立哲在深圳河口福田秋茄林潮间带一年逐季共记录海洋线虫 28种, 其中, 优势种均属于单宫目种类, 分别是隆线虫科的Daptonemasp., 条线虫科的Metalinhomeussp.和Terschellingiasp.以及轴线虫科的Paradontophorasp.。郭玉清对厦门凤林红树林湿地小型底栖动物的研究指出, 海洋线虫是最优势的类群, 丰度占 76.1%—96.3%, 优势种分别是色矛目的Sabatieriasp.,Spilophorellasp.和Chromadorinasp.。对香港秋茄林湿地的研究指明其线虫优势种都以沉积物食性种类占优势, 显示以上3个生境中, 同一植被的线虫群落某种程度的相似性(蔡立哲等, 2000b; 郭玉清, 2008)。
3.3.7 渤海小型底栖动物数量的十年际变化 依据渤海重点基金2008年8月和2009年6月2个航次与20世纪90年代重大基金3个航次, 以及20世纪80年代“中-美”合作3个航次, 共8个航次的对比研究发现, 过去 30余年, 渤海小型底栖动物最优势类群海洋线虫的丰度和生物量逐渐增加, 而底栖桡足类丰度却在逐渐减少, 因此线虫桡足类数量比值(N/C)逐渐增加, 这与沉降粒度变细、有机质含量增加,导致大型底栖动物群落改变有关。但是否可看作是对渤海人类干扰胁迫影响的一种响应, 尚待进一步探讨(张志南等, 2017)。
3.4 植物种入侵对线虫群落的影响
长江口入侵互花米草群落与两种本地植物群落的对比实验表明, 入侵植物种显著地降低了线虫营养结构的多样性, 减化了底栖食物网结构。入侵植物群落中, 食细菌线虫丰度的增加反映了分解过程、分解速率和营养通路的改变, 依次改变了河口生态系统地下水的营养盐循环。沉积物水含量、电导率、细菌和凋落物的生产量是形成线虫群落的最重要的环境因子(Chenet al, 2007)。
3.5 底栖生物的粒径谱和次级生产力
3.5.1 概述 粒径谱(Size Spectra或Biomass Size Spectra, BSS)在国际上已广泛被用于各种水体的群落结构和生态过程机制解释。若用单位粒级的正态化生物量密度[B(w)=B(w)/Δw], 取代Sheldon型粒径谱的纵坐标, 则得到正态化粒级谱(Normalized Biomass Size Spectra, NBSS), 由于NBSS图形能够更好的应用于定量分析和积分运算, 因此NBSS得到广泛应用(张志南等, 2017)。在21世纪初, 通过国家“973”项目的实施, 先后在胶州湾和东黄海建立了底栖动物粒径谱, 验证了底栖粒径谱的多峰状分布, 并利用粒径谱理论对南黄海底栖动物群落生产力和耗氧量进行了计算, 获得可比较的结果(林岿璇等, 2004; 邓可等, 2005; 华尔等, 2009; Huaet al, 2013)。其后, 通过国家自然科学基金重点项目, 在渤海已建立 Sheldon型粒径谱图形基础上, 对三峰模式进行了深入的探讨, 各站位波峰波谷的位置基本一致, 三峰结构在粒级上分别对应自由生活海洋线虫、其他小型底栖动物类群和大型底栖动物。这表明, 线虫和其他小型动物类群在食物网中的功能是有差异的(Huaet al, 2013)。
3.5.2 粒径谱与底栖生物次级生产力 邓可等借鉴 Blanio等浮游生物群落次级生产力计算公式, 将粒径谱方法引入南黄海和渤海底栖动物次级生产力计算(张志南等, 2017)。用粒径谱方法计算得到渤海底栖动物次级生产力年平均为8.52g dwt/(m2·a) [4.87—15.68 g dwt/(m2·a)], 研究站位大型底栖动物的次级生产力占总次级生产力的 60%—89%, 相应的, 小型底栖动物次级生产力仅占总次级生产力的 11%—40%。NBSS方法计算的小型底栖动物次级生产力与固定系数法(P/B=9)结果相近但略低(ANOVA,F=4.863,P=0.05), 原因是P/B值随环境和类群的不同存在显著的差异(邓可等, 2005)。由此, NBSS方法对不同粒级生物分别计算, 考虑了生物大小对P/B值的影响, 其结果更可靠。与以营养级或分类阶元为基础的次级生产力计算方法相比, 粒径谱方法较简单,但仍需更广的海域和不同的季节加以验证。
3.6 摄食生态学
3.6.2 渤海小型底栖动物的稳定同位素示踪摄食实验 研究站位底栖微藻天然δ13C值为–21.02±0.31。线虫天然 δ13C 值为–20.23±1.32, 其他类群的天然δ13C值为–19.75±1.17。研究站位底栖微藻初级生产力水平远低于潮间带, 仅为 116.41μgC/(10cm2·h), 研究站位水深和悬浮物对光照的遮蔽可能是导致底栖微藻群落本身生产力低的原因。
小型底栖动物的摄食率为3.33μgC/(10cm2·h), 线虫对底栖微藻未表现出明显的摄食富集。其他类群的总摄食量高于线虫。
小型底栖动物对底栖微藻的摄食仅占初级生产力的2.86%, 渤海底栖生态系统可能以碎屑食物链为主, 啃食食物链相对作用较小(张志南等, 2017)。
3.7 实验室培养和生活史的研究
3.7.1 海洋线虫 王诗红等(1994)开启了我国养虾池-优势线虫Diploaimellasp.的实验室培养, 该种在玉米粉琼脂培养基中大量培养成功。在温度为(25±1)℃和盐度为 20条件下, 观察了胚胎及胚后发育过程并绘制了生长曲线, 分析了蜕皮周期及幼龄个体的年龄划分。观察到最短世代时间Tmin=16d, 计算得出实验室种群的内禀增长率rm=0.185/d。在培养条件下的生殖方式为孤雌生殖(王诗红等, 1994)。
王摆等(2007)以玉米琼脂培养基在(23±0.5)℃盐度为 26.7条件下成功地培养了来自栈桥潮间带底泥中的一种线虫Chromadorinasp.观察到雌雄线虫的交配, 据推测该种为两性生殖, 最短世代时间为14d(王摆等, 2007)。
国外海洋线虫的培养始自20世纪60年代, 至20世纪80年代已利用多种培养线虫开展河口沉积物的生物测试(Tietjenet al, 1984), 实验种群的培养是开展实验生态, 种群动态, 生态毒理和分子生物学研究的前提条件, 我国此类研究亟待加强。
3.7.2 底栖猛水蚤 陈世杰1988年开展了厦门港尖真额猛水蚤(Euterpe actifrons)的室内培养, 平均密度达 3689ind/L, 目前增殖率 0.149。陈珂(2004)设计和应用 100L封闭循环式培养系统培养美丽猛水蚤(Nitocresp.)产量达 68—182ind/(L·d), 最大收获量为1.28×105ind。
目前底栖猛水蚤中大规模培养比较成功的有日角猛水蚤属(Tisbe), 虎斑猛水蚤属(Tigriopus)和美丽猛水蚤属(Nitocre)。除了作为饵料生物培养以外, 我国学者还开展了杀虫剂、环境内分泌干扰物及重金属等污染物对猛水蚤的毒理学研究。近期, 联系全球变暖, 利用实验室种群开展了多项海洋酸化的生物学影响效应研究(陈世杰, 1988; 许永香等, 1992; 修瑞琴等, 1995; 陈珂, 2004; 朱丽岩, 2005; 张达娟等,2011; 曾艳艺等, 2011; 孙艳桃等, 2014; 韦晓慧等,2014; Caoet al, 2015)。
3.8 环境监测
3.8.1 自然种群和群落指示种对环境扰动的响应青岛湾有机质污染潮间带研究发现, 海洋线虫的最大密度可达 10298.6ind/(10cm2), 而且海洋线虫密度与小头虫(Capitella capitata)有着相似的分布特点。依据线虫群落多样性指数, 线虫取食类型及桡足类的相对丰度, 得出该湾有机质污染程度正在缓解的结论(张志南等, 1993)。种类组成分析发现, 栈桥后瘤线虫(Metoncholaimus molesZhanget al, 1983)是最优势的种群, 其密度沿污染梯度有一显著的季节稳定的空间分布图型, 可作为该有机质污染带的生物指示种(张志南等, 1993; 党宏月等, 1996)。
3.8.2 线虫与底栖猛水蚤桡足类密度比值(N/C)N/C比值, 方便, 快速, 免去繁琐的分类鉴定, 在国内有机质污染研究中得到广泛的应用, 但由于受到季节变化及沉积物粒度的影响而限制了它的利用,在潮下带由于受沉积环境的异质性以及线虫和桡足分布的不均匀性, 是否能应用于污染监测有待进一步研究。在渤海湾水域的研究中发现,N/C比值反映了渤海的污染状况, 但N/C值受沉积物取样的深度影响大。而 Sun等 (2014)通过对青岛和舟山具有相似沉积物粒度组成但受不同程度人类活动干扰沙滩的比较研究, 证实了N/C比值在沙滩应用的有效性并发现该比值与沙滩沉积物中的叶绿素a含量呈正相关。总之,N/C值的单独使用尚在探索中, 判断一个海域的污染状况应利用多元统计分析, 综合多种生物和非生物因子进行判断(郭玉清等, 2002; 张青田等,2008, 2012)。
3.8.3 线虫群落结构对扰动的响应 20世纪90年代对渤海中、南部, 胶州湾和湄州湾共 11个站位的线虫群落结构进行比较分析。共鉴定线虫154种, 分属于 107个属29个科, 在种类组成分析基础上分别在种、属和科的水平上进行聚类和排序, 结果表明:①论证了黄河口水下三角洲, 莱州湾和渤海中部这一特有环境梯度的存在, 即高沉积速率的扰动-生物扰动-海流的物理控制。②莱州湾和渤海湾中部的线虫群落与20世纪80年代相比, 种类组成有了显著的改变, 非选择沉积食性(1B)显著增加, 而附生生物食性(2A)显著减少, 表明有机碎屑量的显著增加, 这与调查海域沉积物中粉砂含量增加有关。③在物种和属水平上的多元统计图形, 显示线虫群落结构无显著差别, 即, 对线虫而言, 鉴定到属的样品分析可满足监测的需要, 省略了繁琐的分类到种的鉴定。④对渤海线虫群落的Caswell中性模型分析显示, 大部分站位的“V”值位于“–2—2”范围内, 说明未受到外界因子的显著扰动。渤海中部站位的“V”值为–2.054指示环境的稳定性增强。而莱州湾站位的“V”值为–2.624, 显示已受到外界因子显著的扰动, 这与黄河口输送大量的粉砂质沉积物有关, 中性模型分析结果与上述种类组成分析结果基本上是吻合的(张志南等, 2000b, 2001a、b)。
3.8.4 对浒苔大量暴发的监测 (1) 砂质潮间带。2012年浒苔暴发早期小型底栖动物的平均密度为(1036±135)ind/(10cm2), 大量暴发期平均密度为(815±196)ind/(10cm2), 消退后为(865±201)ind/(10cm2)。浒苔大量暴发增加了海洋线虫的优势度, 降低了桡足类的比例, 显示了对小型底栖生物数量和类群结构的负面影响, 不同潮间带影响有所不同。浒苔暴发可影响叶绿素浓度和溶氧含量, 最终影响小型底栖动物的数量和结构分布(华尔等, 2015)。
(2) 南黄海 2008年夏季浒苔暴发末期的监测。黄海小型底栖动物总体现有量较2007年低约1/3, 并在浒苔暴发区域下降更为明显。但南黄海冷水团海域却略有上升, 原因可能是浒苔暴发后碎屑的沉降和降解过程所致(吴秀芹等, 2010)。
3.8.5 海洋线虫群落对台风的即时响应 2003年6月末, 在长江口一站位监测到“Soudelar”台风过后,线虫丰度增加 66.1%, 而在 5—8cm沉积物层线虫丰度增加了96.9%, 沉积物的叶绿素a(Chla)和脱镁叶绿酸a(Pheoa)分别增加了147.3%和56.4%, 底水氧增加了 8倍, 这指明, 台风引起水层混合, 再悬浮和生物沉降过程是导致线虫数量增加的主要环境因子(Huaet al, 2010)。
3.8.6 实验生态与野外试验 (1)实验室微型受控生态系的研究包括砂滩海洋线虫对缺氧的响应(华尔等, 2012), 有机质富集对砂质滩小型底栖动物的影响(Wanget al, 2011)。
(2) 野外试验, 香港维多利亚港污水停排后小型底栖动物和线虫群落响应试验; 大连湾局部海域石油污染对潮间带海洋线虫群落的影响; 象山港电厂热水排放对小型底栖动物群落的影响。分类多样性指数用于环境扰动监测(Guoet al, 2002; Liuet al, 2011;Lvet al, 2011; 谢礼等, 2012)。
3.9 国内小型底栖生物分子生物学研究进展
在国内开展较早的小型底栖生物分子生物学研究始于 2005年, 沈锡权等(2005)开展了单条线虫基因组DNA提取和18S rDNA基因PCR扩增的方法学研究, 并采用以18S rDNA为分子标记的限制性片段长度多态性(RFLP)方法进行了青岛潮间带沉积物海洋线虫的分子生物多样性研究(沈锡权, 2005; Shenet al, 2007); 邹丽珍(2006)则以18S rDNA为分子标记,研究了东北太平洋多金属结核区深海四个沉积物环境DNA样品的分子多样性。这类分子生物学研究方法, 由于缺乏形态分类学鉴定, 对所获得的 DNA序列只能通过与现有数据库中较少的序列进行比对,来获得小型底栖生物的分类学信息。中国海洋大学底栖生物实验室以海藻附植线虫为研究对象, 通过单条线虫DNA提取, 将形态分类与DNA分子鉴定相结合, 成功推演了基于18S rDNA序列的15种海藻线虫的分子系统发育关系, 并在新种的形态学描述中首次结合了 DNA分子生物学信息(Zhanget al, 2012);在国家自然科学基金(40730847号, 41376146号)的支持下, 以国际条形码数据库BOLD为平台, 初步建立起中国海洋线虫 DNA条形码参考数据库(张志南等,2017)。具体进展如下:
3.9.1 岩礁附藻线虫的分子生物学多样性及系统演化 以18S rDNA部分序列为分子标记, 对青岛太平角岩礁 15种附藻线虫进行了分子多样性研究, 并根据最大似然模型, 构建了 15种线虫的系统演化关系。该研究结合2007年11月至2008年10月之间开展的连续 12个月的附藻线虫生态学调查, 共鉴定出35个形态学种(或分类实体单元)。优势种为Enopluscf.communis和Eurystomina ophthalmopra, 前者依据形态和分子生物学特征被定为新种Enoplus taipingensis(Zhanget al, 2012; 张志南等, 2017)。
实验结果以18S rDNA作为分子标记较成功地推演了15种海藻线虫的系统演化关系。相对于色矛目,嘴刺目(Enoplida)分支得到较高的自展置信值支持。实验证明, 以18S rDNA系列为标记进行的海藻线虫分子鉴定是十分可行的, 因为每个线虫种都得到了100的自展置信值。
3.9.2 中国海洋线虫DNA条形码参考数据库 截止 2014年 7月, 已完成黄渤海潮间带和潮下带海洋线虫共192个个体, 52种线虫的条形编码, 其中, 获得COI-5P标准条码36个(占19%), 完成编码的种类为 15种(占 29%), 共 19个 DNA 条形码索引号(Barcode Index Number, BIN)。另外, 该数据库还包括127个个体(占 66%)40种海洋线虫(占 77%)的 18S rDNA有效序列。以上结果证明, 海洋线虫18S rDNA部分序列的编码成功率要远高于 COI-5P, 这表明, 相对于更加保守的18S rDNA细胞核糖体基因, COI线粒体基因应用通用引物进行扩增和测序的难度更大; 这也说明, 为什么国际上对于线虫(淡水、海洋和寄生)的分子鉴定大部分采用18S rDNA基因(张志南等, 2017)。
从分类学构成分析, 该数据库包含的 52种海洋线虫隶属于 5个目, 以体型较大的嘴刺目(Enoplida)所占比例最高(54%), 其次是色矛目(Chromadorida)(17%)和单宫目(Monhysterida)(15%)。这一方面反映了取样生境的线虫分类组成, 也与个体大小对 DNA条形码的难易程度影响有关(张志南等, 2017)。
随着高通量测序等分子生物学技术的快速发展和以DNA分子鉴定为手段进行小型底栖生物多样性研究的意识增强, 在我国的小型底栖生物分类学和生态学研究中运用分子生物学手段将逐步变成一种趋势和常态(史本泽, 2016; 张志南等, 2017)。
3.10 海洋生态动力学——水层-底栖耦合模式
养虾池水层-底栖耦合模式。我国水层-底栖生态系统耦合模式的研究, 首先应用于养虾池生态演变过程(翟雪梅等, 1998)。模式中浮游和底栖生态系统的耦合主要是通过 POC沉降、小型底栖生物和微生物的摄食消化、沉积物营养盐的滤出和生物扰动等过程来实现。吴增茂等建立了胶州湾生态系统的水层-底栖耦合生态模式, 并成功地模拟了胶州湾 13个生态度量的季节变化, 这是我国第一个水层-底栖耦合生态系统模型, 也是小型底栖生物作为一个重要的连接首次进入水层-底栖耦合模型(吴增茂等, 1999;余光耀等, 1999)。Zhang等(2006)在胶州湾模式基础上建立了渤海多箱模型, 将渤海分为6个区域, 对其水层-底栖耦合生态系统动力学特征及其演变进行了模拟分析, 结果显示, 13%的光合作用初级生产进入水层主食物链, 20%转移至底栖系统, 44%为植物呼吸所消耗, 其余转化为 POC形式存在于水体中, 其中小型底栖生物是联结水层和底栖系统的一个关键变量。以上结果为渤海水层-底栖耦合网络模型和底栖碎屑食物链各组分的研究提供了依据和支持。
3.11 高层次人才培养
20世纪1986—1987年间中国海洋大学首次为硕士生开设了线虫学专题(重点是自由生活海洋线虫形态分类学和生态学)和小型底栖生态学及海洋线虫分类学的硕士生学位课。据四大系统20余个单位的不完全统计, 至今我国共培养小型底栖生物方向硕士生90余名, 博士生20名, 已出站和在站博士后4名。科研成果共获得省部级奖励 6项。1986年正式设立的国家自然科学基金, 多个重大项目的实施和系列卓有成效的国际合作, 极大地推动了“小型底栖生物学”这一学科在我国的确立和发展(张志南等, 2017)。
4 展望——应重点关注的方向
4.1 海洋底栖生态系统对全球变化的响应
海洋生态系统在结构和功能上, 从一种局域的稳态向另一种局域稳态的突然转变称作状态转移(regime shifts)。驱动状态转移有三种过程: (1)非生物过程, 如全球变暖(温度升高)及大气和海洋大尺度的振荡(oscillation); (2)生物过程由过度捕捞导致的食物网重建和关键种的动态变化, 如上升流系统沙丁鱼和鱼种群的更替; (3)生境结构的改变, 可由自然的非生物事件引起, 如飓风, 或由人类活动导致, 如珊瑚礁的炸鱼及由于红树林的破坏导致育幼场的丢失,外来种的引入是人类活动影响的另一个例子(Deyounget al, 2004; Mantua, 2004; Kraberget al,2011)。
以上三种驱动过程往往综合在一起, 难以分开。以上过程及其响应的空间尺度依生态系统类型而异。可从几千米(如珊瑚礁)到几千千米的海盆尺度(如北太平洋)。然而, 以上三种驱动过程和响应过程的时间尺度从数年到跨越数个年代际。当今, 国际上已确定的状态转移, 大多集中在鱼类、浮游动物和浮游植物。大型底栖动物的状态转移研究历史尚短, 小型底栖动物的状态转移研究则刚刚起步。
状态转移的研究需要多学科交叉和大数据, 特别是长时间系列数据(至少跨越 4个年代际)的支持,为了探测系统内部的机制, 往往需要跨越几个营养级。借助多元统计分析和多种模型, 探测诊断已经出现的状态转移, 并预测未来可能发生的状态转移, 为生态系统健康评估和管理提供科学依据是当今全球变化和生态响应热点之一(Mantua, 2004)。
4.2 海洋底栖生物多样性——生态功能的考虑
从功能多样性的角度, 应考虑海洋底栖生物多样性在生物地球化学循环中的作用, 生态系统生产力和食物网结构。
海洋生物多样性与陆地生物多样性最大的区别在于, 他与生态学过程紧密联系在一起。这是因为海洋是三维立体空间, 物种的生命周期短、周转率高,初级生产力和物质的分解过程由微型生物驱动, 高的门类多样性, 以及沿岸系统生态学过程, 离岸的生物海洋学过程和野外的实验生态学之间立界分明。在海洋生物多样性的研究中, 既要考虑水层系统的多样性, 又需考虑底栖生态系统, 包括沉积物环境的多样性。因为两者是借助能流、物质循环和生活史紧密联系在一起的(张志南, 2000)。
除了时间尺度和空间尺度, 还应考虑生物结构的尺度, 即从基因一直到生态系统。生物多样性的计算不应只停留在一个营养级上, 应开展多个营养级功能多样性的比较。海洋生物多样性的生态功能考虑已成为当今海洋生物多样性和生物海洋学的一个研究热点(Heipet al, 2003)。
4.3 深海化能合成生态系统(ChEss)
ChEss包括热液、冷泉、鲸骨, 以及其他高度还原生境形成的生态系统, 是当今国际海洋生物普查计划的现场研究项目之一。由于对极端条件高压、低温、缺氧或无氧环境的适应, 该生态系统的研究对全球生物地理学、物种形成、深海资源开发和生命起源探索, 具有重大理论意义和潜在的应用价值。
自 1977年热液生物群落发现后, 在全球海洋中系统100多个热液口生物群落中已发现超过550种大型底栖动物, 其特点是生物量非常高, 但生物多样性很低且具有极高比例的地方种(Vanreuselet al, 2010;Levinet al, 2016)。
2010年在比利时根特大学召开的ChEss研讨会,搭建了一个研究还原条件与小型底栖生物的平台RoMeio, 以总结和推动这一年轻的科学领域。已有的成果显示, 大型和小型底栖动物的数量与热液排出量呈负相关, 小型动物与大型动物的数量呈负相关,暗示与摄食有关。深海自由生活海洋已报导近700种,而严格意义的ChEss生境中记载的线虫不足 100种,线虫丰度往往超过邻近正常海底区, 但往往单一种占优势, 属的丰度与热液和冷渗的溢出呈负相关; 冷渗中出现的线虫种和属, 类似于栖息在深水处的浅水种, 而非典型的深海类群, 在热液口线虫区系与邻近海底有高度的相似性, 说明是对极端条件适应的结果。当前, 除了进行小型底栖生物的分类, 还应联系特定环境条件开展还原生态系统的整体研究(Vanreuselet al, 2010)。
活动性和非活动性化能合成系统之间的过渡带(ecotone)的研究, 是当前一个研究热点。过渡带广泛分布, 有特有的环境条件和群落, 又充当能量、水、营养盐、颗粒物、有机质和生物等转移的通道, 有可能成为遗传和分类多样性热点和新的演化中心(Levinet al, 2016)。
4.4 加强小型底栖生物的基础生物学研究
在当前应着重加强小型底栖生物两个主要类群,即海洋线虫和海洋猛水蚤的基础生物学研究, 包括传统的形态学研究, 加大分子生物学融合力度, 争取最短时间内摸清我国小型底栖生物的资源和数量,包括物种数、底栖食物网的定量研究、指示种、代表种、模式种的室内培养和种群动态。尽早地纳入底栖生态系统服务过程, 适应国家需求, 为海洋生态系统健康评估, 全球变化响应和生态系统管理提供依据(张志南等, 2017)。
4.5 探讨分选程序的自动化和分类鉴定的智能化
小型底栖生物的分选过程和到种的分类鉴定是困扰小型底栖生物深层研究的两大瓶颈。探索使用高新光电技术、图型识别、分子标记、人工智能和互联网技术, 逐步实现底栖生态和生物海洋学的野外及室内与其他学科的同步或准同步现场观测和实时数据共享, 是未来国内外重点攻克的难题之一。
4.6 加快数据库平台的建立
基于国际条形码 BOLD数据库平台, 尽快构建和完备中国海自由生活海洋线虫和底栖猛水蚤 DNA条形码参考数据库(张志南等, 2017)。
4.7 加速高层次人才培养
充分利用国际合作及有关平台, 加大拓宽有关三大洋、南极、北极国际航次参与度, 扩展观测研究的时空尺度, 加大多学科交叉力度, 特别是小型底栖生物分类学、生态学、分子生物学与现代高新技术,微生物过程和生物地化过程的结合, 加速高层次人才培养。
4.8 重视科普教育和公益宣传
开展各种形式的科普教育和公益性活动, 特别加强青少年学生的海洋意识, 加深对海洋生物、海底生命过程的了解和探索, 壮大海洋底栖生物研究的潜在后备力量。
致谢 本文首在2016年11月海口中国海洋湖沼学会海洋底栖生物学分会成立大会上宣读。感谢徐奎栋教授的鼓励和支持, 感谢中国海洋大学海洋生命学院硕士研究生生高春子同学协助修改。
马 林, 2011.中国海底栖桡足类的分类学研究.青岛: 中国科学院研究生院(海洋研究所)博士学位论文
王 摆, 汝少国, 于子山等, 2007.自由生活海洋线虫Chromadorinasp.的生活史研究.水生生物学报, 31(5):751—754
王小谷, 周亚东, 张东声等, 2013.2005年夏季东太平洋中国多金属结核区小型底栖生物研究.生态学报, 33(2):492—500
王诗红, 张志南, 1994.养虾池海洋线虫的培养及生活史的初步研究.青岛海洋大学学报, 24, (supp): 206—213
王彦国, 2008.台湾海峡及其邻近海域小型底栖动物生态学研究.厦门: 厦门大学硕士学位论文
王家栋, 类彦立, 徐奎栋等, 2011.黄海冷水团及周边海域夏初小型底栖动物现存量及空间分布研究.海洋与湖沼,42(3): 359—366
韦晓慧, 慕芳红, 孙艳桃等, 2014.海洋酸化条件下铜、镉对日本虎斑猛水蚤的急性毒性效应.生态学报, 34(14):3879—3884
邓 可, 张志南, 黄 勇等, 2005.南黄海典型站位底栖动物粒径谱及其应用.中国海洋大学学报, 35(6): 1005—1010
叶树明, 高少佳, 潘 佚等, 2006.激光诱导荧光检测技术在小型底栖生物分离系统中应用的研究.科技通报, 22(3):395—399, 425
叶树明, 潘 佚, 高少佳等, 2005.深海小型底栖生物标本的自动分离仪设计.江南大学学报(自然科学版), 4(3):240—243
史本泽, 2016.不同生境中海洋线虫分类及小型底栖生物群落结构研究.北京: 中国科学院大学博士学位论文
朱丽岩, 2005.几种环境内分泌干扰物对青岛近海常见海洋动物的毒性效应.青岛: 中国海洋大学博士学位论文
华 尔, 李 佳, 董 洁等, 2012.砂质潮间带自由生活海洋线虫对缺氧的响应——微型受控生态系研究.生态学报,32(13): 3975—3986
华 尔, 张志南, 2009.黄河口邻近海域底栖动物粒径谱研究.中国海洋大学学报, 39(5): 971—978
华 尔, 张志南, 张 艳, 2005.长江口及邻近海域小型底栖生物丰度和生物量.生态学报, 25(9): 2234—2242
华 尔, 阎鹏旭, 贺阳阳等, 2015.大型绿藻暴发对砂质潮间带小型底栖动物群落的影响.海洋湖沼通报, (3): 45—63
刘晓收, 许 嫚, 张敬怀等, 2014.南海北部深海小型底栖动物丰度和生物量.热带海洋学报, 33(2): 52—59许永香, 修瑞琴, 郑 静, 1992.八种毒物对Nitocra spinipes的毒性研究.生态科学, (2): 64—66
孙 松, 2012.中国区域海洋学: 生物海洋学.北京: 海洋出版社
孙艳桃, 慕芳红, 韦晓慧等, 2014.海水酸化对日角猛水蚤和日本虎斑猛水蚤摄食率的影响.海洋湖沼通报, (3):105—111
杨俊毅, 王春生, 刘镇盛等, 2005.热带北太平洋深海小型底栖生物大尺度空间分布.海洋学研究, 23(3): 23—29
吴秀芹, 徐奎栋, 于子山等, 2010.2008年浒苔大暴发末期黄海小型底栖动物现存量及空间分布.应用生态学报, 21(8):2140—2147
吴增茂, 余光耀, 张志南等, 1999.胶州湾北部水层生态动力学模型与模拟Ⅱ.胶州湾北部水层生态动力学的模拟研究.青岛海洋大学学报, 29(3): 429—435
邹丽珍, 2006.中国合同区小型底栖动物及其深海沉积物中18S rDNA基因多样性研究.厦门: 国家海洋局第三海洋研究所硕士学位论文
沈锡权, 2005.青岛沿岸自由生活海洋线虫18S核糖体RNA基因扩增及序列变异分析.青岛: 中国海洋大学硕士学位论文
沈锡权, 杨官品, 廖梅杰, 2005.单条固定线虫基因组DNA提取及18S rRNA基因PCR扩增.海洋科学, 29(5): 33—36
张 艳, 2006.南黄海小型底栖生物群落结构与多样性的研究.青岛: 中国海洋大学博士学位论文
张 艳, 张志南, 邓 可等, 2005.青岛岩礁附植小型底栖动物ATP含量的研究.中国海洋大学学报, 35(5): 799—806
张达娟, 李少菁, 王桂忠等, 2011.二氧化碳酸化和盐酸酸化对几种桡足类的急性毒性比较.厦门大学学报(自然科学版), 50(3): 631—636
张志南, 1991.秦皇岛砂滩海洋线虫的数量研究.青岛海洋大学学报, 21(1): 63—75
张志南, 2000.水层-底栖耦合生态动力学研究的某些进展.青岛海洋大学学报, 30(1): 115—122
张志南, 刘素美, 周 红等, 2017.渤海底栖生物次级生产力与生物多样性.北京: 科学出版社, 409
张志南, 刘晓收, Boucher G等, 2005.底栖生物生产力现场测试系统(BCSPM)的建立及其初步实验结果.中国海洋大学学报, 35(2): 277—282
张志南, 李永贵, 图立红等, 1989.黄河口水下三角洲及其邻近水域小型底栖动物的初步研究.海洋与湖沼, 20(3):197—208
张志南, 谷 峰, 于子山, 1990a.黄河口水下三角洲海洋线虫空间分布的研究.海洋与湖沼, 21(1): 11—19
张志南, 林 霞, 于子山, 1994.大连石槽岩滩附植小型动物的初步研究.青岛海洋大学学报, 24(3): 373—383
张志南, 林岿旋, 周 红等, 2004b.东、黄海春秋季小型底栖生物丰度和生物量研究.生态学报, 24(5): 997—1005
张志南, 周 宇, 韩 洁等, 1999.生物扰动实验系统(AFS)的基本结构和工作原理.海洋科学, 23(6): 28—30
张志南, 周 宇, 韩 洁等, 2000a.应用生物扰动实验系统(Annular Flux System)研究双壳类生物沉降作用.青岛海洋大学学报, 30(2): 270—276
张志南, 周 红, 2003.自由生活海洋线虫的系统分类学.青岛海洋大学学报, 33(6): 891—900
张志南, 周 红, 2004a.国际小型底栖生物研究的某些进展.中国海洋大学学报, 34(5): 799—806
张志南, 周 红, 2007.小型底栖生物调查, 海洋生物卷, 海洋调查规范2007 (GB-国家标准).北京: 国家标准局出版社
张志南, 周 红, 郭玉清等, 2000b.渤海海洋线虫生物多样性的比较研究.中国学术期刊文摘, 6(1): 93—95
张志南, 周 红, 郭玉清等, 2001a.黄河口水下三角洲及其邻近水域线虫群落结构的比较研究.海洋与湖沼, 32(4):436—444
张志南, 周 红, 慕芳红, 2001b.渤海线虫群落的多样性及中性模型分析.生态学报, 21(11): 1808—1814
张志南, 党宏月, 于子山, 1993.青岛湾有机质污染带小型底栖生物群落的研究.青岛海洋大学学报, 23(1): 83—91
张志南, 钱国珍, 1990b.小型底栖生物取样方法的研究.海洋湖沼通报, (4): 37—42
张志南, 慕芳红, 于子山等, 2002.南黄海鱼产卵场小型底栖生物的丰度和生物量.青岛海洋大学学报, 32(2):251—258
张青田, 王新华, 胡桂坤, 2012.底栖线虫和桡足类丰度比与环境的关系分析.南开大学学报(自然科学版), 45(5):52—57
张青田, 胡桂坤, 2008.小型底栖生物在海洋生态监测中的应用.海洋信息, (4): 28—29
陈世杰, 1988.厦门港尖额真猛水蚤室内培养的研究.水产学报, 12(4): 339—345
陈珂, 2004.三种海洋桡足类摄食、生殖和发育的研究.青岛:中国海洋大学硕士学位论文
陈海燕, 周 红, 张志南等, 2011.青岛岩礁海藻附植动物的区系组成和季节动态.中国海洋大学学报, 41(7—8):85—90
林岿璇, 张志南, 王睿照, 2004.东、黄海典型站位底栖动物粒径谱研究.生态学报, 24(2): 241—245
林岿璇, 张志南, 韩 洁, 2003.南麂列岛海洋自然保护区潮间带小型生物初步研究.青岛海洋大学学报, 33(2):219—225
季如宝, 张志南, 1994.14C示踪法测定养虾池小型底栖动物对底栖硅藻的摄食.青岛海洋大学学报, 24(8): 199—205
余光耀, 吴增茂, 张志南等, 1999.胶州湾北部水层生态动力学模型与模拟Ⅰ.胶州湾北部水层生态动力学模型.青岛海洋大学学报, 29(3): 421—428
修瑞琴, 傅迎春, 许永香, 1995.硒与氟离子对美丽猛水蚤的联合毒性.环境科学, 16(1): 26—28
党宏月, 黄 勃, 张志南, 1996.青岛湾有机质污染潮间带底栖生物研究Ⅱ.小型底栖动物生态特点.海洋科学集刊,37: 91—102
郭玉清, 2008.厦门凤林红树林湿地自由生活海洋线虫群落的研究.海洋学报, 30(4): 147—153
郭玉清, 张志南, 慕芳红, 2002.渤海海洋线虫与底栖桡足类数量之比的应用研究.海洋科学, 26(12): 27—31
黄 勇, 2005.南黄海小型底栖生物生态学和海洋线虫分类学研究.青岛: 中国海洋大学博士学位论文
黄宗国, 林 茂, 2012.中国海洋物种和图集上卷: 中国海洋物种多样性(上册和下册).北京: 海洋出版社
黄宗国主编, 2008.中国海洋生物种类与分布(增订版).北京:海洋出版社, 1191
曾艳艺, 黄小平, 2011.日本虎斑猛水蚤的重金属急性毒性及其作为标准测试生物的潜在意义.生态毒理学报, 6(2):182—188
谢 礼, 林 霞, 朱艺峰, 2012.象山港国华宁海电厂附近海域小型底栖动物的群落结构.生态科学, 31(2): 167—172
蔡立哲, 2015.深圳湾底栖动物生态学.厦门: 厦门大学出版社, 303
蔡立哲, 厉红梅, 邹朝中, 2000a.厦门钟宅泥滩海洋线虫群落的种类组成及其多样性.厦门大学学报(自然科学版),39(5): 669—675
蔡立哲, 厉红梅, 邹朝中, 2000b.深圳河口福田泥滩海洋线虫的种类组成及季节变化.生物多样性, 8(4): 385—390
翟雪梅, 张志南, 1998.虾池生态系能流结构分析.青岛海洋大学学报, 28(2): 275—282
Andrássy I, 1976.Evolution as A Basis for the Systematization of Nematodes.London: Pitman Publishing, 288
Appeltans W, Ahyong S T, Anderson Get al, 2012.The magnitude of global marine species diversity.Current Biology, 22(23): 2189—2202
Cao Z Q, Mu F H, Wei X Het al, 2015.Influence of CO2-induced seawater acidification on the development and lifetime reproduction ofTigriopus japonicusMori, 1938.Journal of Natural History, 49(45—48): 2813—2826
Chen H L, Li B, Hu J Bet al, 2007.Effects ofSpartina alterniflorainvasion on benthic nematode communities in the Yangtze Estuary.Marine Ecology Progress Series, 336:99—110
Chen Y Z, Guo Y Q, 2014.Three new species of free-living marine nematodes from East China Sea.Zootaxa, 3841(1):117—126
Chen Y Z, Guo Y Q, 2015.Three new and two known free-living marine nematode species of the family Ironidae from the East China Sea.Zootaxa, 4018(2): 151—175
Coull B C, Chandler G T, 1992.Pollution and meiofauna: field,laboratory, and mesocosm studies.Oceanography Marine Biology, 30: 191—271
deYoung B, Harris R, Alheit Jet al, 2004.Detecting regime shifts in the ocean: data considerations.Progress in Oceanography,60(2—4): 143—164
Eleftheriou A, McIntyre A, 2005.Methods for the Study of Marine Benthos.3rd ed.Chichester, West Sussex, UK:Blackwell Publishing
Fu S J, Cai L Z, Boucher Get al, 2013.Two newRichtersiaspecies from the northern Beibu Gulf, China.Journal of Natural History, 47(29—30): 1921—1931
Gee J M, Mu F H, 2000.A new genus of Cletodidae (Copepoda;Harpacticoida) from the Bohai Sea, China.Journal of Natural History, 34(6): 809—822
Giere O, 2009.Meiobenthology.Berlin, Heidelberg: Springer,327
Guilini K, Bezerra T N, Eisendle-Flöckner Uet al, 2016.NeMys:world database of free-living marine nematodes.[2016-03-28].http://nemys.ugent.be
Guo Y Q, Chen Y Z, Liu M D, 2016.Metadesmolaimus zhanggisp.nov.(Nematoda: Xyalidae) from East China Sea, with a pictorial key toMetadesmolaimusspecies.Cahiers de Biologie Marine, 57(1): 73—79
Guo Y Q, Huang D Y, Chen Y Zet al, 2015.Two new free-living nematode species ofSetosabatieria(Comesomatidea) from the East China Sea and the Chukchi Sea.Journal of Natural History, 49(33—34): 2021—2033
Guo Y Q, Warwick R M, 2001.Three new species of free-living nematodes from the Bohai Sea, China.Journal of Natural History, 35(11): 1575—1586
Guo Y Q, Warwick R M, Zhang Z Net al, 2002.Freeliving marine nematodes as a pollution indicator of Bohai Sea.Journal of Environmental Science, 14(4): 558—562
Heip C H R, Hummel H, Van Avesaath P Het al, 2003.Biodiversity of Coastal Marine Ecosystems: A Functional Approach to Coastal Marine Biodiversity: Book of Abstracts.Renesse, the Netherlands: Netherlands Institute of Ecology,Centre for Estuarine and Marine Ecology
Heip C H R, Vincx M, Vranken G, 1985.The ecology of marine nematodes.Oceanography and Marine Biology: An Annual Review, 23: 399—489
Higgins R P, Thiel H, 1988.Introduction to the Study of Meiofauna.Washington, D.C.: Smithsonian Press, 488
Hope W D, Zhang Z N, 1995.New nematodes from the Yellow Sea,Hopperia hexadentatan.sp.andCervonema deltensisn.sp.(Chromadorida: Comesomatidae), with observations on morphology and systematics.Invertebrate Biology, 114(2):119—138
Hua E, Zhang Z N, 2007.Four newly recorded free-living marine nematodes (Comesomatidae) from the East China Sea.Journal of Ocean University of China, 6(1): 26—32
Hua E, Zhang Z N, Warwick R Met al, 2013.Pattern of benthic biomass size spectra from shallow waters in the East China Seas.Marine Biology, 160(7): 1723—1736
Hua E, Zhang Z N, Yu Z Set al, 2010.Preliminary study on the immediate response of the nematode community to Typhoon Soudelor.Deep Sea Research Part II: Topical Studies in Oceanography, 57(11—12): 1064—1070
Huang Y, Cheng B, 2012.Three new free-living marine nematode species of the genusMicoletzkyia(Phanodermatidae) from China Sea.Journal of the Marine Biological Association of the United Kingdom, 92(5): 941—945
Huang Y, Gao Q, 2016.Two new species of Chromadoridae(Chromadorida: Nematoda) from the East China Sea.Zootaxa, 4144(1): 89—100
Huang Y, Wang H X, 2015.Review ofOnyxCobb (Nematoda:Desmodoridae) with description of two new species from the Yellow Sea, China.Journal of the Marine Biological Association of the United Kingdom, 95(6): 1127—1132
Huang Y, Xu K D, 2013.Two new species of the genusParacyatholaimusMicoletzky (Nematoda: Cyatholaimidae)from the Yellow Sea.Journal of Natural History, 47(21—22):1381—1392
Huang Y, Zhang Z N, 2005.Three new species of the genusBelbolla(Nematoda: Enoplida: Enchelidiidae) from the Yellow Sea, China.Journal of Natural History, 39(20):1689—1703
Huang Y, Zhang Z N, 2006a.A new genus and three new species of free-living marine nematodes from the Yellow Sea, China.Journal of Natural History, 40(1—2): 5—16
Huang Y, Zhang Z N, 2006b.Five new records of free-living marine nematodes in the Yellow Sea.Journal of Ocean University of China, 5(1): 29—34
Huang Y, Zhang Z N, 2006c.New species of free-living marine nematodes from the Yellow Sea, China.Journal of the Marine Biological Association of the United Kingdom, 86(2):271—281
Huang Y, Zhang Z N, 2006d.Two new species of free-living marine nematodes (Trichotheristus articulatussp.n.andLeptolaimoides punctatussp.n.) from the Yellow Sea, China.Russian Journal of Nematology, 14(1): 43—50
Huang Y, Zhang Z N, 2007.One new species of free-living marine nematodes (Enoplida, Anticomidae,Cephalanticoma)from the Huanghai Sea.Acta Oceanologica Sinica, 26(3):84—89
Huang Y, Zhang Z N, 2009.Two new species of Enoplida(Nematoda) from the Yellow Sea, China.Journal of Natural History, 43(17—18): 1083—1092
Huang Y, Zhang Z N, 2010.Three new species ofDichromadora(Nematoda: Chromadorida: Chromadoridae) from the Yellow Sea, China.Journal of Natural History, 44(9—10): 545—558 Hulings N C (Editor), 1971.Proceedings of the First International Conference on Meiofauna.Smithsonian Contributions to Zoology 76.Washington D.C.: Smithsonian Institution Press, 1—205
Hulings N C, Gray J S (Editors), 1971.A Manual for the Study of Meiofauna.Smithsonian Contributions to Zoology 78.Washington D.C.: Smithsonian Institution Press, 83
Huys R, Gee J M, Moore C Get al, 1996.Marine and brackish water harpacticoid copepods Part 1: keys and notes for identification of the species.In: Barnes R S K, Crothers J H.Synopses of the British Fauna, New Series 51.Shrewsbury:Field Studies Council, 1—352
Huys R, Mu F H, 2008.Description of a new species ofOnychostenheliaItô (Copepoda, Harpacticoida, Miraciidae)from the Bohai Sea, China.Zootaxa, 1706: 51—68
Jiang W, Huang Y, 2015a.Paragnomoxyalagen.nov.(Xyalidae,Monhysterida, Nematoda) from the East China Sea.Zootax,4039(3): 467—474
Kraberg A C, Wasmund N, Vanaverbeke Jet al, 2011.Regime shifts in the marine environment: the scientific basis and political context.Marine Pollution Bulletin, 62(1): 7—20
Levin L A, Baco A R, Bowden D Aet al, 2016.Hydrothermal vents and methane seeps: rethinking the sphere of influence.Frontiers in Marine Science, 3: 72
Li Y X, Guo Y Q, 2016a.Free living marine nematodes of the genusParodontophora(Axonolaimidae) from the East China Sea, with descriptions of five new species and a pictorial key.Zootaxa, 4109(4): 401—427
Li Y X, Guo Y Q, 2016b.Two new free-living marine nematode species of the genusAnoplostoma(Anoplostomatidae) from the mangrove habitats of Xiamen Bay, East China Sea.Journal of Ocean University of China, 15(1): 11—18
Liu X S, Cheung S G, Shin P K S, 2011.Response of meiofaunal and nematode communities to sewage pollution abatement: a field transplantation experiment.Chinese Journal of Oceanology and Limnology, 29(6): 1174—1185
Liu X S, Zhang Z N, Huang Y, 2005.Abundance and biomass of meiobenthos in the spawning ground of anchovy (Engraulis japanicus) in the Southern Huanghai Sea.Acta Oceanologica Sinica, 24(3): 94—104
Liu X S, Zhang Z N, Huang Y, 2007.Sublittoral meiofauna with particular reference to nematodes in the southern Yellow Sea,China.Estuarine, Coastal and Shelf Science, 71(3—4):616—628
Lv Y, Zhang W D, Gao Yet al, 2011.Preliminary study on responses of marine nematode community to crude oil contamination in intertidal zone of Bathing Beach, Dalian.Marine Pollution Bulletin, 62(12): 2700—2706
Ma L, Li X Z, 2011.Delavalia qingdaoensissp.nov.(Harpacticoida, Miraciidae), a new copepod species from Jiaozhou Bay, Yellow Sea.Crustaceana, 84(9): 1085—1097 Mantua N, 2004.Methods for detecting regime shifts in large marine ecosystems: a review with approaches applied to North Pacific data.Progress in Oceanography, 60(2—4):165—182
Mare M F, 1942.A study of a marine benthic community with special reference to the micro-organisms.Journal of the Marine Biological Association of the United Kingdom, 25(3):517—554
McIntyre A D, 1969.Ecology of marine meiobenthos.Biological Reviews, 44(2): 245—288
Montagna P A, 1984.In situ measurement of meiobenthic grazing rates on sediment bacteria and edaphic diatoms.Marine Ecology Progress Series, 18: 119—130
Montagna P A, Yoon W B, 1991.The effect of freshwater inflow on meiofaunal consumption of sediment bacteria and microphytobenthos in San Antonio Bay, Texas, U.S.A.Estuarine, Coastal and Shelf Science, 33(6): 529—547
Mu F H, Gee J M, 2000.Two new species ofBulbamphiascus(Copepoda: Harpacticoida: Diosaccidae) and a related new genus, from the Bohai Sea, China.Cahiers de Biologie Marine, 41(2): 103—135
Mu F H, Huys R, 2002a.New species ofStenhilia(Copepoda,Harpacticoida, Diosaccidae) from the Bohai Sea (China)with notes on subgeneric division and phylogenetic relationships.Cahiers de Biologie Marine, 43: 179—206
Mu F H, Huys R, 2004.Canuellidae (Copepoda, Harpacticoida)from the Bohai Sea, China.Journal of Natural History, 38(1):1—36
Mu F H, Somerfield P J, Warwick R Met al, 2002b.Large-scale spatial patterns in the community structure of benthic harpacticoid copepods in the Bohai Sea, China.The Raffles Bulletin of Zoology, 50(1): 17—26
Nicholls A G, 1935.Copepods from the interstitial fauna of a sandy beach.Journal of the Marine Biological Association of the United Kingdom, 20(2): 379—405
Platt H M, Zhang Z N, 1982.New species of marine nematodes from Loch Ewe, Scotland.Bulletin of the British Museum(Natural History) Zoology, 42(4): 227—246
Remane A, 1933.Verteilung und organisation der benthonischen mikrofauna der kieler bucht.Wiss Meeresunters Abt Kiel,21(2): 163—221
Schratzberger M, Ingels J, 2016.Meiofauna matters: the roles of meiofauna in benthic ecosystems.Draft Book of Abstracts for 16th International Meiofauna Conference, July 3—8,2016.Heraklion, Greece: Hellenic Centre for Marine Research, Biology Department, University of Crete (UoC),Department of Marine Studies, University of Piraeus (UP),International Association of Meiobenthologists (IAM)
Shen X Q, Yang G P, Liu Y J, 2007.Nematode diversity of Qingdao coast inferred from the 18S ribosomal RNA gene sequence analysis.Journal of Ocean University of China,6(2): 132—136
Sun J, Huang Y, 2016.A new genus of free-living nematodes(Enoplida: Enchelidiidae) from the South China Sea.Cahiers de Biologie Marine, 57(1): 51—56
Sun X Y, Zhou H, Hua Eet al, 2014.Meiofauna and its sedimentary environment as an integrated indication of anthropogenic disturbance to sandy beach ecosystems.Marine Pollution Bulletin, 88(1—2): 260—267
Tietjen J H, Lee J J, 1984.The use of free-living nematodes as a bioassay for estuarine sediments.Marine Environmental Research, 11(4): 233—251
Vanreusel A, De Groote A, Gollner Set al, 2010.Ecology and biogeography of free-living nematodes associated with chemosynthetic environments in the deep sea: a review.PLoS One, 5(8): e12449
Wang J N, Zhou H, Zhang Z Net al, 2011.Effects of organic enrichment on sandy beach meiofauna: a laboratory microcosm experiment.Journal of Ocean University of China, 10(3): 246—254
Warwick R M, 2016.Evolving paradigms: the history of meiofaunal research.Draft book of abstracts for 16th International Meiofauna Conference, July 3—8, 2016.Heraklion, Greces: Hellenic Centre for Marine Research,Biology Department, University of Crete (UoC), Department of Marine Studies, University of Piraeus (UP), International Association of Meiobenthologists (IAM)
Wells J B J, 2007.An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea).Zootaxa, 1568:1—872
Wu J H, Chen H L, Zhang Y Z, 2016.Latitudinal variation in nematode diversity and ecological roles along the Chinese coast.Ecology and Evolution, 6(22): 8018—8027
Zhang X L, Wu Z M, Li Jet al, 2006.Modeling study of seasonal variation of the pelagic-benthic ecosystem characteristics of the Bohai Sea.Journal of Ocean University of China, 5(1):21—28
Zhang Y, Zhang Z N, 2006b.Two new species of the genusElzalia(Nematoda: Monhysterida: Xyalidae) from the Yellow Sea, China.Journal of the Marine Biological Association of the United Kingdom, 86(5): 1047—1056
Zhang Z N, 1983.Three new species of free-living marine nematodes from a sublittoral station in Firemore Bay,Scotland.Cahiers de Biologie Marine, 24(2): 219—229
Zhang Z N, 1990.A new species of the genusThalassironusde Man, 1889 (Nematoda, Adenophora, Ironidae) from the Bohai Sea, China.Journal of Ocean University of Qingdao,20(3): 103—108
Zhang Z N, 1991.Two new species of marine nematodes from the Bohai Sea, China.Journal of Ocean University of Qingdao, 21(2): 49—60
Zhang Z N, 1992.Two new species of the genusDorylaimopsisDitlevsen, 1918 (Nematoda: Adenophora, Comesomatidae)from the Bohai Sea, China.Chinese Journal of Oceanology and Limnology, 10(1): 31—39
Zhang Z N, 2005.Three new species of free-living marine nematodes from the Bohai Sea and Yellow Sea, China.Journal of Natural History, 39(23): 2109—2123
Zhang Z N, Huang Y, 2005.One new species and two new records of free-living marine nematodes from the Huanghai Sea.Acta Oceanologica Sinica, 24(4): 91—97
Zhang Z N, Ji R B, 1994.The first record ofTerschellingia longicaudataDe Man 1907 in prawn pond.Journal of Ocean University of Qingdao, 24(S83): 214—221
Zhang Z N, Platt H M, 1983.New species of marine nematodes from Qingdao, China.Bulletin of the British Museum(Natural History) Zoology, 45(5): 253—261
Zhang Z N, Zhou H, 2012.Enoplus taipingensis, a new species of marine nematode from the rocky intertidal seaweeds in the Taiping Bay, Qingdao.Acta Oceanologica Sinica, 31(2):102—108
Zhang Z N, Zhou H, Nightingale C L, 1997.Phytal meiofauna of a rocky shore at the Cape d’Aguilar marine reserve, Hong Kong.In: Morton B.The Marine Flora and Fauna of Hong Kong and Southern China IV.Proceeding of the Eighth International Marine Biological Workshop, Hong Kong.Hong Kong: Hong Kong University Press, 205—217
MEIOFAUNA STUDY FOR THE FORTY YEARS IN CHINA——PROGRESS AND PROSPECT
ZHANG Zhi-Nan, ZHOU Hong, HUA Er, MU Fang-Hong, LIU Xiao-Shou, YU Zi-Shan
(College of Marine Life Sciences,Ocean University of China,Qingdao266003,China)
Based on the data collected from the 16thInternational Meiofauna Conference, Heraklion, Greece, 2016,together with the available information, the history, present status, trends and some important progress are in reviews,which include the methodology for meiofauna, taxonomy of free-living marine nematodes and benthic copepods,distribution of abundance and biomass, effect ofSpartina alterniflorainvasion on nematode communities, size spectrum and secondary production, feeding ecology, culture in laboratory and life cycle, environmental monitoring, molecular biology and systematic evolution of marine nematodes, benthic-pelagic coupling model and senior qualified personnel training.Some strategy and tactics to enhance the study in this field are also suggested
meiofauna; marine nematodes; biodiversity; benthic-pelagic coupling; global change
* 国家自然科学基金重点项目, 40730847号; 海洋公益性行业科研专项项目, 201505004号; 国家自然科学基金项目,41376146号。张志南, 教授, 博士生导师, E-mail: znzhang37@163.com
2017-01-25, 收修改稿日期: 2017-03-29
猜你喜欢
今日农业(2022年14期)2022-09-15
中成药(2018年2期)2018-05-09
琴童(2017年7期)2017-07-31
小学科学(2017年5期)2017-05-26
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
小学阅读指南·低年级版(2016年12期)2017-01-05
科技知识动漫(2016年7期)2016-07-29
天然产物研究与开发(2016年6期)2016-06-05
