
基于林层的典型中亚热带天然阔叶林树种组成与多样性*
2017-12-05 05:42:14马志波黄清麟庄崇洋郑群瑞
林业科学 2017年10期
马志波 黄清麟 庄崇洋 郑群瑞 王 宏
(1.中国林业科学研究院资源信息研究所 国家林业局林业遥感与信息技术重点实验室 北京 100091; 2.建瓯万木林省级自然保护区管理处 建瓯 353105)
基于林层的典型中亚热带天然阔叶林树种组成与多样性*
马志波1黄清麟1庄崇洋1郑群瑞2王 宏1
(1.中国林业科学研究院资源信息研究所 国家林业局林业遥感与信息技术重点实验室 北京 100091; 2.建瓯万木林省级自然保护区管理处 建瓯 353105)
【目的】 分析各林层物种组成及多样性,探讨各林层之间的联系和群落结构的形成机制,以期为深入了解典型森林群落组成成分在垂直空间中的分布规律、揭示生物多样性维持机制提供依据。【方法】 在福建万木林自然保护区内设5块50 m×50 m标准地,使用最大受光面法自上而下将乔木层划分为第Ⅰ,Ⅱ和Ⅲ亚层,第I,II亚层属于受光层,第Ⅲ亚层为非受光层,采用常见物种多样性指标分析各层的组成与多样性,用相似性系数探讨各层之间的联系。【结果】 5块标准地均表现为由第Ⅰ亚层降至第Ⅲ亚层物种丰富度和多样性增加; 各样地相同亚层的物种组成及数量各不相同; 调查到的所有树种中,有35个树种可生长到第Ⅰ亚层,最大树高为30~40 m,但是各样地累计只有10个树种在乔木层的3个亚层连续分布,有20个树种(含4种灌木)能生长到第Ⅱ亚层,最大树高约为25 m,有30个树种(含13种灌木)只能分布在第Ⅲ亚层,最大树高为16~17 m; 受光层内的树种是乔木层的优势种和重要伴生树种,包括常绿树种木荷、浙江桂、香樟、刨花楠、观光木、福建含笑、乐东拟单性木兰和栲类等,也包括落叶树种枫香、拟赤杨、蓝果树和苦枥木; 最大高度限于非受光层即第Ⅲ亚层的常绿树种有野含笑、水丝梨、椤木石楠、虎皮楠和厚皮香,落叶树种有合欢、野柿和南岭黄檀; 标准地内的大部分灌木树种只分布在第Ⅲ亚层,但山矾、黄瑞木、弯蒴杜鹃和尖叶水丝梨除外,它们也可生长到第Ⅱ亚层; 第Ⅰ和第Ⅱ亚层的物种组成相似性大于第Ⅰ和第Ⅲ亚层及第Ⅰ和整个乔木层的相似性; 第Ⅰ亚层树种的更新储备主要分布于第Ⅱ亚层; 受光层的树种丰富度与多样性和非受光层有明显差异,二者的分界面也是空间生态位分化最显著的界面,而该界面正是最大受光面。【结论】 5块标准地乔木层内的优势种和重要伴生树种主要分布在受光层; 生态位分化最显著的位置是最大受光面,按最大受光面划分出的受光层和非受光层,在组成种的丰富性和多样性方面也具有明显的差别,受光层和非受光层分化对近自然经营中目标树的选择具有潜在的指导意义; 可生长到最高层的树种中,只有个别树种可在3个亚层连续分布,暗示受光层中的优势树种是不固定的,说明除了生态位分化,群落物种组成与丰富度还受随机因素影响。
天然常绿阔叶林; 中亚热带; 乔木层; 成层性; 相似性
亚热带常绿阔叶林带是我国东部湿润森林区域内最大的植被带(宋永昌, 2001),在提供林产品、生物多样性维持和水土保持等方面发挥着不可替代的作用。天然常绿阔叶林乔木层本身具有明显的成层性,可以划分为2~3个亚层(中国植被编委会, 1980; 王梅峒, 1988; 陈灵芝等, 1997; 祝燕等, 2008)。在天然常绿阔叶林典型群落或大型样地(宋永昌等, 2015)研究中,一般是把乔木层作为与灌木层、草本层并列的成分进行分析(贺金生等, 1998)。合理划分乔木层有助于进一步分析树种的生长特性、揭示群落演替过程中的种群动态、探讨不同垂直层次内的环境因子(光照、温度、湿度等)特征及各层植物的响应等。但是涉及亚热带森林群落乔木亚层特征的研究较少,而且分层方法主要依赖于研究者的经验(蚁伟民等, 1999; 樊后保, 2000; 叶万辉等, 2008; 赵丽娟等, 2013)。实际上,光是林木优先争夺的重要资源,高度在植物对光的竞争中起重要作用(Falsteretal., 2003; Chenetal., 2010)。在这个意义上,森林群落乔木层的成层性体现了林木高生长策略的差异,代表着不同林木在光竞争中的地位。近期有研究提出了主要基于林木接受光照差异的中亚热带天然阔叶林乔木层划分新方法“最大受光面法”(庄崇洋等, 2017),但是在群落特征研究中的应用尚未开展。
福建省建瓯市万木林自然保护区是我国设立最早的保护区之一,保护区内天然植被类型以典型中亚热带常绿阔叶林为主(陈灵芝等, 1997; 宋永昌, 2004),是研究中亚热带天然阔叶林生态学和林学特征的理想场所(吴承祯等, 1996; 刘茂松等, 1998; 黄清麟等, 1999; 2003; 金钊等, 2007; 裴广廷等, 2015),研究者已对保护区内森林群落的生物多样性有较充分的研究(吴承祯等, 1996; 朱锦懋等, 1997; 郑群瑞, 1999),对林层特征研究也有所涉及(黄清麟等, 1995; 2003; 庄崇洋等, 2017),但是对不同林层的多样性研究开展较少。
本研究选择福建省建瓯市万木林保护区内典型中亚热带天然阔叶林群落为对象,采用最大受光面法(庄崇洋等, 2017)划分乔木亚层、研究典型中亚热带天然阔叶森林群落各乔木亚层的组成与多样性及各亚层间的关系,以期为深入了解典型森林群落组成成分在垂直空间中的分布规律、揭示生物多样性的维持机制提供依据。
1 研究区概况
福建省建瓯万木林自然保护区(118°08′22″—118°09′23″ E, 27°02′28″—27°03′32″ N)是1956年林业部划定的全国首批19个天然林禁伐区之一,至今已经有600年以上有记载的保护史(何友钊, 1990),保护区森林植被的主体是中亚热带天然阔叶林即“典型常绿阔叶林”(陈灵芝等, 1997; 宋永昌, 2004),在人类活动频繁的低海拔丘陵山地(保护区海拔230~556 m)能得以保存,极其罕见。保护区位于闽北武夷山和鹫峰山之间,面积189 hm2,属中亚热带季风气候,年均气温18.7 ℃,年降水量1 670 mm,空气相对湿度81%,全年无霜期277天,全年日照时数1 812.7 h。土壤为花岗岩发育的红壤。
2 研究方法
2.1样地设置与调查
2015年冬季在保护区内选择较平坦的典型天然群落地段,设置5块50 m×50 m标准地,分别标记为1~5,均为典型中亚热带天然阔叶林(黄清麟, 1998; 黄清麟等, 1999; 2003),1号标准地为木荷(Schimasuperba)-光叶山矾(Symplocoslancifolia)群落,2号标准地为猴欢喜(Sloaneasinensis)-木荷群落,3号标准地为木荷-新木姜子(Neolitseaaurata)群落,4号标准地为木荷-浙江桂(Cinnamomumchekiangense)群落,5号标准地为浙江桂-木荷群落。1号标准地地位于下坡位,2和3号标准地位于相同坡面的中上坡位和中下坡位,4和5号标准地位于另一面坡的中上坡位和中下坡位。将每块标准地划分为25块10 m×10 m样地,对所有样地中胸径≥5 cm的木本植物进行每木调查,鉴定、记录种名,并准确测量胸径、树高(用测高杆和测高器测量)。调查过程中按最大受光面法(庄崇洋等, 2017)现场判断、记录林木的层次归属。使用最大受表面法自上而下将乔木层划分为第Ⅰ,Ⅱ和Ⅲ亚层,第Ⅰ,Ⅱ亚层属于受光层,第Ⅲ亚层为非受光层。在每块标准地选择对角线上的3块样地进行灌木层与草本层植物调查,记录灌木层乔木与灌木幼树的数量、大小与分布情况,记录草本层中乔木与灌木幼苗及草本植物的数量、大小与分布情况。
2.2数据处理
物种重要值 IV计算公式(宋永昌, 2001)为IV=(Dr+Pr+Fr)/3,Dr,Pr和Fr分别为该物种的相对多度、相对显著度和相对频度,都用百分数表示。所有物种的IV总和为100%。
Shannon-Wiener多样性指数(H′)和均匀度(E)计算公式(Magurran, 2004)为:

式中:S为物种丰富度即总物种数,pi为物种i的多度占的比例,pi=ni/N,N为各物种个体数之和,ni为物种i的个体数。
两亚层间物种组成的相似性指数CS计算公式(王伯荪等, 1985)为CS=2c/(a+b),c为2个亚层的共有种数,a,b分别为2个亚层的物种数。
重要值、Shannon-Wiener多样性指数、均匀度指数和相似性指数的计算均使用Excel软件完成。
3 结果与分析
3.1群落特征
1~5号标准地都具有高大的群落外貌,最大树高分别为37.0,40.0,35.0,33.5和38.0 m,基于胸高断面积的加权平均树高分别为26.8,27.9,23.9,24.3和25.3 m。各标准地最大林木的胸径都不低于80.0 cm,林分平均胸径分别为21.0,25.7,22.0,23.4和21.9 cm。1~5号标准地胸径≥5.0 cm的林木密度分别为1 164,952,1 076,1 056和1 056株·hm2。根据最大受光面法,5块标准地受光层与非受光层之间的临界高度分别为17,16.5,17,17和16 m,非受光层即第Ⅲ亚层,在受光层中,第Ⅰ,Ⅱ亚层间的临界高度分别为25,27,25,23和25 m。
5块标准地都不存在占绝对优势的树种。1号标准地重要值排在前两位的树种为木荷和光叶山矾,重要值分别为23.5%和9.6%; 2号标准地为猴欢喜和木荷,重要值分别为16.1%和10.5%; 3号标准地为木荷和新木姜子,重要值分别为12.8%和8.7%; 4号标准地为木荷和浙江桂,重要值分别为20.1%和12.6%; 5号标准地为浙江桂和木荷,重要值分别为17.6%和13.4%。
各样地灌木层中以更新幼树为主,真正的灌木有山矾(S.sumuntia)、细枝柃(Euryaloquaiana)、细齿叶柃(E.nitida)、黄瑞木(Adinandramillettii)、狗骨柴(Diplosporadubia)、沿海紫金牛(Ardisiapunctata)、尖叶水丝梨(Sycopsisdunnii)、鼠刺(Iteachinensis)、三花冬青(Ilextriflora)和罗浮冬青(I.lohfauensis)。各样地的草本层不发达,盖度为15%±5%。1号标准地有6种草本植物: 草珊瑚(Sarcandraglabra)、狗脊蕨(Woodwardiajaponica)、单羽耳蕨(Polystichumsimplicipinum)、三叶崖爬藤(Tetrastigmahemsleyanum)、团叶陵齿蕨(Lindsaeaorbiculata)和扇叶铁线蕨(Adiantumflabellulatum)。2~5号标准地草本植物种较少,2号标准地有草珊瑚和狗脊蕨,3号标准地有金毛狗(Cibotiumbarometz)、草珊瑚和狗脊蕨,4和5号标准地仅调查到狗脊蕨。
3.2乔木层及其各亚层的多样性
各标准地乔木层物种丰富度表现为1lt;5lt;4lt;3lt;2,Shannon-Wiener指数表现为1lt;4lt;5lt;2lt;3,均匀度指数表现为1lt;2lt;4lt;5lt;3,1号样地的物种丰富度和多样性水平相对较低,2~5号样地相对较高(表1)。由于位于相同的坡面且空间距离接近,在相同的封禁保护条件下,2和3号样地的物种丰富度相近,4和5号样地不仅物种丰富度差别不大,多样性指数和均匀度指数也非常接近。Shannon-Wiener指数的排序情况与丰富度的排序情况更接近。各亚层物种丰富度的变化规律相同,5块标准地都是越向下物种越多,Shannon-Wiener指数(H′)也是越向下越大(表1)。均匀度(E)没有统一的变化规律性,从最高层向下,1,2和5号标准地E值越来越小; 3,4号标准地先增加再降低(表1)。
3.3各亚层的主要树种及数量特征
各标准地不同亚层的主要树种(重要值排前5位的树种)重要值之和占所在亚层重要值总和的比例都是从第Ⅰ亚层到第Ⅲ亚层递减(表2)。在树种组成上,1号标准地包含一定量的阳性先锋树种——以枫香为代表,他们无法在林下更新,随演替进展,将被群落中的木荷和栲(Castanopsis)等常绿树种所替代; 2~5号标准地各个亚层都是中性和耐荫的常绿树种占优势,群落处于相对稳定状态。各标准地Ⅰ亚层的优势种胸高断面积大于其他亚层或乔木层优势种的胸高断面积,以2号标准地为例: 在乔木层中猴欢喜的重要值最大,但是其植株较小、只分布于Ⅱ和Ⅲ亚层,所有58株猴欢喜的胸高断面积(1.2 m2)尚不及Ⅰ亚层中7株木荷(合计2.4 m2)或4株拉氏栲(C.lamontii,合计2.0 m2)的胸高断面积。
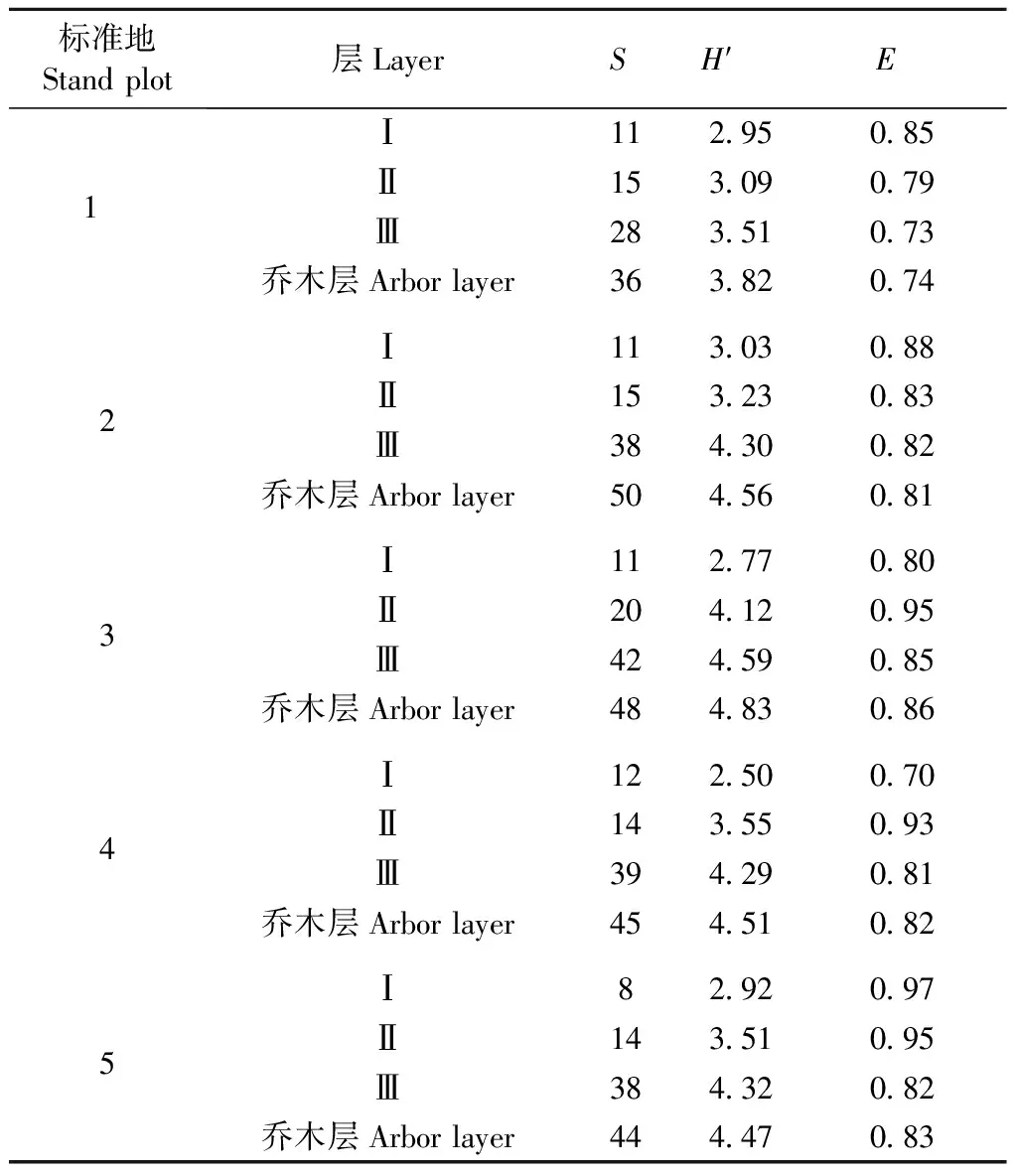
表1 乔木层和各亚层的物种丰富度S、Shannon-Wiener多样性指数H′和均匀度(E)Tab.1 Species abundance (S),biodiversity (H′) andevenness (E) of arbor layer and sub-layer
3.4各亚层树种的生长特性
在5块标准地能生长到第Ⅰ亚层的35个树种中,木荷、浙江桂、米槠(C.carlesii)能生长到30~35 m或更高,直径达60 cm以上,且株数较多,是群落的优势种或者重要的伴生树种。高度能达到30 m以上但数量较少的,有常绿阔叶树种拉氏栲、罗浮栲(C.faberi)、丝栗栲(C.fargesii)、刨花楠(Machiluspauhoi)、香樟(Cinnamomumcamphora)、观光木(Tsoongiodendronodorum)、福建含笑(Micheliafujianensis)和乐东拟单性木兰(Parakmerialotungensis)(它们是群落中的优势树种或伴生树种),还有落叶树种枫香、拟赤杨(Alniphyllumfortunei)、蓝果树(Nyssasinensis)和苦枥木(Fraxinusinsularis)(它们属于阳性先锋树种)。福建山矾(S.fukienensis)、红皮树(Styraxsuberifolius)、山杜英(Elaeocarpussylvestris)、新木姜子、刺叶樱(Prunusspinulosa)、木荚红豆(Ormosiaxylocarpa)和多穗石栎(Lithocarpuspoplystachyus)的高度相对较低,能生长到25~30 m,最大胸径相对较小,最大不超过60 cm,他们是主要伴生树种或偶见种(刺叶樱、木荚红豆和多穗石栎各有1株)。具有潜力但尚未生长到第Ⅰ亚层的还有青冈栎(Cyclobalanopsisglauca)、闽楠(Phoebebournei)、沉水樟(C.micranthum)、苦槠(C.sclerophylla)、美秀栲(C.jucunda)、南岭锥(C.fordii)、厚壳桂(Cryptocaryachinensis)和格木(Erythrophleumfordii),除闽楠、沉水樟、厚壳桂有2株外,其余均仅有1株。
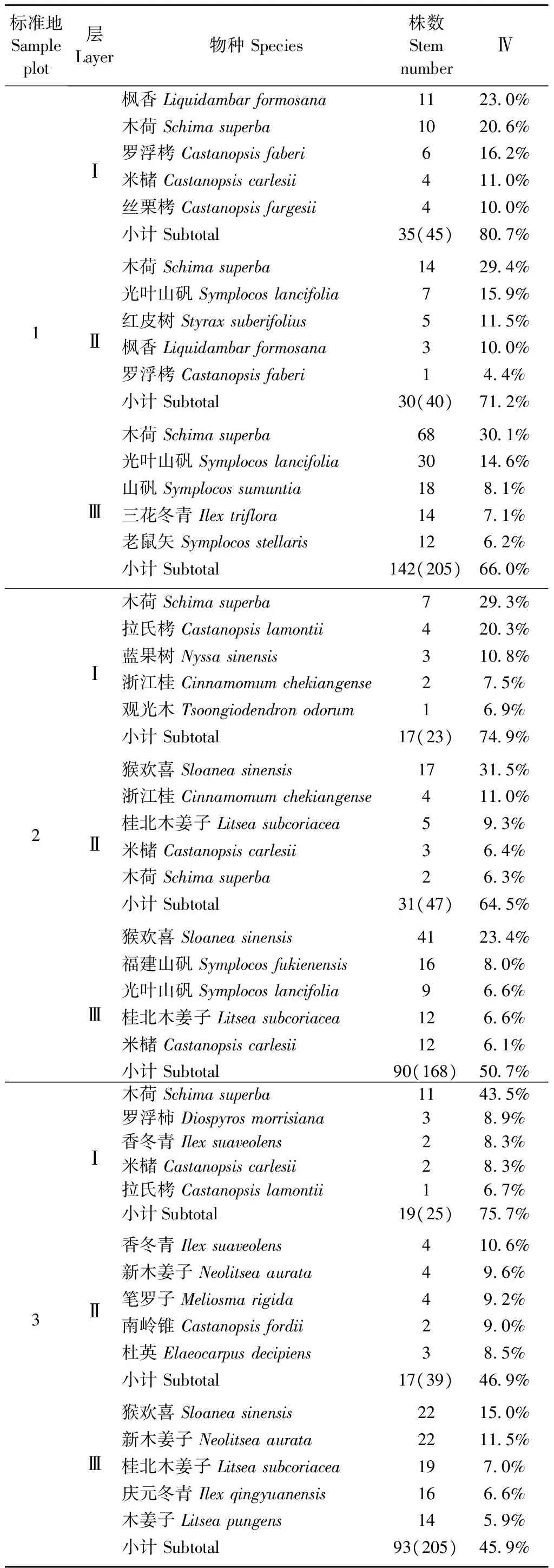
表2 各亚层内重要值排前5位的树种及其数量特征①Tab. 2 IV of top five tree species in each sub-layer andthere quantitative feature
①括号中数字为亚层内所有林木的总株数 Numbers in brackets are total stems in each sub-layer.
每块标准地约有10种分布在第Ⅰ亚层,多数在3个亚层中间断分布,只有个别最高层树种能在3个乔木亚层连续分布: 1号标准地有福建山矾、罗浮栲和木荷,2号标准地有红皮树、木荷和浙江桂,3号标准地有米槠、木荷、香冬青、新木姜子和笔罗子,4号标准地有木荷、木姜子和浙江桂,5号标准地有浙江桂。红皮树、香冬青、木姜子、新木姜子和笔罗子在各标准地第Ⅰ亚层均为1株。
能生长到第Ⅱ亚层但不能达到第Ⅰ亚层的有20个种(含4种灌木),其中光叶山矾、桂北木姜子、猴欢喜、山黄皮、杜英、庆元冬青和三花冬青,最大高度25 m,最大胸径20~40 cm,他们的株数较多,是重要的伴生树种。华杜英(E.chinensis)、薯豆(E.japonicus)、延平柿(Diospyrostsangii)和毡毛泡花树(Meliosmarigidavar.pannosa)同样可以生长到20~25 m,但株数较少。灌木树种山矾、黄瑞木、弯蒴杜鹃(Rhododendronhenryi)、尖叶水丝梨也可以生长到第Ⅱ亚层,但最大胸径普遍较小,只有1.6%的个体胸径超过20 cm。
只能生长在第Ⅲ亚层中的乔木树种(不包括灌木)有17种,最大树高约16 m,最大胸径很少超过20 cm,大部分属于常绿阔叶树种,如野含笑(M.skinneriana)、水丝梨(S.sinensis)、椤木石楠(Photiniadavidsoniae)、虎皮楠(Daphniphyllumoldhami)、厚皮香(Ternstroemiagymnanthera)和树参(Dendropanaxdentiger)等,只有合欢(Albiziajulibrissin)、野柿(D.kakivar.silvestris)、南岭黄檀(Dalbergiabalansae)属于落叶阔叶树种。在5块标准地的下林冠层还分布有灌木树种鼠刺、檵木(Loropetalumchinense)、台湾冬青(I.formosana)、连蕊茶(Camelliafraterna)、细枝柃、细齿叶柃和赤楠(Syzygiumbuxifolium)等。大多数灌木树种较同层乔木树种细弱,最大胸径不超过15 cm,前述个别能生长到第Ⅱ亚层的除外。
3.5各亚层的相似性分析
在1~5号样地,越低的亚层与乔木层整体的相似性越大(表3)。大多数灌木树种只能分布在第Ⅲ亚层和灌木层即更新层,个别能分布在第Ⅱ亚层,因此越低的乔木亚层与灌木层的相似性越大,但是低于相应亚层与整个乔木层的相似性。
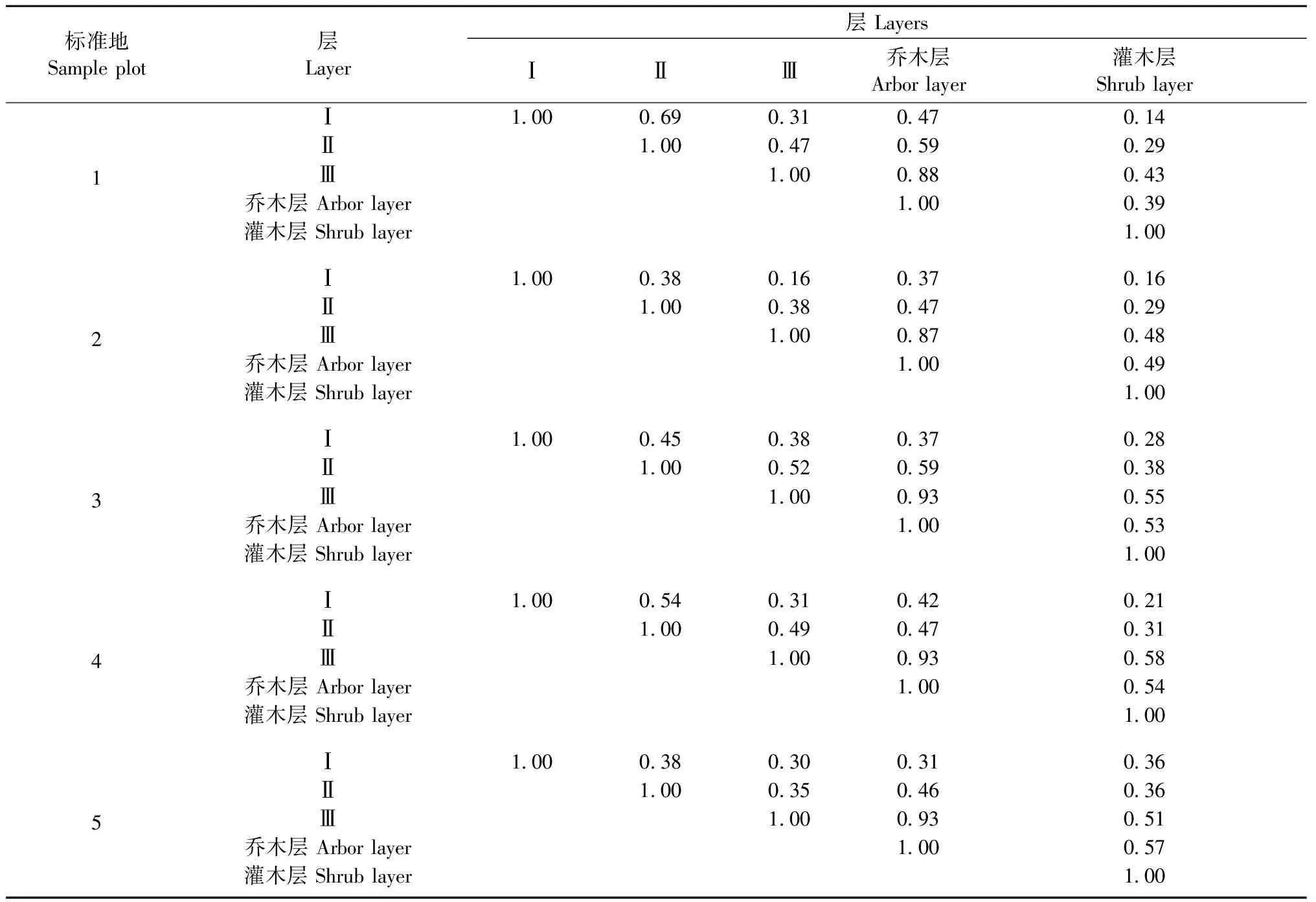
表3 层间的相似性Tab. 3 Coefficient of similarity between different layers
第Ⅰ亚层与其他2个亚层物种组成的相似性有统一的规律(表3),均表现为第Ⅰ和Ⅱ亚层的相似性大于第Ⅰ和Ⅲ亚层的相似性,也大于第Ⅰ亚层和整个乔木层的相似性。两两亚层之间的相似性没有统一的规律,1,4和5号标准地,都是第Ⅰ和Ⅱ亚层的相似性最大; 2号标准地第Ⅰ和Ⅱ亚层与第Ⅱ和Ⅲ亚层的相似性系数相等且较大,第Ⅰ亚层与第Ⅲ亚层的相似性较低; 3号标准地第Ⅱ和Ⅲ亚层的相似性最大。
4 讨论
本研究中,各标准地第Ⅰ和Ⅱ亚层树种组成与整个乔木层的树种组成差异都较大,第Ⅲ亚层与乔木层的差异较小。由于各亚层代表着不同高度的空间位置,因此推断在第Ⅲ亚层林木向第Ⅱ亚层“冲刺”阶段,空间生态位分化的效应最显著。而第Ⅱ与第Ⅲ亚层之间的分界面正是受光层与非受光层的分界面,即最大受光面,这可能是最大受光面(庄崇洋等, 2017)的生态学意义所在。典型林分受光层与非受光层的分化现象,对区域内的目标树经营实践具有潜在的指导意义——发生分化的时期可能正是选择目标树的时机,在分化前、后选择目标树可能是不恰当的,在分化前目标树可能因较大的竞争压力而被自然稀释,在分化后目标树可能已经定型,留给经营调整的空间过低。
在多样性仅次于热带雨林的亚热带常绿阔叶林(贺金生等, 1998),存在多样性维持机制的多种解释(Shenetal., 2009; Legendreetal., 2009; Zhuetal., 2010; Bothetal., 2011; Bruelheideetal., 2011; Luoetal., 2011; Langetal., 2012; Wangetal., 2016),主要包括随机因素、空间异质性和生态位分化。本研究中,各样地亚层之间组成种的相似性结果(表3)以及只有个别树种能在3个亚层连续分布的现象,说明存在树种的随机迁移与替代过程(Lavoredetal., 2002; Hérault, 2007; Bruelheideetal., 2011)。在光资源竞争中受光层林木占有明显的优势地位却又能与非受光层林木共存不同垂直高度分布各不相同的(或者说最大高度不同的)树种的进化史、生物学特色及种群生态学特征(Kohyamaetal., 2009; 2012; Onodaetal., 2014)各不相同,今后需进一步探讨。
总之,物种丰富、结构复杂的森林群落具有复杂的生物多样性形成和维持机制,可能存在多种共同起作用的驱动力,需要深入开展综合性研究。
5 结论
按最大受光面法,调查的典型中亚热带天然阔叶林群落的乔木层都可以分成3个亚层,代表着不同林木在光竞争中的地位,具有生物学意义。以整个乔木层为参照、以相似性代表层与层间的分化程度时,受光层与非受光层之间分化最显著,而受光层与非受光层之间的分界面正是最大受光面。
1号木荷-光叶山矾群落的组成种和多样性水平最低,丰富度(S)为36,Shannon-Wiener多样性指数(H′)为3.82。2号猴欢喜-木荷群落和3号木荷-新木姜子群落的丰富度和多样性水平较接近,S值分别为50和48,H′分别为4.56和4.83。4号木荷-浙江桂群落和5号浙江桂-木荷群落的丰富度和多样性水平接近,S值分别为45和44,H′分别为4.51和4.47。各标准地中,越低的亚层树种丰富度和多样性水平越高,相同亚层的丰富度和多样性水平接近,第Ⅰ亚层S值为8~11,H′为2.5~3.03; 第Ⅱ亚层S值为15~20,H′为3.09~4.12; 第Ⅲ亚层S值为28~42,H′为3.51~4.59。
木荷、浙江桂、米槠、栲类、观光木、福建含笑和乐东拟单性木兰等能生长到第Ⅰ亚层的树种在光资源竞争中占据绝对优势,最大树高可达(30±5)m甚至40 m,最大胸径40~60 cm,部分树种可超过60 cm。光叶山矾、桂北木姜子、猴欢喜等只能生长到第Ⅱ亚层的林木,最大高度为(25±2)m,最大胸径20~40 cm,它们是重要的伴生树种。灌木树种山矾、黄瑞木、尖叶水丝梨和弯蒴杜鹃的高度也可达到第Ⅱ亚层,但胸径基本不超过20 cm。野含笑、虎皮楠、厚皮香、树参、合欢、野柿和南岭黄檀等只能在第Ⅲ亚层分布的林木,最大树高不超过16或17 m,最大胸径很少超过20 cm。第Ⅲ亚层还分布有大量灌木,但是更为细弱,最大胸径一般不超过15 cm。林木的垂直分布特点和受光层与非受光层的分化现象对近自然经营活动具有潜在的指导意义,对目标树的选择及选择时机有指导意义。
陈灵芝,陈清朗,刘文华. 1997. 中国森林多样性及其地理分布.北京: 科学出版社.
(Chen L Z,Chen Q L,Liu W H.1997. Forest diversity and its geographical distribution in China. Beijing: Science Press. [in Chinese])
樊后保.2000. 格氏栲群落的结构特征. 林业科学, 36(2): 6-12.
(Fan H B. 2000. Structure features ofCastanopsiskawakamiicommunity. Scientia Silvae Sinica, 36(2): 6-12. [in Chinese])
贺金生, 陈伟烈, 李凌浩. 1998. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征. 植物生态学报, 22(4): 303-311.
(He J S, Chen W L, Li L H. 1998. Community diversity of the main types of the evergreen broad-leaved forest in the eastern part of the middle subtropical China. Acta Phytoecologica Sinica, 22(4): 303-311. [in Chinese])
何友钊. 1990. 建瓯县万木林保护区史事考∥中国林学会林业史学会. 林史文集: 第一集. 北京: 中国林业出版社.
(He Y Z. 1990. Examining for the historical events in Wanmulin Nature Reserve∥Society of Forestry Historiography of Chinese Society of Forestry. Corpus of forestry historiography I. Beijing: China Forestry Publishing House.[in Chinese])
黄清麟. 1998. 中亚热带天然阔叶林可持续经营技术研究. 北京: 北京林业大学博士学位论文.
(Huang Q L. 1998. Study on sustainable management technology of natural broad-leaved forest of mid-subtropical zone. Beijing:PhD thesis of Beijing Forestry University. [in Chinese])
黄清麟, 董乃钧, 李元红. 1999. 福建中亚热带天然阔叶林的类型与特征.山地学报, 17(4): 368-374.
(Huang Q L, Dong N J, Li Y H. 1999. Main types and characteristics of natural broad-leaved forest of the mid-subtropical zone in Fujian Province. Journal of Mountain Science, 17(4): 368-374. [in Chinese])
黄清麟, 李志明, 郑群瑞. 2003. 福建中亚热带天然阔叶林理想结构探讨. 山地学报, 21(1): 116-120.
(Huang Q L, Li Z M, Zheng Q R.2003. A discussion on the ideal structure of natural broad-leaved forest in the mid-subtropical zone of Fujian Province. Journal of Mountain Science, 21(1): 116-120. [in Chinese])
黄清麟, 罗发潘, 郑群瑞. 1995. 闽北天然阔叶林林层特征研究. 福建林学院学报, 15(1): 17-21.
(Huang Q L, Luo F P, Zheng Q R. 1995. Vertical structure of natural broad-leaved forest in North Fujian. Journal of Fujian College of Forestry, 15(1): 17-21. [in Chinese])
金 钊, 杨玉盛, 董云社, 等. 2007. 福建万木林自然保护区米槠和杉木细根分解动态. 地理研究, 26(3): 491-500.
(Jin Z, Yang Y S, Dong Y S,etal. 2007. Studies on decomposition of fine roots ofCastanopsiscarlesiiandCunninghamialanceolata(chinese fir) in Wanmulin Natural Reserve, Fujian Province. Geographical Research, 26(3): 491-500.[in Chinese])
刘茂松, 冯 霞, 姜志林, 等. 1998. 万木林主要森林群落特征及演替动态. 南京林业大学学报:自然科学版, 22(2): 5-10.
(Liu M S, Feng X, Jiang Z L,etal. 1998. The community features and succession of the flora in Wanmulin Natural Reserve. Journal of Nanjing Forest University:Natural Sciences Edition, 22(2): 5-10.[in Chinese])
裴广廷, 马红亮, 林 伟, 等. 2015. 氨基酸添加对亚热带森林红壤氮素转化的影响. 生态学报, 35(23): 7774-7784.
(Pei G T, Ma H L, Lin W,etal. 2015. Effects of amino acid additions on nitrogen transformation in subtropical forest soil. Acta Ecologica Sinica, 35(23): 7774-7784. [in Chinese])
宋永昌.2001. 植被生态学. 上海: 华东师范大学出版社.
(Song Y C. 2001. Vegetation ecology. Shanghai: East China Normal University Press.[in Chinese])
宋永昌. 2004. 中国常绿阔叶林分类试行方案. 植物生态学报, 28(4): 435-448.
(Song Y C. 2004. Tentative classification scheme of evergreen broad-leaved forests of China. Acata Phytoecologica Sinica, 28(4): 435-448. [in Chinese])
宋永昌, 阎恩荣, 宋 坤. 2015. 中国常绿阔叶林8大动态监测样地植被的综合比较. 生物多样性, 23(2): 139-148.
(Song Y C, Yan E R, Song K.2015. Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, China. Biodiversity Science, 23(2): 139-148.[in Chinese])
王伯荪, 彭少麟. 1985. 鼎湖山森林群落分析. IV.相似性和聚类分析. 中山大学学报:自然科学版, (1): 31-38.
(Wang B S, Peng S L.1985. Analysis on the forest communities of Dinghushan, Guangdong. IV. Similarity and cluster analysis.Acta Scientiarum Naturalium Universitatis Sunyatseni, (1): 31-38. [in Chinese])
王梅峒. 1988. 江西亚热带常绿阔叶林的生态学特征. 生态学报, 8(3): 249-255.
(Wang M T. 1988. The ecological features of evergreen broad-leaved forest of subtropical in Jiangxi Province. Acta Ecologica Sinica, 8(3): 249-255. [in Chinese])
吴承祯, 洪 伟, 陈 辉, 等. 1996. 万木林中亚热带常绿阔叶林物种多样性研究. 福建林学院学报, 16(1): 33-37.
(Wu C Z, Hong W, Chen H,etal. 1996. Species diversity of subtropical evergreen broad-leaved forest in Wanmulin mountain. Journal of Fujian College of Forestry, 16(1):33-37. [in Chinese])
叶万辉, 曹洪麟, 黄忠良, 等. 2008. 鼎湖山南亚热带常绿阔叶林内20公顷样地群落特征研究. 植物生态学报, 32(2): 274-286.
(Ye W H, Cao H L, Huang Z L,etal. 2008. Community of structure of a 20 hm2lower subtropical evergreen broadleaved forest plot in Dinghushan, China. Journal of Plant Ecology, 32(2): 274-286. [in Chinese])
蚁伟民, 曹洪麟, 王伯荪, 等. 1999. 鼎湖山格木群落的组成种类和结构特征. 热带亚热带植物学报, 7(1): 7-14.
(Yi W M, Cao H L, Wang B S,etal. 1999. Species composition and structural feature inErythrophleumfordiicommunity in Dinghushan. Journal of Tropical and Subtropical Botany, 7(1): 7-14. [in Chinese])
赵丽娟, 项文化, 李家湘, 等. 2013. 中亚热带石栎-青冈群落物种组成、结构及区系特征. 林业科学, 49(12): 10-17.
(Zhao L J, Xiang W H, Li J X,etal. 2013. Floristic Composition, structure and phytogeographic characteristics in aLithocarpusglaber-Cyclobalanopsisglaucaforest community in the subtropical region. Scientia Silvae Sinicae, 49(12): 10-17. [in Chinese])
郑群瑞. 1999. 万木林自然保护区珍稀特有野生植物. 福建林学院学报, 19(1): 70-73.
(Zheng Q R. 1999. The investigation on rare and peculiar wild plants in Wanmulin Nature Reserve. Journal of Fujian College of Forestry, 19(1): 70-73.[in Chinese])
中国植被编委会. 1980. 中国植被. 北京: 科学出版社.
(Vegetation of China Editorial Committee. 1980. Vegetation of China. Beijing: Science Press.[in Chinese])
朱锦懋, 姜志林, 郑群瑞, 等. 1997. 福建万木林自然保护区森林群落物种多样性. 南京林业大学学报, 21(4): 11-16.
(Zhu J M, Jiang Z L, Zheng Q R,etal. 1997. A study on the species diversity in the forest community of Wanmulin Natural Reserve Fujian Province. Journal of Nanjing Forestry University, 21(4): 11-16. [in Chinese])
祝 燕, 赵谷风, 张俪文, 等. 2008. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构. 植物生态学报, 32(2): 262-273.
(Zhu Y, Zhao G F, Zhang L W,etal. 2008. Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, East China. Journal of Plant Ecology, 32(2): 262-273. [in Chinese])
庄崇洋, 黄清麟, 马志波, 等. 2017. 中亚热带天然阔叶林林层划分新方法——最大受光面法. 林业科学, 53(3): 1-11.
(Zhuang C Y, Huang Q L, Ma Z B,etal. 2017. A new method of storey identification of natural broad-leaved forests in mid-subtropical zone—maximum light receiving plane. Scientia Silvae Sinicae, 53(3): 1-11. [in Chinese ])
Aschehoug E T, Brooker R, Atwater D Z,etal. 2016. The Mechanisms and consequences of interspecific competition among plants. Annual Review of Ecology, Evolution, and Systematics, 47: 263-281.
Both S, Fang T, Böhnke M,etal. 2011. Lack of tree layer control on herb layer characteristics in a subtropical forest, China. Journal of Vegetation Science, 22(6): 1120-1131.
Bruelheide H, Böhnke M, Both S,etal. 2011. Community assembly during secondary forest succession in a Chinese subtropical forest. Ecological Monographs, 81(1): 25-41.
Chen S Y, Zhang J L, Jia P,etal. 2010. Effects of size variation and spatial structure on plastic response of plant height to light competition. Chinese Science Bulletin:Ecology, 55(12), 1135-1141.
Falster D S, Westoby M. 2003. Plant height and evolutionary games. Trends in Ecology and Evolution, 18(7), 337-343.
Hérault B. 2007. Reconciling niche and neutrality through the emergent group approach. Perspectives in Plant Ecology Evolution amp; Systematics, 9(2): 71-78.
Kohyama T, Takada T. 2009. The stratification theory for plant coexistence promoted by one-sided competition. Journal of Ecology, 97(3):463-471.
Kohyama T S, Takada T. 2012. One-sided competition for light promotes coexistence of forest trees that share the same adult height. Journal of Ecology, 100(6): 1501-1511.
Lang A C, Härdtle W, Baruffol M,etal. 2012. Mechanisms promoting tree species co-existence, experimental evidence with saplings of subtropical forest ecosystems of China. Journal of Vegetation Science, 23(5): 837-846.
Lavorel S, Garnier E. 2002. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the holy grail. Functional Ecology, 16(5): 545-556.
Legendre P, Mi X, Ren H,etal. 2009. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology, 90(3): 663-674.
Luo Z, Mi X, Chen X,etal. 2012. Density dependence is not very prevalent in a heterogeneous subtropical forest. Oikos, 121(8): 1239-1250.
Magurran A E. 2004. Measuring biological diversity. Oxford: Blackwell Science.
Shen G, Yu M, Hu X S,etal. 2009. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity. Ecology, 90(11): 3033-3041.
Wang X, Wiegand T, Kraft N J B,etal. 2016. Stochastic dilution effects weaken deterministic effects of niche-based processes in species rich forests. Ecology, 97(2): 347-360.
Zhu Y, Mi X C, Ren H B. 2010. Density dependence is prevalent in a heterogeneous subtropical forest. Oikos, 119(1): 109-119.
(责任编辑 于静娴)
SpeciesCompositionandDiversityofTypicalNaturalBroad-LeavedForestinCentralSubtropical:BasedonArborLayerStratification
Ma Zhibo1Huang Qinglin1Zhuang Chongyang1Zheng Qunrui2Wang Hong1
(1.KeyLaboratoryofForestryRemoteSensingandInformationTechnology,StateForestryAdministrationResearchInstituteofForestResourceInformationTechniques,ChineseAcademyofForestryBeijing100091; 2.AdministrativeOfficeofWanmulinProvincialNatureReserveJian’ou353105)
【Objective】The study was to determine the species composition and diversity of different forest layers and to determine how vertical distribution of trees affect the community structure and its underlying mechanisms with the view of arbor layer’s stratification.【Method】Five 50 m×50 m sample plots were investigated in Wanmulin Provincial Nature Reserve, Fujian Province. With maximum light receiving plane (MLRP) method, arbor layer of each plots were divided into three sub-layers from top to bottom, the sub-layer I, II and III . The sub-layer I and II belonged to light receiving layer (LRL), the sub-layer III was non-light receiving layer (NLRL). Then the species composition and diversity of each sub-layer were studied using importance value, diversity index and similarity index. 【Result】The species richness and diversity increased when the arbor layer decreased from sub-layer I to sub-layer III of the five plots. The composition and number of species varied in the same sub-layers of different plots. The maximum tree height of 35 tree species can reach about 30 m even 40 m and they have the potential to distribute at top layer of the community, i.e. sub-layer I, but there were only 10 tree species could successive distribute at all the three sub-layers among all species of all plots. The maximum height of 16 tree species and 4 shrub species was about 25 m and sub-layer II was the limit of what they could reach. The maximum height of 17 tree species and 13 shrub species was 16-17 m and sub-layer III was the limit of what they could reach. Tree species in LRL were dominant species and important associated species of arbor layer, including evergreen treesSchimasuperba,Cinnamomumchekiangense,C.camphora,Machiluspauhoi,Tsoongiodendronodorum,Micheliafujianensis,Parakmerialotungensisand some trees species ofCastanopsis, andLiquidambarformosana,Alniphyllumfortunei,NyssasinensisandFraxinusinsularis. Tree species only distributed in NLRL included evergreen treesM.skinneriana,Sycopsissinensis,Photiniadavidsoniae,Daphniphyllumoldhami,Ternstroemiagymnanthera, and deciduous treesAlbiziajulibrissin,Diospyroskakivar.silvestrisandDalbergiabalansae. Most shrubs species could only distribute in NLRL exceptSymplocossumuntia,Adinandramillettii,RhododendronhenryiandS.dunniicould reach to sub-layer. Priority should be given to dominant species of sub-layer I when determining dominant species of the community, because their basal areas were obviously bigger than trees of other sub-layers. The similarity of sub-layer I and II was bigger than that of sub-layer I and III or sub-layer I and arbor layer. The differentiation was prominent at the critical plane LRL and NLRL when the similarity was the degree of differentiation and the arbor layer was the reference, suggesting spatial niche separation being prominent at MLRP. 【Conclusion】In the five sample plots, dominant species and important associated tree species of arbor layer were mainly distributed in the LRL. Niche differentiation was prominent at MLRP. There were obvious difference between LRL and NLRL in species richness and diversity, which were identified according to MLRP. The differentiation of the LRL and the NLRL has potential practical significance for the selection of target trees in close-to-nature management so that the relationship between the two should be further studied in the future. Very few tree species could distribute in 3 successive sub-layers of each plots among all tree species that could grow to sub-layer I, suggesting that there were almost no particular species could be permanent dominant species in the LRL and species composition and richness in the 5 plots appeared to be caused also by random species replacement, which needs to be further studied in future.
natural evergreen broad-leaved forest; mid-subtropical zone; arbor layer; stratification; similarity
10.11707/j.1001-7488.20171002
2017-03-27;
2017-09-13。
国家自然基金科学项目“中亚热带天然阔叶林林层特征研究”(31370633)。
*黄清麟为通讯作者。
S718.5
A
1001-7488(2017)10-0013-09
猜你喜欢
中国林副特产(2022年4期)2022-08-08 05:50:30
建筑与预算(2022年6期)2022-07-08 13:59:18
散文诗世界(2022年5期)2022-05-23 02:02:55
绿色中国(2019年18期)2020-01-04 01:57:08
防护林科技(2019年12期)2019-04-14 04:01:18
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
花卉(2016年14期)2016-07-31 18:46:24
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:55
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
