
利用分子标记分析22种水稻10个抗稻瘟病基因的基因型
2017-11-27 17:12贺叶晨星雷子伊李建粤
上海师范大学学报·自然科学版 2017年5期
韩 笑, 贺叶晨星, 张 梅, 雷子伊, 张 涛, 李建粤
(上海师范大学 生命与环境科学学院 植物种质资源开发协同创新中心,上海 200234)
利用分子标记分析22种水稻10个抗稻瘟病基因的基因型
韩 笑, 贺叶晨星†, 张 梅†, 雷子伊†, 张 涛, 李建粤*
(上海师范大学 生命与环境科学学院 植物种质资源开发协同创新中心,上海 200234)
为了明确已克隆的部分稻瘟病抗性基因在上海市主栽水稻品种以及新培育的水稻品系中的分布,选取22份水稻为材料,利用分子标记检测技术,对10个已被克隆的抗稻瘟病基因进行基因型鉴定及分析.结果表明,22种水稻均含Pi36、Pi37、Pi-d2、Pib抗性基因,而Pi-kh、Pi9、Pi2、Pi-ta、Pikm抗性基因在22种水稻中分布各有不同,所有被测水稻中均不携带Pi25抗性基因.
水稻; 分子标记; 稻瘟病抗性基因
稻瘟病是水稻重要的灾害之一,严重时使水稻减产40%~50%以上,甚至绝收.实践证明,利用水稻自身携带的抗病基因是控制稻瘟病害最经济最有效的方法,可以有效地保证粮食安全,减少化学药剂使用量,降低生产成本和保护环境[1].由于稻瘟病菌变异较快[2],一个水稻抗性品种连续多年种植后会使病原菌产生优势小种或新的变异.防治稻瘟病不仅需要发掘抗性持久、抗谱广的基因作为抗源,选育出具有主效抗病基因水稻新品种,同时也可以通过多基因聚合的方式进一步提高水稻对不同稻瘟病生理小种的抵抗效果.
目前大约有100个稻瘟病抗性基因被定位到除 3号染色体外的其余 11条染色体上,其中27个稻瘟病抗性基因已被克隆[3].随着分子生物学和生物技术的迅速发展,不少稻瘟病抗性基因相关的分子标记相继被开发出来,并被用在含有多个抗性基因聚合的研究中.Hittalmani等[4]利用分子标记辅助选择(MAS)技术将抗稻瘟病基因Pi1、Piz5和Pita聚合到水稻品系“BL125中”;Narayanan等[5]通过MAS技术将抗稻瘟病基因Pi1、Pi5和抗白叶枯病基因Xa21在水稻中进行聚合;陈红旗等[6]将3个抗稻瘟病基因聚合到“金23B”水稻;倪大虎等[7]、陈健民等[8]分别将抗稻瘟病基因Pi9 导入不同水稻恢复系中.这些育成的改良品系经抗性鉴定,抗瘟性均得到显著提高.越来越多的抗病基因被挖掘、定位和克隆为水稻抗稻瘟病育种提供了丰富的抗源,加上分子标记辅助选择技术的发展,培育抗病水稻品种的周期缩短,效率提高.
明确上海市水稻品种中究竟具备了哪些抗稻瘟病基因,可为今后利用分子标记辅助在上海地区培育抗稻瘟病水稻新品种研究奠定基础.本研究利用现有报道[9-14]已建立的分子标记,对22种目前上海市主栽的常规水稻和杂交水稻亲本以及部分新培育的水稻新品系进行10个与稻瘟病抗性相关基因的基因型分析.研究结果可为今后利用分子标记辅助培育抗稻瘟病水稻新品种提供参考.
1 材料与方法
1.1水稻材料及基因组DNA提取
22种上海市主栽常规水稻、杂交稻亲本水稻品种或新培育的水稻品系分别为:“嘉花1号”、“秀水123”、“秀水134”、“光明粳3号”(即“gm01”)、“宝农34”、“金丰”、“松早香1号”、“松香早粳”、“松香软粳”、“香软早粳”、“青香软粳”、“嘉晚123”、“武运5103”、“8004”、“29185”、“光明粳2号”(即“银香28”)、“上师大18号”、“上师大19号”、“香宝农34”、“R44”、“繁14”、“上师大5号”.
所有水稻材料取幼嫩叶片进行基因组DNA的提取.
1.222种水稻稻瘟病抗性基因型分析
参考前人报道,分别合成能够扩增Pi-kh、Pi9、Pi-ta、Pi2、Pi25、Pikm、Pi36、Pi37、Pi-d2、Pib共10个基因的引物序列(表1).引物均由上海捷瑞生物工程有限公司合成.
PCR反应为25 μL 体系,含2.5 μL 10×Buffer(含Mg2+),2.5 μL dNTP,每种引物各0.5 μL (2 mmol·L-1),0.3 μL Taq 酶(5 U·μL-1),1 μL DMSO,2.0 μL 模板DNA,用 ddH2O 补足至25 μL体系.不同基因聚合酶链式反应(PCR)扩增的程序,除了退火温度和延伸时间有差异(表1),其他PCR参数均为:94 ℃预变性5 min;94 ℃变性30 s,退火时间30 s,延伸温度72 ℃,共 35 个循环;最后72 ℃延伸10 min,于4 ℃放置3 min.Pi25基因PCR 产物在电泳检测前需要先经HincⅡ内切酶酶切.酶切体系(15 μL):PCR产物8.0 μL,内切酶0.4 μL(10 U·μL-1),10 × Buffer 1.5 μL,ddH2O 5.1 μL,混匀后适宜温度酶切2 h 左右.
反应产物经不同浓度的琼脂糖凝胶进行电泳检测(表1),溴化乙啶染色,在紫外凝胶成像仪上观察.
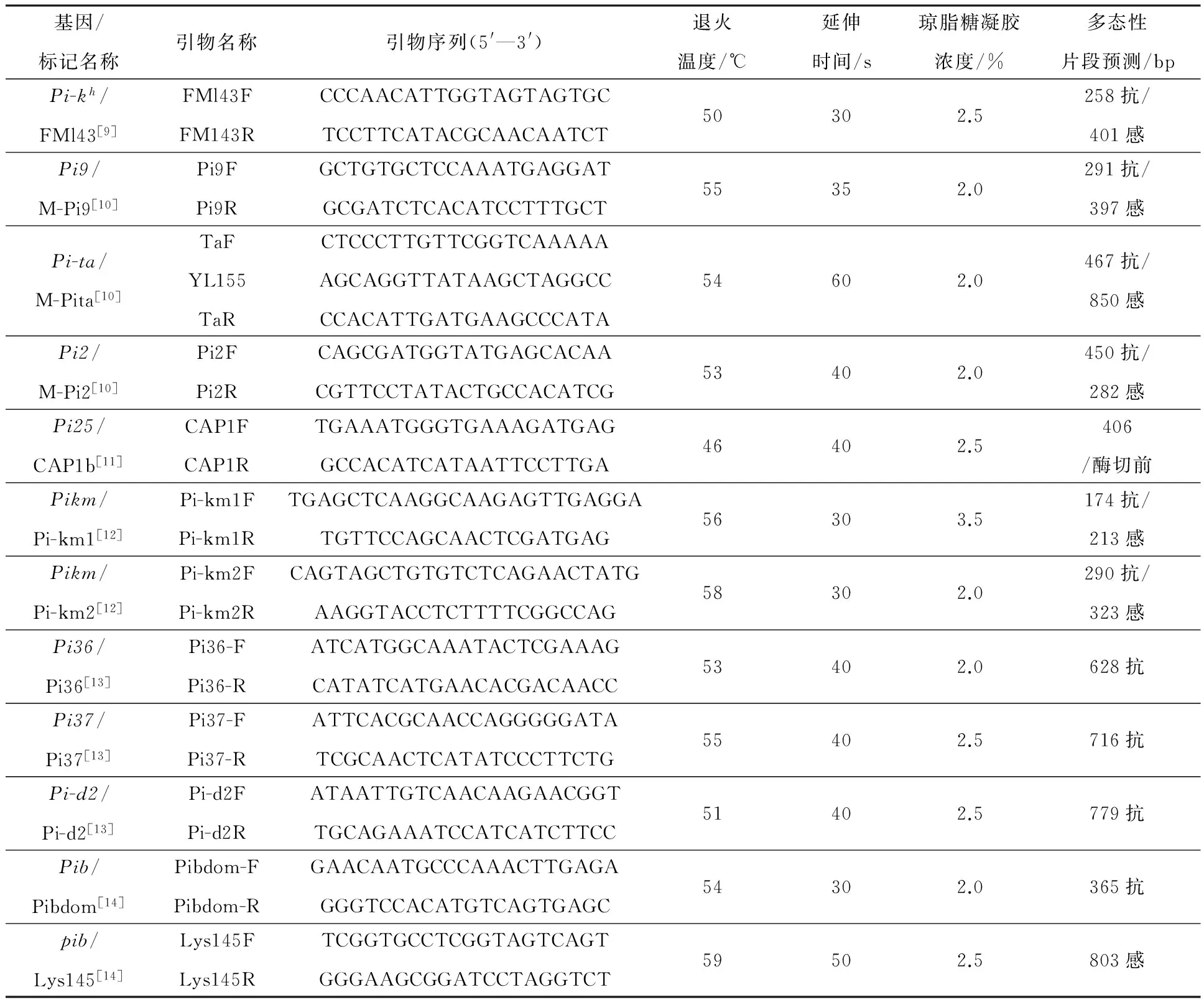
表1 稻瘟病相关基因基因型鉴定涉及的引物序列及相关信息
2 结果与分析
2.1Pi-kh基因型分析
稻瘟病抗病基因Pi-kh位于第11染色体,是Pi-k基因座上5个等位基因中的一个,是稻瘟病广谱抗性基因,对许多中国稻瘟病生理小种有较强的、稳定的抗性[15].与抗病基因相比,感病基因存在143个脱氧核苷酸插入、37个脱氧核苷酸缺失和38个单核苷酸多态性(SNP)[9].利用王军等[9]根据感病基因序列存在143 bp插入的特征设计的InDel标记FM143可检测Pi-kh基因.检测结果显示,“光明粳3号”、“金丰”、“松早香1号”、“松香软粳”、“嘉晚123”、“29185”、“光明粳2号”、“上师大18号”、“繁14”、“上师大5号”水稻能扩增出258 bp的片段,属于含有Pi-kh抗病基因的品种或品系,其余12种水稻都只能扩增出401 bp片段,判断其为不含有Pi-kh抗病基因水稻品种或品系,如图1(a)所示.
2.2Pi9基因型分析
Pi9是目前公认的优良抗稻瘟病基因,该基因位于第6染色体近着丝粒部位,含有6个串联排列的 NBS-LRR,至少对21 个稻瘟病菌小种具有抗性,是一个稻瘟病广谱抗性基因[10].利用高利军等[10]设计的分子标记M-Pi9检测,如含有291 bp特异条带的水稻对稻瘟病菌表现抗或中抗.22种水稻检测结果显示,“宝农34”、“松早香1号”、“松香早粳”、“松香软粳”、“香软早粳”、“青香软粳”、“嘉晚123”、“武运5103”、“上师大19号”、“上师大5号”能扩增出291 bp片段,如图1(b)所示,属于含有Pi9抗性基因的水稻品种或品系,另外12种水稻扩增片段为397 bp,为不含有Pi9抗性基因.

图1 22种水稻10个稻瘟病基因型检测.(a) Pi-kh基因;(b) Pi9基因;(c) Pi-ta基因;(d) Pi2基因;(e) Pi25基因;(f) Pikm1基因;(g) Pikm2基因;(h) Pi36基因;(i) Pi37基因;(j) Pi-d2基因;(k) Pib基因; m)100 bp DNA marker;1:嘉花1号;2:秀水123;3:秀水134;4:光明粳3号;5:宝农34;6:金丰;7:松早香1号;8:松香早粳;9:松香软粳;10:香软早粳;11:青香软粳;12:嘉晚123;13:武运5103;14:8004;15:29185;16:光明粳2号;17:上师大18号;18:上师大19号;19:香宝农34;20:R44;21:繁14;22:上师大5号
2.3Pi-ta基因型检测
Pi-ta是第2个被克隆的稻瘟病抗性基因,位于第12染色体近着丝粒部位.利用高利军等[10]设计的分子标记M-Pita检测Pi-ta基因,如含有467 bp特异性条带的水稻均表现为抗稻瘟病类型,含850 bp的水稻均表现为感病类型.22种水稻检测结果显示,“嘉花1号”、“秀水123”、“秀水134”、“松早香1号”、“松香早粳”、“松香软粳”、“香软早粳”、“嘉晚123”能扩增出467 bp的片段,是含有Pi-ta抗性基因的水稻品种或品系,如图1(c)所示,其他14种水稻只能扩增出850 bp条带,属于不含有Pi-ta抗性基因的水稻.
2.4Pi2基因型检测
抗稻瘟病基因Pi2也位于第6染色体近着丝粒部位,与Pi9互为等位基因[16].Chen 等[17]研究发现,中国792个稻瘟病菌中,只有7.55%的病菌能侵染携带Pi2 的亲本“C101A51”.利用高利军等[10]设计的分子标记M-Pi2检测Pi2基因,如含有450 bp 特异条带的水稻属于含有Pi2抗病基因的品种或品系.22种水稻检测结果显示,只有“嘉花1号”、“秀水123”、“秀水134”、“繁14” 4种水稻能扩增出450 bp的片段,如图1(d)所示.
2.5Pi25基因型检测
抗稻瘟病基因Pi25也位于第6染色体.利用王惠梅等[11]设计的分子标记CAPS/HincⅡ检测Pi25基因,能特异性识别并酶切抗性等位基因.检测结果显示,22种水稻扩增出的片段大小为406 bp,但均未被酶切.根据结果判断这22个水稻品种或品系均不含Pi25抗性基因,如图1(e)所示.
2.6Pikm基因型检测
抗稻瘟病基因Pikm位于第11染色体长臂近末端区.Pikm由2个紧密连锁的具有独立功能NBS-LRR类基因(Pikm1-TS和Pikm2-TS)组成,Pikm1-TS和Pikm2-TS的编码产物均是NBS-LRR类抗病蛋白.利用范方军等[12]设计的分子标记检测Pikm,对2 个基因分别用Pi-km1 分子标记和Pi-km2分子标进行检测,以2 对引物同时检测到目的片段(Pikm1片段长度174 bp,Pikm2片段长度291 bp)为含有完整Pikm基因.
Pi-km1标记对22种水稻检测结果显示,22种水稻均能扩增出174 bp的片段,如图1(f)所示,但是对于Pi-km-2标记检测22种水稻,只有“秀水123”、“秀水134”、“光明粳3号”、“宝农34”、“金丰”、“松早香1号”、“松香软粳”、“嘉晚123”、“武运5103”、“香宝农34”、“上师大5号”水稻能扩增出290 bp特异的抗性片段,如图1(g)所示.由此表明,在这11种水稻中含有完整的Pikm抗性基因.
2.7Pi36基因型检测
Pi36位于第8染色体.利用宋广树等设计的分子标记Pi-36检测,如能够扩增出628 bp的条带表明含有Pi36抗性基因[13].对22种水稻基因组DNA的检测结果发现,22种水稻均扩增出628 bp片段,均含有抗性基因Pi36(图1H).
2.8Pi37基因型检测
Pi37位于第1染色体.利用宋广树等[13]设计的分子标记Pi-37检测,含有抗性基因Pi37的水稻能扩增出716 bp条带.对22种水稻基因组DNA检测结果发现,22种水稻均扩增出716 bp片段,均含有抗性基因Pi37,如图1(i)所示.
2.9Pi-d2基因型检测
利用宋广树等[13]设计的分子标记Pi-d2检测,含有抗性基因Pi-d2的水稻能扩增出779 bp条带.对22种水稻基因组DNA检测结果发现,22种水稻均扩增出779 bp片段,均含有抗性基因Pi-d2,如图1(j)所示.
2.10Pib基因型检测
Pib基因是第一个被克隆的水稻稻瘟病抗性基因,位于第2染色体上.根据刘洋等[14]建立的检测感病等位基因pib的显性标记Lys145和检测抗病等位基因Pib的显性标记Pibdom 组成一套水稻抗稻瘟病基因Pib显性分子标记.抗病性显性分子标记Pibdom 能特异性扩增出一条365 bp 片段,感病性显性分子标记Lys145 能特异性扩增出一条803 bp 的片段.22种水稻检测结果显示,Pibdom均能被检测出365 bp大小的片段,如图1(k)所示,而Lys145检测结果多次重复均无目标条带形成.由此表明22种水稻材料均含有Pib抗性基因.
2.1110个抗性基因在22种水稻中分布汇总
汇总10个抗性基因在22种水稻中的分布,并计算每个抗性基因在22种水稻中出现的频率.
22种水稻都含有多个稻瘟病抗性基因,但数量不同,其中“松早香1号”和“松香软粳”含有抗性基因多达8个,其次是“秀水123”、“秀水134”、“上师大5号”,都含有7个抗性基因.“8004”和“R44”两种水稻在22种水稻中含有的抗性基因数量最少,都仅有4个(表2).
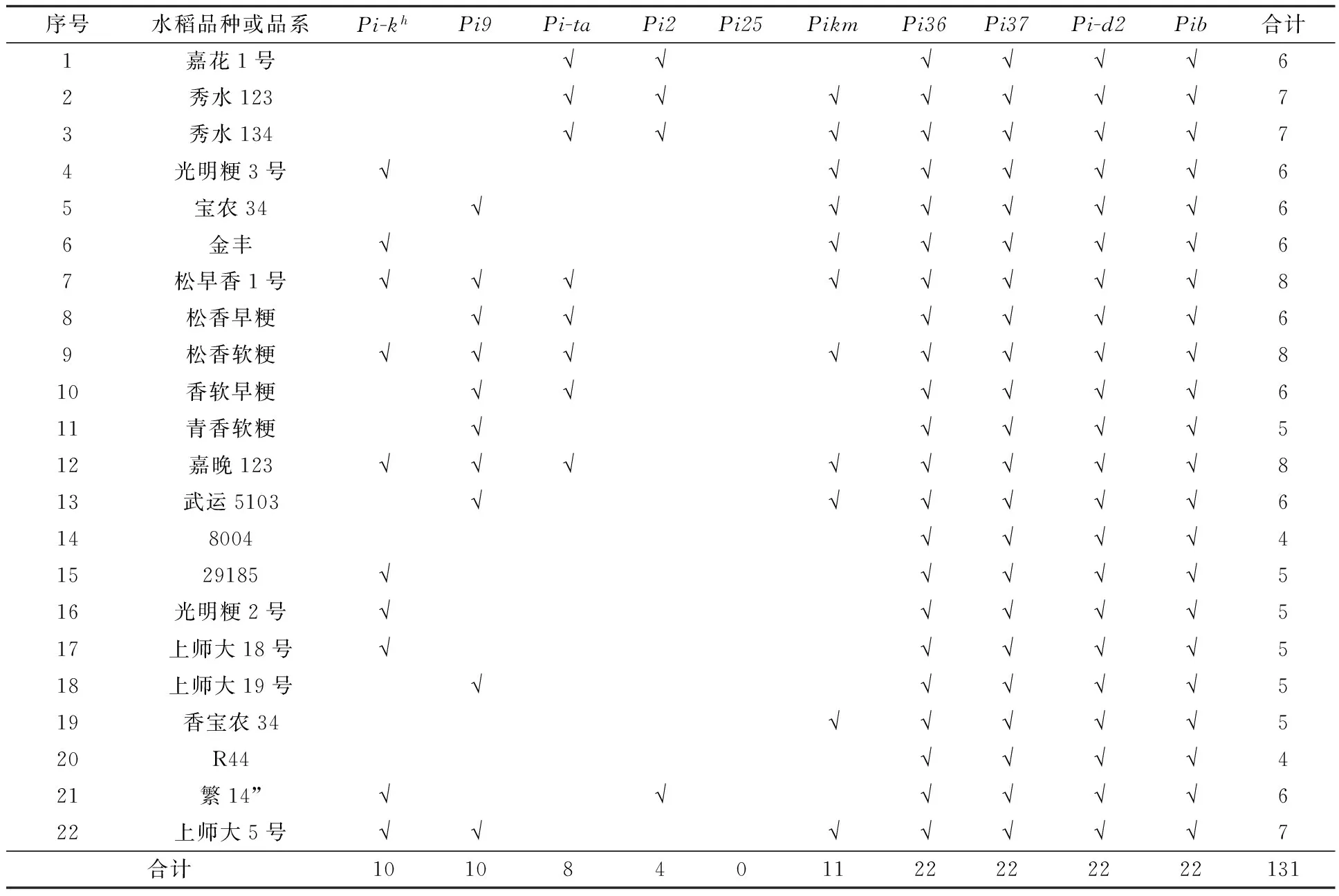
表2 10个稻瘟病抗性基因在22种水稻中出现次数汇总
注:表示含有抗性基因
在上海市目前主裁的常规水稻和杂交稻亲本及最近培育的22种水稻中,对于Pi36、Pi37、Pi-d2、Pib4个基因出现的频率较高,都到达100%出现次数.Pi25基因出现次数为0,频率最低.其次是Pi2基因,只出现了4次,出现频率为18.2%(图2).
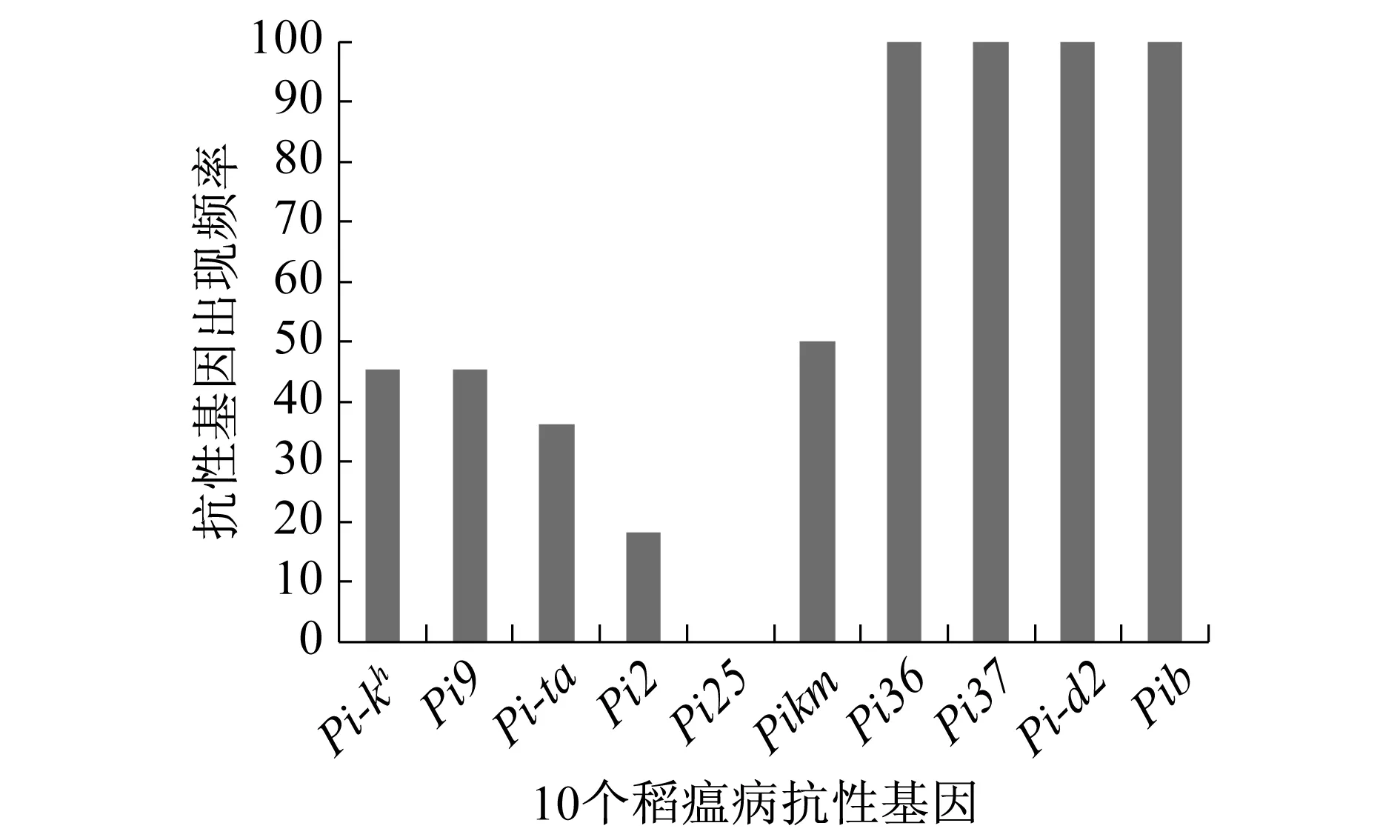
图2 10个稻瘟病基因型在22种水稻中出现的频率
3 讨 论
传统的水稻抗病育种主要依靠田间抗病性考察对目标植株进行筛选.这种育种方法既耗时耗力,同时因为太受环境条件影响也很难达到理想的效果.分子标记辅助选择技术作为随着分子生物学及生物信息学技术发展而产生的新选择手段,因其具有准确、快速,且不受环境影响等优点,已经广泛应用于作物育种实践.实践证明,将多个抗菌谱不同的抗稻瘟病基因聚合到同一个水稻品种中,是培育广谱持久抗稻瘟病品种、应对稻瘟病菌高度易变性的有效策略.
本研究利用已经克隆的10个稻瘟病抗性基因建立的特异性分子标记,对上海市目前主要推广及新培育的22种水稻品种或品系进行检测.结果表明,这22种水稻均含Pi36、Pi37、Pi-d2、Pib4个抗性基因;Pi-kh、Pi9、Pi2、Pi-ta、Pikm抗性基因在22 种水稻中各有不同类型;所有检测品种或品系中均不具有Pi25抗性基因;而且Pi2基因在上海市22种水稻中出现的频率也比较低.由此建议可通过分子标记辅助方法将Pi25基因引入上海市水稻品种或品系,以及将Pi2基因更多地引入上海市水稻品种或品系,可以更全面地提高上海市水稻对不同稻瘟病生理小种的抗性效果.
稻瘟病基因Pi54,最初被命名为Pi-kh[12],因此Pi54和Pi-kh属同一抗病基因.现有报道显示,目前已克隆的较多稻瘟病抗性基因,不仅有较多基因会成簇分布在一个染色体的特定区域,每个基因位点还有较多的等位形式.如Pi9、Pi2、Pi50、Pigm、Piz、Piz-t同为Piz位点上的复等位基因,Pikh、Pikm、Pikp、Piks、Pikg与Pik也是互为等位基因[16].本研究中,对于有些水稻在Pi2基因和Pigm基因的检测中呈现杂合状态,也可能是存在有不同的等位形式所致.
王忠华等[18]认为,是否存在Pi-ta抗病基因与田间抗性相吻合.杨杰等[19]报道发现,曾经和目前在上海市主栽的一些“秀水”系列水稻,如“嘉花1号”、“秀水123”、“秀水134”都同时含有Pib、Pi-ta两个抗性基因,并认为由此赋予这些水稻具有较强抗稻瘟病表型.这与本研究检测结果相同.根据田间分析发现,含有Pi-ta抗病基因与水稻抗穗瘟之间存在极显著相关性,并且比Pib、Pikm和Pi54抗性基因具有相对更强的抗病效果[12].在本研究检测的22种水稻中,只有8种水稻含有Pi-ta抗病基因.为了使更多上海市水稻品种及品系能够具备抵抗穗瘟效果,可以利用分子标记辅助技术将Pi-ta抗病基因进行导入操作.
李洪亮等[20]研究认为,Pi2基因对稻瘟病抗性略强于Pi1基因,但两者单独存在时的水稻抗病效果都不是很理想,而对于同时含有Pi1和Pi2两个基因的水稻,对稻瘟病的抗性效果明显好于Pi1或Pi2单独存在的水稻.含有Pi2抗性基因对稻瘟病的抗性效果也好于Pitz抗性基因[10].张羽等[21]研究发现,Pi9、Pi-ta、Pikm、Pib、Piz-t这5个抗性基因位点对水稻的抗病影响力基本一致,并且可能都属于微效基因,它们各自单独存在时,对稻瘟病抗性效果都较弱,但是,随着抗性基因位点数量的增多,品种的抗病性呈上升趋势.由此表明,对于高抗稻瘟病水稻育种,有必要将多个抗性基因聚合到一个水稻品种中.
本研究检测结果表明,上海市大多主推水稻品种或一些新培育的水稻品系,都具有较多种抗稻瘟病基因,这可能从一定程度上反映出这些水稻品种或品系具有较好的稻瘟病抗性水平,但这些水稻的实际抗病效果还需要通过田间检测进行验证.开展本研究,也为今后利用分子标记辅助更进一步在上海市水稻品种或品系中聚合更多的稻瘟病抗性基因研究奠定了基础.
[1] 李彬,邓元宝,颜学海,等.一个粳稻来源抗稻瘟病基因的鉴定、遗传分析和基因定位 [J].作物学报,2014,40(1):54-62.
Li B,Deng Y H,Yan X H,et al.Indentification genetic analysis and gene mapping of a rice blast resistance gene inJaponicarice [J].Acta Agronomica Sinica,2014,40(1):54-62.
[2] 赵正洪,张岳平,瞿华香.水稻稻瘟病菌及其抗性遗传研究进展 [J].湖南农业科学,2008(5):1-4.
Zhao Z H,Zhang Y P,Qu H X.Research progress on resistance inheritance of rice blast fungus [J].Hu′nan Agricultural Sciences,2008(5):1-4.
[3] 朱小丽,陶跃之,周波.水稻抗稻瘟病基因的复等位分布特征及其在基因挖掘中的应用 [J].浙江农业学报,2015,27(10):1789-1796.
Zhu X L,Tao Y Z,Zhou B.Distribution pattern of mutiple alleles of rice blast resistance genes and its application in gene cloning [J].Acta Agriculturae Zhejiangensis,2015,27(10):1789-1796.
[4] Hittalmani S,Parco A,Mew T V,et al.Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice [J].Theoretical and Applied Genetics,2000,100(7):1121-1128.
[5] Narayanan N N,Baisakh N,Oliva N P,et al.Molecular breeding:Marker-assisted selection combined with biolistic transformation for blast and bacterial blight resistance in indica rice (cv.CO39) [J].Molecular Breeding,2004,14(1):61-71.
[6] 陈红旗,陈宗祥,倪深,等.利用分子标记技术聚合3个稻瘟病基因改良金23B的稻瘟病抗性 [J].中国水稻科学,2008,22(1):23-27.
Chen H Q,Chen Z X,Ni S,et al.Pyramiding three genes with resistance to blast by marker-assisted selection to improve rice blast resistance of Jin 23B [J].Chinese Journal of Rice Science,2008,22(1):23-27.
[7] 倪大虎,易成新,李莉,等.利用分子标记辅助选择聚合水稻基因Xa21和Pi9(t) [J].分子植物育种,2005,3(3):329-334.
Ni D H,Yi C X,Li L.et al.PyramidingXa21 andPi9(t) in rice by marker-assisted selection [J].Molecular Plant Breeding,2015,3(3):329-334.
[8] 陈健民,付志英,王锋,等.分子标记辅助培育双抗稻瘟病和白叶枯病杂交稻恢复系 [J].分子植物育种,2009,7(3):465-470.
Chen J M,Fu Z Y,Wang F,et al.Breeding hybrid rice restoring line with double resistance to rice blast and bacterial blight by marker-assisted selection [J].Molecular Plant Breeding,2009,7(3):465-470.
[9] 王军,杨杰,朱金燕,等.稻瘟病抗病基因Pi-kh功能标记的开发及江苏粳稻品种中Pi-kh的变异 [J].中国水稻科学,2014,28(2):141-147.
Wang J,Yang J,Zhu J Y,et al.Development of functional marker for rice blast resistance genePi-khand natural variation atPi-khlocus in japonica rice in Jiangsu Province,China [J].Chinese Journal of Rice Science,2014,28(2):141-147.
[10] 高利君,高汉亮,颜群,等.4个抗稻瘟病基因分子标记的建立及在水稻亲本中的分布 [J].杂交水稻,2010,25(S1):294-298.
Gao L Q,Gao H L,Yan Q,et al.Establishment of markers for four blast genes and marker distribution in rice parents [J].Hybrid Rice,2010,25(S1):294-298.
[11] 王惠梅,陈洁,施勇烽,等.稻瘟病抗性基因Pi25特异性CAPS标记的开发与验证 [J].作物学报,2012,38(11):1960-1968.
Wang H M,Chen J,Liu Y F,et al.Development and validation of CAPS markers for marker-assisted selection of rice blast resistance genePi25 [J].Acta Agronomica Sinica,2012,38(11):1960-1968.
[12] 范方军,王芳权,刘永峰,等.Pi-b,Pi-ta,Pikm和Pi54对水稻穗颈瘟的抗性评价 [J].华北农学报,2014,29(3):221-226.
Fan F J,Wang F L,Liu Y F,et al.Evaluation of resistance to rice panicle blast with resistant genesPi-b,Pi-ta,PikmandPi54 [J].Acta Abriculturae Boreali-Sinica,2014,29(3):221-226.
[13] 宋广树,孙蕾,刘志全,等.吉林省主要水稻品种稻瘟病抗性分析及分子辅助改良 [J].西北农业学报,2016,25(5):677-683.
Song G S,Sun L,Liu Z Q,et al.Rice blast resistance analysis and molecular assistant improvement of main rice varieties in Jilin Province [J].Acta Agriculturae Boreali-occidentalis Sinica,2016,25(5):677-683.
[14] 刘洋,徐培洲,张红宇,等.水稻抗稻瘟病Pib基因的分子标记辅助选择与应用 [J].中国农业科学,2008,41(1):9-14.
Liu Y,Xu P Z,Zhang H N,et al.Marker-assisted selection and application of blast resistant genePibin rice [J].Chinese Journal of Rice Science,2008,41(1):9-14.
[15] Zhai C,Lin F,Dong Z Q,et al.The isolation and characterization ofPik,a rice blast resistance gene which emerged after rice domestication [J].New Phytologist,2011,189(1):321-334.
[16] 何秀英,王玲,吴伟怀,等.水稻稻瘟病抗性基因的定位、克隆及育种应用研究进展 [J].中国农学通报,2014,30(6):1-12.
He X Y,Wang L,Wu W H,et al.The progress of mapping,isolation of the genes resisting to blast and their breeding application in rice [J].Chinese Agricultural Science Bulletin,2014,30(6):l-12.
[17] Chen H L,Chen B T,Zhang D P,et al.Pathotypes ofPyriculariagriseain rice fields of central and southern China [J].Plant Disease,2001,85(8):843-850.
[18] 王忠华,贾育林,吴殿星,等.水稻抗稻瘟病基因Pi-ta的分子标记辅助选择 [J].作物学报,2004,12,30(12):1259-1265.
Wang Z H,Jia Y L,Wu D X,et al.Molecular markers-assisted selection of the rice blast resistance genePi-ta[J].Acta Agronomica Sinica,2004,30(12):1259-1265.
[19] 杨杰,杨金欢,王军,等.稻瘟病抗病基因Pita和Pib在中国水稻地方品种中的分布 [J].华北农学报,2011,26(3):1-6.
Yang J,Yang J H,Wang J,et al.Distribution of two blast resist ant genesPitaandPibin landrace rice in China [J].Acta Abriculturae Boreali-Sinica,2011,26(3):1-6.
[20] 李洪亮,李荣田.稻瘟病抗性基因Pi1和Pi2的聚合及其育种价值分析 [J].北方水稻,2010,40(5):7-12.
Li H L,Li R T.The polymerization of rice-blast resistance genesPi1 andPi2 and analysis on breeding value [J].North Rice,2010,40(5):7-12.
[21] 张羽,冯志峰,张先平,等.5个稻瘟病抗性基因的基因型分型与抗性评价研究 [J].西北农业学报,2014,27(5):1929-1936.
Zhang Y,Feng Z F,Zhang X P,et al.Study on genotyping and resistance evaluation for five rice blast resistance genes [J].Southwest China Journal of Agricultural Sciences,2014,27(5):1929-1936.
(责任编辑:顾浩然,郁 慧)
Genotypingof10blastresistancegenesin22ricevarietiesbymolecularmarker
Han Xiao, Heye Chenxing†, Zhang Mei†, Lei Ziyi†, Zhang Tao, Li Jianyue*
(Development Center of Plant Germplasm Resources,College of Life and Environmental Sciences,Shanghai Normal University,Shanghai 200234,China)
Genotyping of 10 blast resistance genes,which were cloned from 22 leading rice cultivars or new bred rice strains grown mainly in Shanghai,was carried out by using molecular marker detection technology for clearing these genes′ distribution in the 22 rice cultivars or strains.The results showed that 22 rice cultivars or strains all contained rice blast resistance genesPi36、Pi37、Pi-d2 andPib,while the distributions ofPi-kh、Pi9、Pi2、Pi-taandPikmwere different in above different rice cultivars or strains.And all the rice samples detected did not containPi25.
rice; molecular marker; blast resistance gene
Q 75
A
1000-5137(2017)05-0654-08
2017-08-23
上海市科委现代农业科技创新项目(16391900800);上海植物种质资源工程技术研究中心项目(17DZ2252700)
韩 笑(1996-),女,在读本科生.
导师简介: 李建粤(1958-),女,教授,主要从事植物分子遗传学方面的研究.E-mail:lijianyue01@shnu.edu.cn
*
†在完成本研究试验操作过程具有同等贡献,并列第二作者.
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29
农技服务(2023年2期)2023-03-15
今日农业(2022年4期)2022-06-01
作物学报(2022年6期)2022-04-08
园林科技(2020年2期)2020-01-18
浙江农业学报(2017年1期)2017-05-17
河南农业(2016年6期)2016-11-26
现代农业(2016年5期)2016-02-28
现代农业(2015年1期)2015-02-28
作物研究(2014年6期)2014-03-01
