
NaCl胁迫对不同白榆品系生物量及光合作用的影响
2017-11-23 02:33:52张晓晓殷小琳李红丽贾淑友
生态学报 2017年21期
张晓晓, 殷小琳,李红丽,*,苏 丹,贾淑友, 董 智
1 山东农业大学林学院/山东省土壤侵蚀与生态修复重点实验室/泰山森林生态站, 泰安 271018 2 中国水利水电科学研究院, 北京 100038
NaCl胁迫对不同白榆品系生物量及光合作用的影响
张晓晓1, 殷小琳2,李红丽1,*,苏 丹1,贾淑友1, 董 智1
1 山东农业大学林学院/山东省土壤侵蚀与生态修复重点实验室/泰山森林生态站, 泰安 271018 2 中国水利水电科学研究院, 北京 100038
以3种一年生白榆品系(Y65、Y1、Y34)为试验材料,通过测定5种浓度 NaCl处理(0、50、85、120、155 mmol/L)下其生物量、叶绿素含量及光合参数等指标的变化,探讨3个白榆品系的耐盐性差异及其光合参数的表征。结果表明:与对照相比,白榆品系生物量及叶绿素含量随NaCl胁迫增强逐渐降低,高NaCl胁迫(>85 mmol/L)会显著抑制生物量的累积及破坏叶绿素的合成。低NaCl胁迫(≤85 mmol/L)下,3个品系的光合机制以气孔限制为主,通过提高其气孔限制值而降低蒸腾作用,以提高水分利用效率(WUE)而适应盐分胁迫;而高NaCl胁迫(>85 mmol/L)下,则以非气孔限制为主,通过降低WUE而减少根系对地下水分与盐离子的吸收,以此来维持自身生长。3个白榆品系中,使非气孔限制转变为Pn下降主因的NaCl胁迫浓度不同,Y65、Y1的转折点为85 mmol/L,而Y34的转折点为50 mmol/L。结论:3个供试白榆品系中Y65的综合耐盐性较高,是盐碱地种植白榆品系的优先选择。
NaCl胁迫;白榆;光合特性;耐盐性
土壤盐渍化是植物生长中经常遇到的自然逆境之一,当土壤溶液中盐分离子超过一定含量时,不仅会对其生长产生抑制作用[1],而且还会引起植物体内生理生化、光合、离子积累等多方面的变化[2-3],并导致植物生物膜功能及结构失调[4],引起植物细胞内氧化还原系统失调、渗透胁迫、离子失衡和离子毒害等危害[5- 7]。通过前人对酸枣、柚、地被观赏竹及不同种源沙枣等耐盐性的研究发现[8- 11],随NaCl胁迫浓度的增加,生物量积累的抑制效应越来越明显,叶绿素含量下降幅度增大;光合机制由气孔限制为主转变为非气孔限制为主,净光合速率、蒸腾速率、气孔导度等光合指标均有不同程度的降低,总体表现为耐盐性相对较低的品种各指标的变化较为明显,而耐盐性较好的品种各指标的变化则相对稳定,即由于不同植物种类或同一种类不同品系的耐盐能力差异较大,NaCl胁迫条件下的生长、生理生化指标、光合特性等表现不尽相同。因此,研究植物的耐盐性对耐盐植物或品种的筛选和盐碱地植物恢复与重建具有重要的意义。
白榆(UlmuspumilaL.)是我国分布最为广泛和重要的速生阔叶用材树种,具有良好的耐旱、耐寒、耐盐碱和抗风能力,属优质的乔木造林树种。已有的研究表明,盐碱胁迫对白榆的生长量、光合性能、游离脯氨酸、可溶性糖的变化特征均有影响[12- 15];采用增殖率与苗高生长量[13]、光合和荧光有效指标以及苗木受害情况[12]的评价表明,不同盐碱胁迫下白榆的耐盐性表现为强耐盐碱型、弱耐盐碱型和对盐碱胁迫敏感型3种响应方式。基于白榆不同品系间耐盐性的差异,本文以3个白榆品系为试材,通过5种不同浓度NaCl处理,研究NaCl胁迫下幼苗生物量积累、叶绿素及光合特性的变化,探讨不同白榆品系对NaCl胁迫的响应特征与方式,初步评价不同白榆品系的耐盐性,为盐碱地耐盐植物种的选育提供科学依据。
1 材料与方法
1.1 试验地概况
山东农业大学林学试验站位于泰安市的南部,东经117°11′,北纬36°16′。属于温带季风大陆性气候,年平均气温12.8 ℃,气温变幅为-6.9—37 ℃,年大于10 ℃的积温为4283.1 ℃,无霜期186.6 d。年降水量600—700 mm,年均相对湿度65%。实验在山东农业大学林学试验站的大棚内进行,大棚四周通风,保持棚内温湿度与自然条件基本一致。
1.2 供试材料
3个供试白榆品系(Y65、Y1、Y34)为山东省林业科学研究院提供的组培苗经大田炼苗后定植至盆内。3个品系的外植体采自于山东省金乡县白洼林场(原林业部和山东省林业厅确定的林木良种基地)白榆基因库,是从220个白榆种质资源中选取40个生长量大、抗病虫能力较强的白榆优良无性系外植体,再经耐盐组织培养获得的具有抗盐能力的21个白榆品种的其中3个。与一般榆树相比,具有生长快、抗盐碱、抗病虫害、树干直、树形好等特性。2014年3月,把供试材料品系定植于直径20cm、深40cm的花盆中,花盆底部衬以无纺布防漏水漏土;以普通壤土、细沙与泥碳土按2∶1∶1的干质量比配置栽培基质,装至盆高3/4处,定期浇水和营养液,保证其成活和生长。试验时间为 2014年7月—2014 年9月。
1.3 试验设计
2014年7月,选择长势一致、无病虫害的健康植株,以0 mmol/L为对照,分别配制质量体积比为50、85、120、155 mmol/L 4种NaCl浓度梯度进行处理,每个处理6个重复,共计90盆。盐处理采用浸泡法,即将盆的3/4至4/5浸入盐水通过底部开孔吸取盐水,每周处理2—3次,每次6—10 h左右;对照组采用清水浸泡代替盐水。为避免盐激效应,对于120、155 mmol/L盐处理,先给予85 mmol/L处理,然后经3次处理,逐次升高NaCl浓度至处理浓度。每经3次浸泡用自来水多次洗盐以消除花盆中的盐分积累,盆栽苗洗盐静置3 h后,随即再进行盐水浸泡处理,如此反复处理。为保证苗木正常生长,对照组与不同盐处理组均按体积加入25% Hoagland作为营养液。除正常盐水浸泡处理、洗盐过程之外,试验苗不额外浇水,不进行人为修剪。待各NaCl浓度处理均1个月后,开始各项指标测定。
1.4 测定内容和方法
1.4.1 生物量测定
盐处理3 个月后,待所有实验测定结束后,将盆栽苗带回实验室内,以自来水冲松盆内土壤,轻轻取出苗木,以自来水冲洗根系和地上部分,去除沙粒及其他表面杂物,再用去离子水洗净,吸干水分,将植株地上部分与地下部分分开,于105 ℃杀青30 min,之后在 75 ℃下烘至恒重,称取干重。
1.4.2 光合特性参数的测定
采用美国PP Systems公司生产制造的CIRAS- 2 光合仪,选晴朗无风的日子,于9:00—11:00选取每个树种中长势健壮的苗木,取其生长良好的中上部叶片,测定苗木的净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)及蒸腾速率(Tr)。测定时采用开放式气路,CO2浓度约为385 μmol/L 左右,选择红蓝光源叶室,设定光合有效辐射(PAR) 为1000 μmol m-2s-1。每个品系不同盐处理浓度各测定3盆,每盆测定3个叶片,每个叶片重复测定3次。光合水分利用效率(WUE)和气孔限制值(Ls)采用以下公式计算:
WUE=Pn/Tr
Ls=1-Ci/Ca
式中,Ci为细胞间隙CO2浓度,Ca为测量时空气中CO2浓度。
1.4.3 叶绿素含量的测定
与测定光合特性参数同步,摘取供试白榆品系的中上部叶片,迅速放入液氮罐,带回实验室,采用丙酮提取法[16],用分光光度计测定Chl(a+b)、Chla和Chlb的含量。
1.5 数据处理
通过Excel对实验数据进行计算和处理,采用SPSS 19.0统计软件利用单因素方差分析(ANOVA)对实验数据进行相关性分析及差异性显著性分析。
2 结果与分析
2.1 NaCl胁迫对不同白榆品系生物量的影响
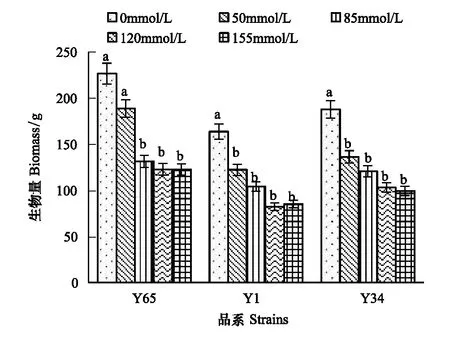
图1 NaCl胁迫对白榆品系生物量的影响 Fig.1 Effects of salt stress on biomass of Ulmus pumila L. strains不同小写字母表示处理间差异显著(P<0. 05)
图1显示,NaCl胁迫抑制了植物的生长发育,随着NaCl胁迫增强,3个白榆品系的生物量显著降低,但在不同NaCl浓度下的降低程度不同,Y65在50 mmol/L处理时与对照相比生物量降幅较小为16.71%,而Y1、Y34在此处理下生物量已明显降低,降幅分别达25.27%、27.45%。各品系在50—85 mmol/L NaCl胁迫时下降幅度较大,120—155 mmol/L NaCl浓度时生物量下降渐趋于平缓,155 mmol/L NaCl浓度时分别下降了45.79%、47.98%、47.07%。
2.2 NaCl胁迫对不同白榆品系叶绿素含量的影响
3个白榆品系的Chl(a+b)、Chla及Chlb含量对NaCl胁迫的响应表现出相似的变化趋势,均随着 NaCl处理浓度的增加而减小(图2)。NaCl浓度为155 mmol/L 时,Y65、Y1、Y34的Chl(a+b)含量分别为各自对照的58.68%、37.65%、48.96%。方差分析显示,同一品系不同盐处理间Chl(a+b)、Chla及Chlb含量差异显著(P<0.05)。不同品系间,各NaCl浓度处理下,Y65的Chl(a+b)含量降幅总体小于Y1、Y34,表明NaCl胁迫对Y65的Chl(a+b)含量变化影响较小。
Y65、Y1、Y34的Chla/b在50—155 mmol/LNaCl浓度处理下呈先升后降的总趋势(图2),在NaCl浓度为120 mmol/L 时均达到最大值,为各自对照值的144.46%、143.76%、144.20%,而在其余NaCl浓度下各品系的Chla/b值均小于对照值。方差分析表明,同一品系间,低NaCl胁迫(≤85 mmol/L)下,各处理间相比差异不显著;而高NaCl胁迫 (>85 mmol/L)下各NaCl浓度梯度与对照相比均差异显著(P<0.05)。

图2 NaCl胁迫对白榆品系叶绿素含量及组成的影响Fig.2 Effects of salt stress on chlorophyll contents and composition of Ulmus pumila L. strains
2.3 NaCl胁迫对不同白榆品系光合特性的影响
2.3.1 净光合速率(Pn)
由表1可知,在50 mmol/L NaCl浓度下,Y65的平均净光合速率达到8.57 μmol m-2s-1,比对照增加了5.76%,说明此时NaCl胁迫对Y65光合速率的促进作用达到显著水平(P<0.05);Y1、Y34的平均净光合速率均下降,Y1与对照值无显著差异,但Y34比对照降低了12.20%,差异明显(P<0.05),可见NaCl胁迫已对Y34起到了抑制作用。在NaCl胁迫≥85 mmol/L时,3个品系的净光合速率随着盐分浓度的增加而降低,NaCl浓度越高,下降越显著,在155 mmol/L NaCl浓度下比各自对照下降了86.42%、73.96%、88.98%,NaCl胁迫对品系净光合速率的抑制作用达到显著水平(P<0.05)。

表1 NaCl胁迫对白榆品系幼苗光合作用的影响
表中不同小写字母表示处理间差异显著(P<0.05)
2.3.2蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci) 及气孔限制值(Ls)
逆境中,植物通过改变气孔的开度等方式来调节与环境的CO2和水汽交换,进而调节光合速率和蒸腾速率,以适应逆境条件。从表1知,不同NaCl浓度下,3个品系的Tr与Gs没有显著的规律性,但其变化趋势呈线性相关。低NaCl胁迫(≤85 mmol/L)下,Y65的Tr、Gs、Ci变化趋势一致,均先升后降,Ls先降后升,且各处理间差异显著(P<0.05),在50 mmol/L NaCl浓度下显著高于对照值,而在85 mmol/L NaCl浓度下降,显著低于对照值。Y1的Tr、Gs、Ci及Ls的变化趋势与Y65一致,但其变化幅度要大于Y65;Y34的Tr、Gs、Ci均随NaCl胁迫的增强而降低,处理间差异不显著。高NaCl胁迫(>85 mmol/L)下, Y1、Y34的Tr、Gs显著降低,Ci则呈升高趋势,Ls降低缓慢;而Y65的Tr、Gs下降并不明显,差异不显著,Ci则呈显著升高,Ls显著降低。结合Pn的变化趋势来看,Y65、Y1在盐浓度从50 mmol/L增大至85 mmol/L时,Pn、Tr与Gs、Ci均降低,而Ls则升高,表明引起光合参数变化的主要因素为气孔限制造成的;而当盐浓度>85 mmol/L时,随着NaCl胁迫继续加剧,Pn和Ls都降低,而Ci上升,此时Pn下降的主要原因已经转变为非气孔因素的限制。Y34则表现为NaCl盐浓度>50 mmol/L时,非气孔限制已成为光合参数下降的主导原因。说明3个品系Tr、Gs、Ci和Ls的不同变化趋势反映了三者间耐盐性与对盐分适应机制的差异。
2.3.3 水分利用率(WUE)
Y65、Y1的水分利用率(WUE)在低NaCl胁迫(≤85 mmol/L)下均先降后升,在85 mmol/L时达到最大值,为各自对照值的119.95%、257.61%;在高NaCl胁迫下(>85 mmol/L),Y65的WUE值显著降低,在155 mmol/L时仅为对照的11.87%,Y1的WUE值也有所降低,但与其对照值差异不显著。Y34的WUE在50 mmol/L NaCl浓度时已达到最大值,但小于Y65及Y1的最大值,为对照的193.57%,可知50 mmol/L NaCl浓度对其水分利用率具有一定促进作用,而后随盐浓度NaCl浓度升高而下降,在155 mmol/L时下降到2.22,显著(P<0.05)低于对照。
2.4 不同参数的相关分析及隶属函数分析
对不同NaCl胁迫下3个白榆品系的11个指标进行相关性分析,结果由表2可知,与生物量呈极显著正相关(P<0.01)的指标有叶绿素含量、chla含量、净光合速率及气孔导度;蒸腾速率与生物量呈显著正相关(P<0.05),即NaCl胁迫下可通过调控以上5个指标直接影响白榆品系的生物量累积,提高植物的耐盐性。Ls与WUE呈极显著正相关(P<0.01),但二者与生物量无显著相关性,对植物的生长代谢起间接的影响作用。
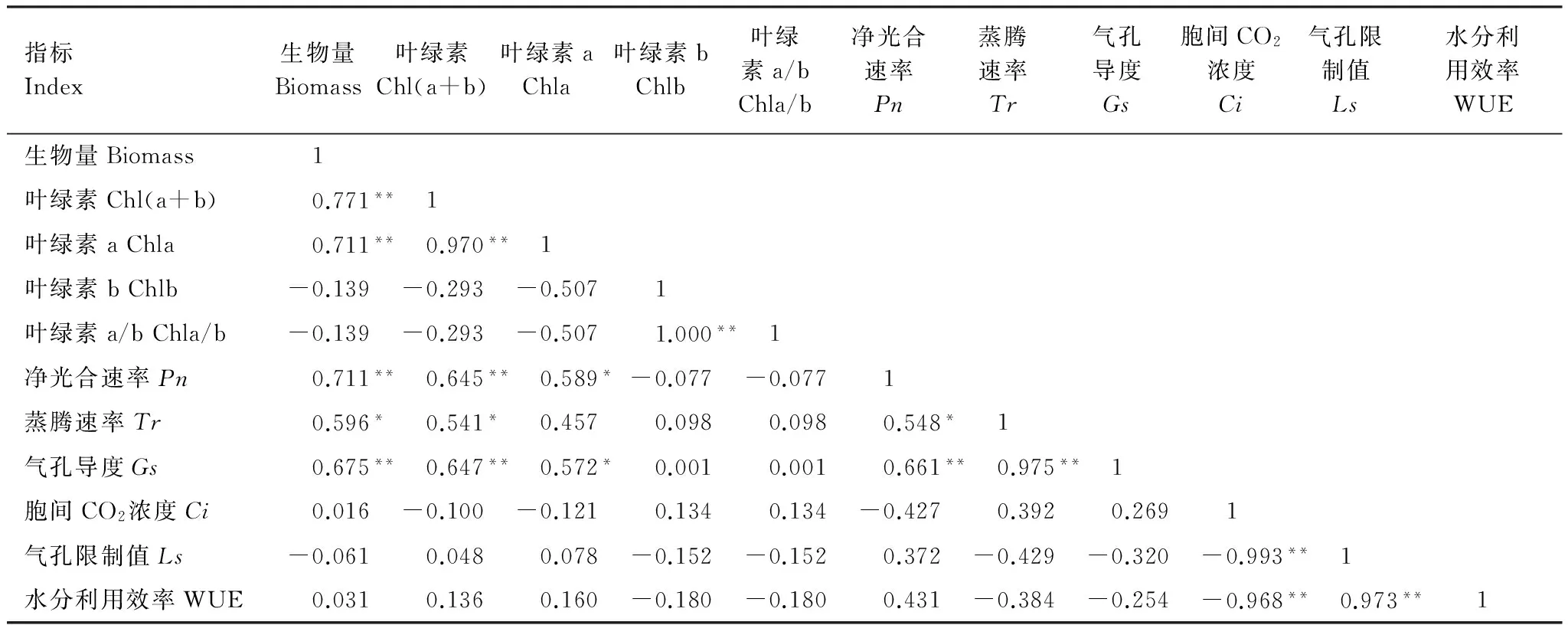
表2 NaCl胁迫下白榆品系各指标间的相关性分析
Chl(a+b):叶绿素(a+b) Chlorophyll (a+b);Chla:叶绿素a Chlorophyll a;Chlb:叶绿素b Chlorophyll b;Chla/b:叶绿素a/b Chlorophyll a/b;Pn:净光合速率Net photosynthetic rate;Tr:蒸腾速率Transpiration rate;Gs:气孔导度Stomatal conductance;Ci:胞间CO2浓度Intercellular CO2concentration;Ls:气孔限制值Stomatal limitation;WUE:水分利用效率Water use efficiency;*表示显著相关(P<0.05),**表示极显著相关(P<0.01)

表3 3个白榆品系耐盐性综合评定
利用隶属函数法对品系各指标进行耐盐性综合评价,结果如表3。由表可知,3个品系耐盐性综合排序为Y65>Y1>Y34。
3 讨论与结论
土壤中的过量盐分会对植物造成渗透胁迫并干扰其营养离子的平衡,进而影响植物的生长、光合作用、渗透调节物质的合成、脂类代谢等过程,最终影响到植物生物量的累积。生物量是植物对NaCl胁迫反应的综合体现,即对NaCl胁迫的综合适应,也是植物耐盐性的直接指标[17-18]。因此,生物量累积值可为植物耐盐性的评估提供可靠的标准。本实验中,3个白榆品系在不同浓度的NaCl胁迫条件下生物量减少,且随NaCl胁迫浓度的增加,生物量降低幅度增大。总体而言,Y65的生物量累积明显大于Y1、Y34,即Y65的耐盐能力高于另外两个品系。
叶绿素是植物进行光合作用的重要物质,在光能的吸收和转换中起着重要作用。本研究中,随着NaCl胁迫增加,白榆品系叶片的Chl(a+b)、Chla、Chlb均降低,可能是NaCl胁迫下叶绿素酶活性增强,促进了叶绿素的降解。相关分析表明,叶绿素含量与净光合速率和植物生长量呈极显著正相关(P<0.01),故叶绿素含量的降低,影响植物的光合作用速率,从而影响植物的生长发育及产量形成。Chla/b值可以反映叶绿体中不同垛叠状态的类囊体膜比例,逆境胁迫下植物Chla/b值变化越小,表明其对逆境的适应性越强[19]。本研究中,各白榆品系的Chla/b值均在2.0—5.0范围内波动。低NaCl胁迫(≤85 mmol/L)下,3个品系Chla/b值变化无显著差异;而在较高NaCl胁迫(120 mmol/L)下,Chla/b显著升高,促进了Chlb的降解,限制了捕光色素蛋白对光能的捕获,表明品系叶绿体存在通过降低对光能的捕获而避免盐害的一种适应性机制。这与谷艳芳等的较高NaCl胁迫导致冬小麦Chla/b提高的研究结果相似[20]。可见,NaCl胁迫对叶绿素的形成造成破坏,影响了叶绿素的合成与分解,进而影响植物的光合作用强度。NaCl胁迫下,3个白榆品系中Y65的Chla/b变化幅度及Chl(a+b)下降幅度最小,即在NaCl胁迫下Y65的叶绿素系统受损伤程度最小,耐盐性最好。
一般认为NaCl胁迫下,引起植物叶片光合效率降低的因素主要有由于气孔的部分关闭导致的气孔限制和叶肉细胞光合活性的下降导致的非气孔限制两类,判定依据主要是胞间 CO2浓度(Ci)、气孔导度(Gs)及气孔限制值(Ls)的变化方向[21- 24]。本试验中,白榆品系对不同NaCl胁迫的光合响应机制不同,低NaCl胁迫(≤85 mmol/L)下,3个品系的光合机制以气孔限制为主;高NaCl胁迫(>85 mmol/L)下,以非气孔限制为主。低 NaCl 胁迫(≤85 mmol/L)条件下,Y65、Y1的Gs、Pn与Ci变化一致,品系通过对自身Gs的调控来适应NaCl胁迫。50 mmol/L NaCl胁迫刺激叶片气孔张开Gs升高,同时与Gs线性相关的蒸腾速率Tr增加,蒸腾拉力增大,进一步促进叶片中水分及矿质营养的吸收和运输,CO2向叶绿体的输送速率提高,使得Ci增加,促进了净光合速率Pn升高。85 mmol/L NaCl胁迫下植物通过降低Gs,控制Tr和Ci,增加植物持水量,以此来适应胁迫,说明Pn变化主要受气孔限制因素的影响。当 NaCl 浓度在120—155 mmol/L时,Y65、Y1的Ci随Gs、Pn的下降而升高,而Ls降低,说明Pn下降的主要原因已由气孔因素限制转变为非气孔因素限制,高NaCl胁迫导致细胞中盐离子的大量累积,不但使植物体内水势降低,Tr下降;同时破坏了叶绿体结构,造成叶片光合器官的损伤,与此对应的Chlb的降解明显增强,Chla/b显著升高(图2),叶肉细胞的光合活性下降。这与刘伟成等[23]关于NaCl胁迫对花期海蓬子光合作用影响的研究结果一致。而Y34在NaCl胁迫大于50 mmol/L时,Gs、Pn均显著下降,Ci升高,Ls下降,表明非气孔限制已经成为主导因素,可知,不同品系对NaCl胁迫的适应程度不同。由此认为,85 mmol/L的NaCl浓度为Y65、Y1白榆品系叶片光合作用气孔与非气孔因素限制的转折点,而Y34的气孔限制与非气孔限制转折点出现在50 mmol/L的NaCl浓度。
植物水分利用效率 WUE(Pn/Tr) 是光合和蒸腾特性的综合反应,WUE 高的植物,其叶片含水量高,持水强,用水节约,限制叶片水分散失,减轻对光合的抑制,提高植物抗逆性[25]。3个白榆品系的WUE随NaCl浓度的增加,总体呈先增后降的变化趋势。根据Cowan[26]和Farquhar[24]等的观点,气孔在短期的行为总是以有限的水分丧失来换取最大的CO2同化量,即气孔的最优化理论。可见,Y65、Y1在50 mmol/L NaCl浓度下WUE的降低是品系对低NaCl胁迫的适应性反应,而85 mmol/L NaCl浓度下,蒸腾速率大幅降低,WUE显著高于对照值,表明适当的NaCl浓度对品系的水分利用率有显著的促进作用。高NaCl胁迫对植物的细胞膜产生破坏,植物Pn下降速率大于Tr的下降速率,通过降低WUE,从而减少根系对地下水分盐离子的吸收,以此来维持自身生长。不同NaCl浓度处理下,3个品系中,Y34的WUE显著小于Y65、Y1,表明其在NaCl胁迫下对水分的有效利用率较低,耐盐性相对较差。
本研究通过对NaCl胁迫处理下白榆品系的11个指标进行隶属函数综合分析,比较客观地反映了各参试品系的耐盐性。结果表明:Y65的耐盐性强于Y1、Y34,即Y65对盐碱地的综合适应能力要优于Y1、Y34。
[1] 王合云, 李红丽, 董智, 陈新闯. 滨海盐碱地不同土壤—树种系统中盐分离子分布与运移. 水土保持学报, 2014, 28(4): 222- 226, 242- 242.
[2] Allakhverdiev S I, Sakamoto A, Nishiyama Y, Inaba M, Murata N. Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II inSynechococcussp. Plant Physiology, 2000, 123(3): 1047- 1056.
[3] 鲁少尉, 齐飞, 李天来. NaCl及等渗PEG胁迫对番茄叶片光合特性及蔗糖代谢的影响. 华北农学报, 2012, 27(3): 136- 141.
[4] Ashraf M, Ali Q. Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (BrassicanapusL.). Environmental and Experimental Botany, 2008, 63(1/3): 266- 273.
[5] 鲁艳, 雷加强, 曾凡江, 徐立帅, 彭守兰, 刘国军. NaCl处理对胡杨生长及生理生态特征的影响. 干旱区研究, 2015, 32(2): 279- 285.
[6] Jaleel C A, Lakshmanan G M A, Gomathinayagam M, Panneerselvam R. Triadimefon induced salt stress tolerance inWithaniasomniferaand its relationship to antioxidant defense system. South African Journal of Botany, 2008, 74(1): 126- 132.
[7] Azooz M M, Shaddad M A, Abdel-Latef A A. The accumulation and compartmentation of proline in relation to salt tolerance of three sorghum cultivars. Indian Journal of Plant Physiology, 2004, 9(1): 1- 8.
[8] 布阿依夏姆·依提. NaCl胁迫下酸枣种子、一年生实生苗及二年生移栽苗的生理特性研究[D]. 乌鲁木齐: 新疆农业大学, 2013.
[9] 马翠兰. 柚(CitrusgrandisOsbeck)对盐胁迫的生理反应及适应性研究[D]. 福州: 福建农林大学, 2002.
[10] 李善春. NaCl盐胁迫下5种地被观赏竹生理特性的研究[D]. 南京: 南京林业大学, 2005.
[11] 杨升. NaC1胁迫下不同种源沙枣幼苗耐盐性差异生理机制研究[D]. 北京: 中国林业科学研究院, 2014.
[12] 李庆贱, 陈志强, 时瑞亭, 张建秋, 王国柱, 张玉玲, 李悦. 白榆家系苗期耐盐碱研究. 北京林业大学学报, 2010, 32(5): 74- 81.
[13] 慕德宇. 离体培养条件下12个白榆优良无性系氯化钠盐分抗性筛选的研究. 山东大学学报: 理学版, 2013, 48(3): 19- 23.
[14] 刘炳响, 王志刚, 杨敏生, 梁海永. 模拟盐胁迫对白榆种子发芽、出苗及幼苗生长的影响. 草业学报, 2012, 21(5): 39- 46.
[15] 刘炳响, 王志刚, 梁海永, 杨敏生. 盐胁迫对不同生境白榆生理特性与耐盐性的影响. 应用生态学报, 2012, 23(6): 1481- 1489.
[16] 李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000.
[17] Vicente O, Boscaiu M, Naranjo M, Estrelles E, Bellés J M, Soriano P. Responses to salt stress in the halophytePlantagocrassifolia(Plantaginaceae). Journal of Arid Environments, 2004, 58(4): 463- 481.
[18] 朱义, 谭贵娥, 何池全, 崔心红, 张群.. 盐胁迫对高羊茅(Festucaarundinacea)幼苗生长和离子分布的影响. 生态学报, 2007, 27(12):5447- 5454.
[19] 赵昕, 吴雨霞, 赵敏桂, 何建新. NaCl胁迫对盐芥和拟南芥光合作用的影响. 植物学报, 2007, 24(2): 154- 160.
[20] 谷艳芳, 丁圣彦, 李婷婷, 王欣, 李俊娇, 张丽霞. 盐胁迫对冬小麦幼苗干物质分配和生理生态特性的影响. 生态学报, 2009, 29(2): 840- 845.
[21] 管志勇, 陈发棣, 滕年军, 陈素梅, 刘浦生. 5种菊花近缘种属植物的耐盐性比较. 中国农业科学, 2010, 43(4): 787- 794.
[22] 裴斌, 张光灿, 张淑勇, 吴芹, 徐志强, 徐萍. 土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响. 生态学报, 2013, 33(5): 1386- 1396.
[23] 刘伟成, 郑春芳, 陈琛, 彭益全, 曾国权, 冀德伟, 陈少波, 谢起浪, 於俊琦. 花期海蓬子对盐胁迫的生理响应. 生态学报, 2013, 33(17): 5184- 5193.
[24] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33(1): 317- 345.
[25] 张继澍. 植物生理学. 北京: 高等教育出版社, 2006: 105- 142.
[26] Cowan I R. Regulation of Water Use in Relation to Carbon Gain in Higher Plants//Lange O L, Nobel P S, Osmond C B, Ziegler H, eds. Physiological Plant Ecology II. Springer Berlin Heidelberg, 1982: 589- 613.
EffectofsaltstressonthebiomassandphotosyntheticcharacteristicsofUlmuspumilaL.strains
ZHANG Xiaoxiao1, YIN Xiaolin2, LI Hongli1,*, SU Dan1, JIA Shuyou1, DONG Zhi1
1ForestryCollegeofShandongAgriculturalUniversity,ShandongProvincialKeyLaboratoryofSoilErosionandEcologicalRestoration,TaishanForestEcosystemResearchStation,Tai′an271018,China2ChinaInstituteofWaterResourcesandHydropowerResearch,Beijing100038,China
Salt stress is one of the major environmental factors affecting plants; it causes many changes in plant metabolism, such as changes in the content and composition of chlorophyll and reduction in the capacity and efficiency of photosynthesis. Salt stress also inhibits biomass accumulation. Therefore, plant biomass, photosynthesis, and chlorophyll content are often used as indicators of salt stress injury in plants.UlmuspumilaL. is a widely distributed and important fast-growing hardwood timber species that has good tolerance to drought, cold, salt, and wind. It is the most important timber species in China′s Northern regions, saline-alkali areas, and desert, where it is used for afforestation, and to provide shelter forest and greenery.Consequently, it is considered to be a promising plant for cultivation in saline land. In response to salt stress, plants have evolved diverse mechanisms that can mitigate the effects of stress and lead to improved plant tolerance. In this study, 1-year-old potted seedlings ofUlmuspumilaL. strains (Y65, Y1, and Y34) were used to examine their changes in chlorophyll content, photosynthetic characteristics, and biomass accumulation under different concentrations of NaCl (CK, 50 mmol/L, 85 mmol/L, 120 mmol/L, and 155 mmol/L). The results showed that the content of chlorophyll and biomass of three theUlmuspumilaL. strains were significantly inhibited under higher-concentration salt stress, whereas no obvious changes were observed for the biomass accumulation of Y65 under lower concentrations of NaCl. Under low salinity stress (NaCl ≤85 mmol/L), the values for leaf photosynthetic rate (Pn), stomatal conductance (Gs), and intercellular CO2concentration (Ci) were decreased, whereas stomatal limitation (Ls) was increased. Stomatal inhibition (or stomatal restriction) was the main cause of the reduction inPnofUlmuspumilaL. strains. Although no stomatal restriction occurred in the 120 mmol/L and 155 mmol/L NaCl treatments, the values forPnandLswere decreased, whereasCiwas increased. Under low-salinity stress (NaCl ≤85 mmol/L), the increase inLsresulted in a decrease in transpiration rate (Tr) and an increase in water use efficiency (WUE) that could reduce the leaf water loss, which alleviated the imbalance between water supply and demand and improved the salt-tolerance ofUlmuspumilaL. strains. Under high-salinity stress (NaCl >85 mmol/L), the absorption capacity of roots was decreased, and both leafLsand WUE decreased, which resulted in a more severe water imbalance. Correlation analysis indicated that the biomass ofUlmuspumilaL. strains was highly significantly positively related to total chlorophyll, chlorophyll a,Pn,Gs, and significantly positively related toTr. Furthermore, WUE was significantly positively related toLs. Collectively, our results indicated that the salt resistance of Y65 was better than that of Y1 and Y34. We accordingly recommend that Y65 is the preferredUlmuspumilaL. strains for afforestation in saline areas, which could provide a scientific basis for further research on the salinity tolerance mechanism.
NaCl stress;UlmuspumilaL.; photosynthetic characteristics; salt tolerance
中国水利水电科学研究院科研专项(SE0145C112015);世界银行贷款山东生态造林项目(SEAP-JC- 2)
2016- 08- 09; < class="emphasis_bold">网络出版日期
日期:2017- 07- 11
*通讯作者Corresponding author.E-mail: lhl@sdau.edu.cn
10.5846/stxb201608091632
张晓晓, 殷小琳,李红丽,苏丹,贾淑友, 董智.NaCl胁迫对不同白榆品系生物量及光合作用的影响.生态学报,2017,37(21):7258- 7265.
Zhang X X, Yin X L, Li H L, Su D, Jia S Y, Dong Z.Effect of salt stress on the biomass and photosynthetic characteristics ofUlmuspumilaL. strains.Acta Ecologica Sinica,2017,37(21):7258- 7265.
猜你喜欢
河南农业科学(2020年2期)2020-03-11 10:44:04
新课程·下旬(2017年7期)2017-08-14 11:12:50
现代园艺·综合版(2017年1期)2017-07-14 18:42:57
防护林科技(2017年9期)2017-04-05 22:56:14
上海农业学报(2016年2期)2016-10-27 00:49:35
萤火(2016年6期)2016-06-18 05:22:26
浙江农业学报(2016年7期)2016-06-15 20:30:08
现代农业科技(2015年17期)2015-11-27 17:44:30
西北林学院学报(2015年5期)2015-01-03 05:12:20
中国糖料(2013年4期)2013-03-02 12:22:47
