
231份烤烟种质资源SSR标记遗传多样性及其与农艺性状和化学成分的关联分析
2017-11-16 05:52童治军陈学军方敦煌曾建敏吴兴富肖炳光
中国烟草学报 2017年5期
童治军,陈学军,方敦煌,曾建敏,吴兴富,肖炳光
云南省烟草农业科学研究院/烟草行业烟草生物技术育种重点实验室/国家烟草基因工程研究中心,昆明 650021
农艺与调制
231份烤烟种质资源SSR标记遗传多样性及其与农艺性状和化学成分的关联分析
童治军,陈学军,方敦煌,曾建敏,吴兴富,肖炳光
云南省烟草农业科学研究院/烟草行业烟草生物技术育种重点实验室/国家烟草基因工程研究中心,昆明 650021
本文旨在分析烤烟种质资源遗传多样性,通过关联分析寻找与烤烟农艺性状和化学成分相关联的分子标记,为分子标记辅助选择育种和提高烤烟育种效率奠定基础。利用均匀分布于烟草24个连锁群上的120个SSR标记分析了231份烤烟种质资源的遗传多样性及群体遗传结构,在此基础上采用TASSEL 5.0 GLM(general linear model)和 MLM (mixed linear model)方法进行标记与表型性状变异(8个农艺性状和5个化学成分)的关联分析。通过遗传多样性分析,共检测出403个等位变异,变异范围2-21个,平均每个标记3.36个;引物的多态性信息含量(PIC)为0.036~0.785,平均值为0.408;供试材料间遗传距离为0.003~0.371,平均值为0.148。通过非加权组平均法(UPGMA)进行聚类分析,在遗传距离(GD)值为0.153水平上可将231份烤烟材料聚为4大类。通过群体遗传结构分析将供试材料划分成3个亚群,且每个亚群分别包含117、38和76份烤烟材料。通过GLM_Q和MLM_Q+K两种关联分析模型,分别检测出17个和9个与13个表型性状显著相关,其中9个标记在两个年份和两种模型中均被检测到与同一性状在P<0.001水平上极显著相关,且MLM_Q+K未能检测到新的关联标记;3个株高相关标记(TM10481、PT54964和TM20580)、2个叶数相关标记(TM10846和PT60728)、1个茎围相关标记(TM25276)和1个叶长相关标记(TM11215)与已报道的QTL定位结果一致,可应用于烤烟分子标记辅助选择育种。
烤烟;遗传多样性;关联分析;农艺性状;化学成分
烤烟(Flue-cured Tobacco)是世界上栽培面积最大的烟草类型,也是中国重要的经济作物和大宗出口的农产品之一。与其他作物一样,常规育种实践中的高强度选择压力导致亲本遗传基础逐渐狭窄,也是目前烤烟育种面临的突出问题[1-2]。由于亲本材料匮乏,品种间遗传相似度高,导致育成烤烟品种遗传基础狭窄、野生及外来种质渗入较少[3-8]。再加之,对烟草基因组信息的缺乏局限了烤烟资源的有效利用[9-16],进而使拓宽种质资源的选择范围、增强资源的选择效率和提高育种的科学性成为亟需解决的问题。鉴于此,在分子水平上对目前收集的烤烟资源进行亲缘关系研究,将极大地降低遗传背景相似的组合,提高育种的科学性和可行性,对亲本的选择具有重要的指导意义。此外,开发并鉴定出与烤烟农艺性状和化学成分连锁或相关的分子标记,可以为易受环境影响且属微效多基因控制的数量性状开展分子标记辅助选择育种奠定科学基础。
烤烟重要的农艺性状和化学成分几乎全部是受微效多基因控制的数量性状,遗传基础复杂,且极易受环境影响,导致传统的育种方法效率不高。关联分析(association analysis),又称连锁不平衡作图(LD mapping)或关联作图(association mapping),是一种以连锁不平衡为基础,自然群体为材料,检测群体内处于连锁不平衡状态的标记或候选基因的遗传变异与目标性状显著关联频率的方法[17-19]。与QTL定位相比,关联分析不需要专门构建作图群体,自然群体或种质资源即可,同时研究的是多个性状多个位点的关联,不受“两亲本范围”的限制[19]。自Thornsberry等[20]将关联分析首次应用到植物上以来,关联分析作为数量性状研究的有效方法,已广泛应用于大麦[21-22]、玉米[23]、小麦[24-25]、水稻[26]等作物遗传研究中,并获得良好的效果。张吉顺等[27]利用MLM_Q+K和MLM_PCA+K两种最优模型,对258份烤烟种质的13个农艺性状进行了关联分析,检测到18个SRAP标记与6个农艺性状显著关联。余义文等[28]通过关联分析发现了1个SSR标记和6个MFLP标记与烟草TSNA含量显著相关。任民等[29]基于已公布的高密度SSR遗传图谱,寻找获得了24个SSR标记位点与烟草致香物质相关联。樊文强等[30]采用混合线性模型(MLM),进行了标记-性状关联分析,获得11个烟草钾含量的关联SSR标记,并利用其中的5个关联标记对另外130份烟草种质进行扫描,验证该关联标记的稳定性。但利用SSR标记在烤烟自然群体中同时开展农艺性状和化学成分关联分析的研究尚未见报道。
本研究以231份国内外烤烟种质资源为材料,利用SSR标记分析其遗传多样性;在群体遗传结构分析的基础上,寻找和确定与烤烟8个农艺性状和5个化学成分相关联的SSR标记,为在分子水平上对烤烟进行遗传研究并进而开展标记辅助选择育种提供理论依据。
1 材料与方法
1.1 实验材料
231份烤烟种质资源由云南省烟草农业科学研究院提供。包括国内资源116份,含地方资源和育成品种,覆盖全国五大烟叶产区,其中东北烟区11份、黄淮烟区43份、西南烟区37份、华中烟区3份和东南烟区22份;国外资源115份,其中美洲87份、亚洲11份、非洲10份和澳大利亚4份。供试材料的详细信息见附表1(因版面原因不在纸质版刊登,详见电子版)。
1.2 田间试验及性状测定
试验材料种植于云南省烟草农业科学研究院研和试验基地,随机区组设计,2次重复,每份材料种植一个小区,每个小区30株,株行距为0.55m×1.1m,连续种植两年(2012—2013)。按当地优质烟生产技术措施进行栽培管理,每个小区选取15株作为测定样本,农艺性状调查包括株高(包括自然株高和打顶株高)、叶数(包括自然叶片数和有效叶数)、节距、茎围、叶长(腰部叶片)和叶宽(腰部叶片)。各小区采收烘烤后取中部叶(15株测定样本的C1F、C2F、C3F等级的混合烟叶),按烟草行业标准测定烟叶的总糖、还原糖、总氮、烟碱和钾含量。其中,总糖和还原糖按照烟草行业标准YC/T159-2002进行检测,总氮、烟碱和钾含量分别依据YC/T161-2002、YC/T160-2002和YC/T217-2007进行检测,农艺性状数据则按《烟草种质资源描述规范和数据标准》[31]进行收集和测定。
1.3 SSR标记分析
基因组DNA提取、纯化及PCR体系配制、扩增、产物的6%非变性聚丙烯酰胺凝胶电泳检测采用Tong等[32]方法进行。参考本实验室前期构建的遗传图谱[15-16],从中选取120个分布于烟草24个连锁群的SSR标记,其中,每条连锁群上均匀选取5个SSR标记。利用挑选出的SSR引物对231份参试烤烟材料进行总DNA的PCR扩增。
1.4 数据分析
利用NTSYS-pc2.10e软件[33]按Similarity程序将Coefficient参数设置为NEI72,计算材料间的遗传距离(Genetic Distance)并结合MEGA 5.1软件[34]中的非加权配对法(UPGMA)程序进行聚类分析并绘制出聚类图。群体遗传结构分析采用Structure 2.3.4软 件(http://pritchardlab.stanford.edu/structure_software/release_versions/v2.3.4/html/structure.html),估计最佳群体数目K,其取值范围在2-15之间,将MCMC(Markov Chain Monte Carlo)开始时的不作数迭代(Length of burn in period)设为10000,然后将不作数迭代后的MCMC设为100000[35]。每个K值重复运行10次,最后计算出每个K值所对应的后验概率值,依据最大似然值的原则选取合适的K值作为最佳群体数目,当K值持续增大时,参照Evanno等[36]的方法计算ΔK选择合适的K值并计算出Q参数。利用SPAGeDi-1.5软件(http://ebe.ulb.ac.be/ebe/Software.html),对基因型数据进行分析并获得个体间亲缘关系Kinship矩阵[37]。采用TASSEL5.0 软件(http://www.maizegenetics.net/#!tassel/c17q9)中提供的一般线性模型(General Linear Model,GLM)和混合线性模型(Mixed Linear Mo-del,MLM)两种模型分别对两个年份及两年的均值数据,结合SSR标记数据和群体结构进行标记-性状关联分析[17]。GLM分析,主要采用GLM_Q模型[18],将群体结构(Q)矩阵作为协变量,进行标记-性状回归分析;MLM分析,主要采用MLM_Q+K模型[19],同时将个体间亲缘系数(K)和群体结构(Q)作为协变量并进行群体结构校正,分析方法选择EMMA(Efficient Mixed-Model Association)[38-39]。
2 结果与分析
2.1 表型性状变异分析
由231份烤烟种质资源的13个性状分析结果可知,国内外烤烟种质资源性状差异明显,变异丰富,可作为进一步关联分析的理想材料(表1)。13个性状的平均变异系数为20.48%,株高、叶数、茎围、叶长、叶宽及总氮的变异系数小于20%,以叶长变异系数最低只有12.21%;节距、总糖、还原糖、烟碱、钾的变异系数高于20%,其中以烟碱的变异系数最高,达到35.73%。说明参试资源的株高、叶数、茎围、叶长、叶宽及总氮等一致性较强,变异范围不大;相较而言,节距、总糖、还原糖、烟碱、钾变异范围更大。13个性状的平均广义遗传率为65.62%,株高、叶数、茎围、节距的广义遗传率在75%以上,其中株高高达85%以上;总糖、还原糖、总氮、钾的广义遗传率低于50%,表明其极易受环境影响。

表1 231份烤烟种质在2012和2013年的表型多样性统计Tab.1 Statistic summary of phenotypic diversities in 231 flue-cured tobacco germplasms in 2012 and 2013
Note:PnH:plant natural height; nLN:natural leaf number; SC:stem circumference; IL:internode length; LL:length of the largest leaf; WL:width of the largest leaf; TH:topping height; eLN:e ff ective leaf number; TS:total sugar; RS:reducing sugar; TN:total nitrogen; NIC:Nicotine; TP:total potassium.
2.2 遗传距离及聚类分析
120对SSR引物共检测出403个等位基因,变幅为2~21个,平均每个标记3.36个。多态性信息含量(PIC)的分布范围为0.036~0.785,平均值为0.408。通过120个SSR标记基因型分析,231份烤烟资源间的遗传距离(Genetic Distance,GD)变异范围为0.003-0.371(附表2,因版面原因不在纸质版刊登,详见电子版),平均值为0.148。其中,庆胜2号(F059)与武功(F070)的GD最小(GD=0.003),表明二者间亲缘关系最近,其次是Q1214_CSIRO 3T(G057)与Q1232_Virginia Gold(G058),GD为0.004。K326(C001)与歪把子(F067)间GD最大,为0.371,说明二者亲缘关系最远,其次是K149(C007)与TI245(G075),GD为0.368。根据非加权配对法(UPGMA)法进行聚类分析(图1)表明,利用SSR标记可将231份烤烟资源相互区分开,在GD值为0.153水平上聚为4大类。云烟2号(F082)、云烟3号(F083)、Coker176(G012)、CV87(F099)等烤烟育种中主要亲本聚在类别Ⅰ(暗青色);Hicks(G028)、Kutsaga E1(G035)、Nordel(G050)等部分国外引进烤烟品种和小黄金1025(F075)、许金一号(F076)、中烟14(F087)等部分国内自育品种聚于类别Ⅱ(红色);云烟87(B001)、云烟97(B002)、K326(C001)、云烟85(B007)、中烟100(B003)等国内主栽烤烟品种聚在类别Ⅲ(粉红色);红花大金元(B005)、581(A004)、翠碧1号(B12)、阿乔密目(F001)、安丘满屋香(F002)等系选品种及地方资源(品种)聚在类别Ⅳ(黄褐色)。

图1 231份烤烟品种SSR标记遗传距离(GD)聚类图Fig.1 A clustering dendrogram based on the genetic distance of SSR markers in 231 flue-cured tobacco varieties
2.3 群体遗传结构分析
烤烟群体遗传结构分析发现,Ln(P(D))值随假定亚群数K值的增大而持续增大(图2-A),参照Evanno等[36]的方法,最终可通过ΔK来确定该群体划分的亚群数目(即K值)。经过计算,在K=3时,ΔK同时出现最大值和拐点(图2-B),故此参试的自然群体应划分为3个亚群。依据已获得的K值绘制231份烤烟品种群体结构图(图3)。分析各烤烟资源对应的Q(第i材料其基因组变异源于第K群体的概率)值(附表3,因版面原因不在纸质版刊登,详见电子版),结果表明231份烤烟品种中有189份材料(占81.82%)在某一亚群中的Q值大于0.85,说明这些烤烟品种遗传组分相对单一,可以明确地划分到3个亚群内;仅有3份材料(F036:辽烟14号、F095:9417和G063:RG12)在任一亚群内Q值均小于0.5而无法明确其归类。最终这3个亚群分别包含117、38和76份材料,表明该烤烟自然群体结构相对较简单且清晰。
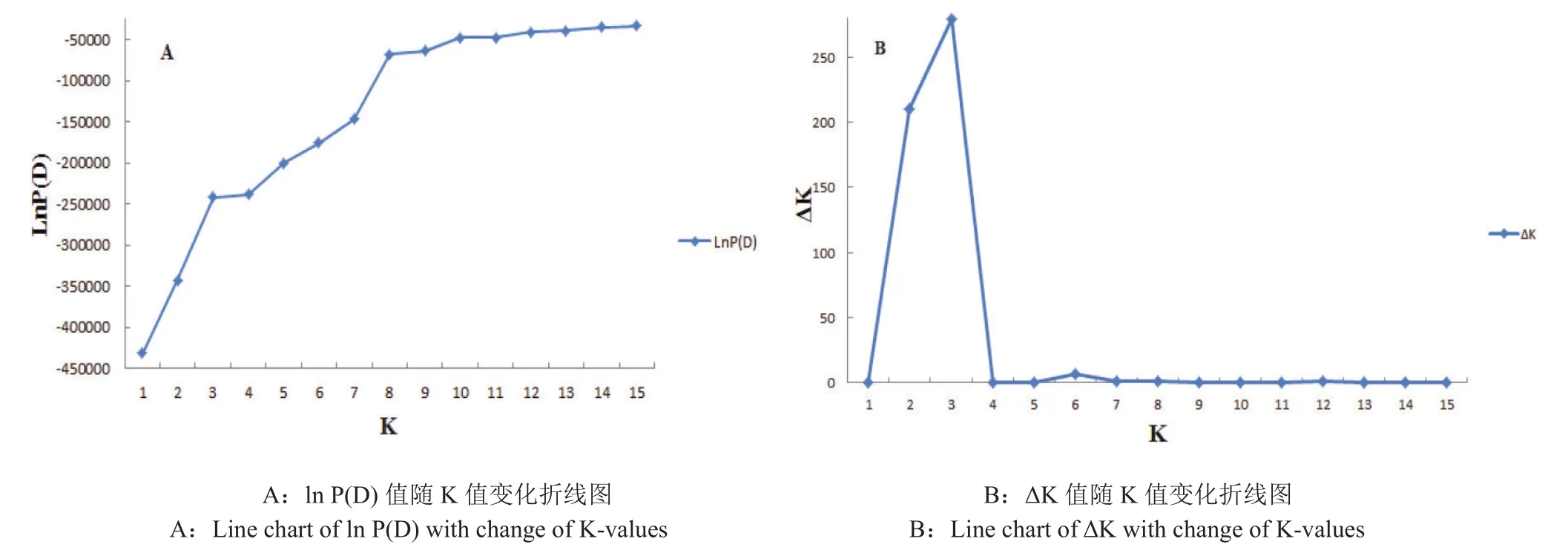
图2 K值与lnP(D)、ΔK值折线图Fig.2 Line chart of K with lnP(D)and ΔK
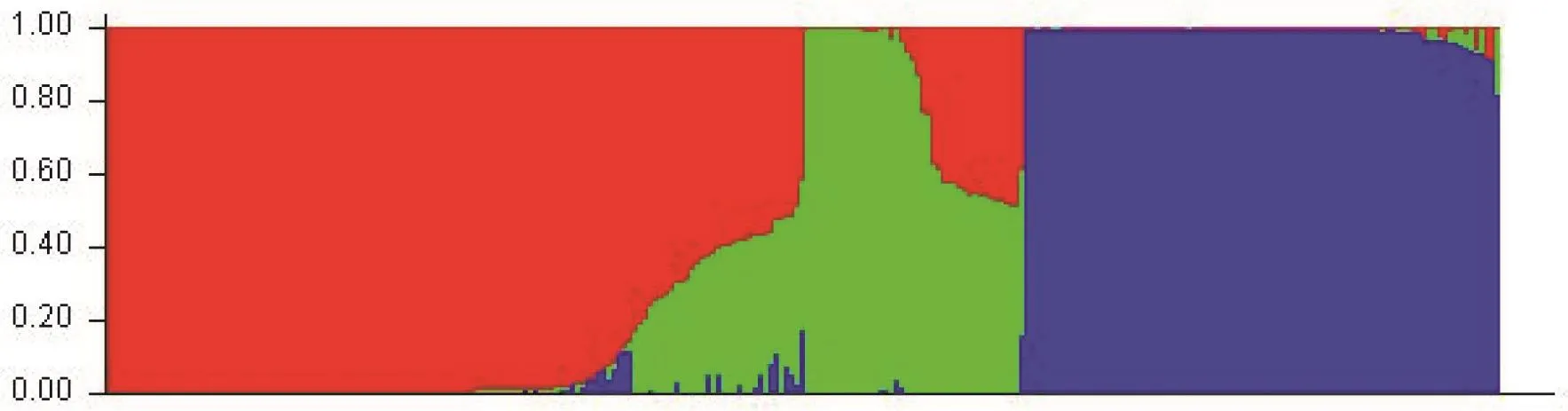
图3 基于SSR标记的231份烤烟品种群体遗传结构Fig.3 Population structure of 231 flue-cured tobacco cultivars accessed by SSR markers
2.4 SSR标记与部分表型性状的关联分析
在GLM_Q和MLM_Q+K模型中,分别以各烤烟材料的对应Q值和Q+K值作为协变量,将SSR标记与8个农艺性状和5个化学成分值共13个表型值进行关联分析,寻找与之相关联的标记,并确定其解释率。GLM_Q分析结果显示(表2),在所检测的120个标记当中,两个年份(2012与2013)共有17个标记与株高、叶数、节距、茎围、叶长、叶宽、总糖、还原糖和总氮11个数量性状在P<0.001水平上显著关联。其中,有13个标记与8个农艺性状显著关联,有4个标记与3个化学成分显著关联,各标记对此11个表型变异解释率为5.03%-22.49%。与株高相关联的SSR标记有4个,与叶数相关的有3个,与节距相关的有3个,与茎围、叶长,叶宽、总糖、还原糖和总氮相关的各有2个。标记TM10481(LG04)、PT54964(LG13)、TM20580(LG17)和PT30200(LG18)与株高相关;TM11179(LG05)、TM10846(LG07)和PT60728(LG22)与叶数相关;PT1279(LG02)和TM10876(LG07)与总糖、还原糖相关。其中,位于第13号(LG13)、第17号(LG17)连锁群上的标记PT54964和TM20580同时与株高和节距显著相关,TM11215(LG17)则与叶长、叶宽显著相关。上述显著相关标记对性状变异的解释率在5.03%与21.33%之间,解释率最大的标记是TM20580(LG17),为21.33%,其次是TM10481(LG04),解释率为19.59%;而解释率最小(5.03%)的是标记TM11179。MLM_Q+K分析结果显示(表2),共有9个标记分别与8个农艺性状和3个化学成分在P<0.001水平上显著相关,比GLM_Q分析结果少8个,即,少的8个标记只在GLM_Q分析中检测到,而MLM_Q+K分析未能检测到新的关联标记。各标记解释率变幅为5.67%-22.49%,且该9个标记与GLM_Q分析结果完全一致,解释率也基本一致。

表2 与性状极显著相关(P<0.001)的SSR标记及其对表型变异的解释率Tab.2 the SSRmarkers associated with traits and their explained phenotypic variations(P<0.001)
3 讨论
3.1 烤烟材料的遗传多样性及群体结构分析
亲本材料的选择是育种工作的基础,由于过度依赖少数骨干亲本材料导致现有的烤烟品种遗传背景高度相似,难以培育出符合生产应用的突破性烤烟品种[1-2,11,40-42]。因此在拓宽烤烟种质资源的基础上分析其遗传多样性,比较资源间遗传背景的相似度,对烟草育种工作具有重要的指导意义。利用SSR分子标记技术对烟草进行遗传多样性的研究,国内外已有报道[7-8,43]。杨柳等[44]利用从102对SSR标记中筛选出的14对引物,对25份普通烟草材料进行遗传多样性分析,结果表明,该25份烟草材料的遗传相似性系数(GS)的范围为0.701~0.959,平均值为0.787,其中17份烤烟种内平均GS则高达0.821,说明烤烟间的遗传相似性较高、遗传基础极其狭窄。Moon等[12]利用分布于烟草24条连锁群上的70对SSR引物对702份收集自全球的烟草种质资源(TIs:Tobacco Introductions)进行了遗传多样性分析,其等位基因数(Na)为2-41个,平均为14.7,检测出的等位基因数目较多;基因多样性变化范围(He)为0.123 ~0.949,平均0.736,基因的多样性水平较高;GS的变幅为0.0007~0.9345,以上结果表明702份TIs烟草种质资源内存在极其丰富的遗传多样性。然而,当利用71对SSR标记对117份美国育成的烤烟品种中的294个等位基因在70年的时间跨度内(1940—2000)进行追踪、分析时,结果表明,可检测的等位基因数目由1940年的294个急剧减少至2000年的13个,即,美国烤烟品种间的遗传多样性极低、遗传基础十分狭窄、品种的趋同性较严重[13]。Fricano等[45]利用分布于烟草7条连锁群上的49个SSR标记对源自世界各地的312份烟草种质资源(TIs)进行了遗传多样性分析,其结果与Moon等[12]利用702份TIs材料的研究结果相反:Na范围为3~13个,平均为6.84,等位基因数目相对较少;He变化范围为0.013~0.841,均值为0.59,基因的多样性水平较低;标记的多态性信息含量(PIC)范围为0.013-0.849,均值为0.431,低于0.5的多态性信息含量值,表明材料间的亲缘关系较近、遗传背景相似。上述类似的问题也存在于本研究所用的231份烤烟材料中,120个SSR位点共检测到403个等位变异,每个位点变异范围为2~21,平均为3.36,等位变异数目较少。多态性信息值PIC的分布范围为0.036~0.785,均值为0.408,多态性不高。遗传距离(GD)的变幅为0.003-0.371,平均值为0.148,遗传距离较小。此外,在GD=0.273时,231份烤烟材料聚为一类,说明供试的烤烟材料亲缘关系较近,遗传基础极其狭窄,需进一步加强外来烟草种质的引进和利用以拓展亲本的遗传多样性。
对种质资源(或自然群体)进行遗传多样性及群体遗传结构分析,是关联作图的前提[46-48]。基于连锁不平衡作图(LDmapping)法并结合主坐标分析(PCoA)和贝叶斯聚类分析,Fricano等[45]将312份TIs烟草材料分别划分为7大类和7个亚群。聚类分析中的7类分别对应白肋烟、黑烟草(Dark)、雪茄烟、香料烟、烤烟、原始烟草(Primitive)和其他(Other),但各大类间仍包含一定量的其他类别烟草材料;群体遗传结构划分中的亚群数目理论上为4个,因雪茄烟、黑烟草、原始烟草和其他4个亚群间无明显的界限应合并为一个亚群,白肋烟、烤烟和香料烟3个类型则各自为一个亚群。分类方法依据的不同及烟草类型间的杂交选育是导致上述两种分类结果不一致的主要原因。在本研究中,通过遗传距离(GD)将231份烤烟材料在GD=0.153处划分为4个大类,其分别包括47、108、21和55份材料。而群体遗传结构分析则将231份材料分为3个亚群,分别包含117、38和76份材料。两者在分类结果上具有较大的差异,造成这种差异的主要原因是两者分类的依据不同:聚类分析依据的是烤烟材料间的遗传距离(GD),反映的是烤烟品种间亲缘关系的远近;而群体遗传结构分类的依据服从Hardy-Weinberg平衡的亚群数目,基于数学模型之上,并计算各供试材料间相应的Q值。因此,本试验中利用Structure 2.3.4软件将供试烤烟材料分为3个群体,具有较高的准确性,避免了人为因素对群体划分的影响[49]。
3.2 烤烟SSR标记的关联分析
相较于 QTL定位,关联分析具有两个突出优点:1)分析时不需要构建群体和遗传图谱,极大的节省了工作量;2)可一次性的从一定规模(数量)的种质资源(材料)中发现(挖掘)优异基因,为遗传育种提供依据。本研究利用TASSEL5.0软件对120个均匀分布于烟草24个连锁群的SSR标记与13个烤烟表型值进行关联分析,以GLM_Q和MLM_Q+K两种模型分别获得17个和9个与株高、叶数、节距、茎围、叶长、叶宽、总糖、还原糖和总氮11个数量性状在P<0.001水平上显著相关的标记,并且MLM_Q+K分析检测到的9个标记均在GLM_Q分析中检测到,且标记解释率大致相当;而另外8个标记并未在MLM_Q+K分析中检测到,其原因是MLM_Q+K分析中同时考虑了群体结构(Q值)和亲缘关系(K值)对关联分析的影响,因此MLM_Q+K分析获得的关联标记数目较MLM_Q少,但结果更加准确。此外,GLM_Q和MLM_Q+K两种模型分析均未检测到与烟碱、钾相关的标记,可能是用于检测的标记数量较少且在烟草基因组中的覆盖度偏低及供试烤烟材料不能提供足够丰富的遗传变异造成的。
本研究选用两年的农艺性状和化学成分表型变异值,通过GLM_Q和MLM_Q+K两种关联分析方法检测出3个与株高相关的SSR标记,分别为TM10481(LG04)、PT54964(LG13)和TM20580(LG17)。且该3个标记与Tong等[50]基于烤烟DH群体对株高QTL定位的结果完全吻合,表明这3个株高相关标记具有很高的稳定性和可靠性。与叶数相关的QTL已被定位在 LG03、LG04、LG06、LG07、LG08、LG14、LG17、LG19和LG22共9个连锁群上,本研究通过关联分析检测出与叶数相关的标记TM10846(LG07)和PT60728(LG22)与Tong等[50]研究获得的标记结果一致。同样,通过GLM_Q和MLM_Q+K两种关联分析方法对两年的表型变异值进行分析,检测出与茎围、叶长相关的标记TM25276(LG08)和TM11215(LG17)也与Tong等[50]对茎围和叶长的QTL定位结果相吻合,只是关联获得的标记数目显著少于QTL定位的结果。节距相关QTL已被定位到连锁群LG05、LG11、LG14、LG17和LG19上,其中LG17连锁群上QTL标记为PT61337和TM20575d[50],本研究通过相关分析检测到的节距相关标记TM20580(LG17)同样位于这一区间,另外1个标记PT54964(LG13)的准确性有待进一步验证。目前已报道5个控制叶宽的QTL,其分别位于LG04、LG05、LG07、LG14和LG22连锁群上[50-52],明显少于其它农艺性状。本研究仅检测出1个与叶宽相关的标记TM11215(LG17),与前人研究结果差异较大,需要验证是否为伪关联,还是存在新的QTL。迄今,有关烟叶化学成分QTL的分析研究非常有限,且仅有的QTL分析仍局限在基于部分烟草遗传连锁图和非SSR标记的条件下进行的[51,53-54]。因此,本研究中通过关联分析获得的与总糖、还原糖和总氮3个化学成分相关的2个标记TM10876(LG7,该标记同时与总糖和还原糖相关)和PT30173(LG17),目前还没有与其相关的文献报道,该结果的真实性需要进一步验证。
4 结论
利用均匀分布于烟草24个连锁群上的120个SSR标记分析了231份烤烟自然群体材料的遗传多样性及群体遗传结构,并通过GLM_Q和MLM_Q+K两种关联分析模型,分别检测出17个和9个与株高、叶数、节距、茎围、叶长、叶宽、总糖、还原糖和总氮11个数量性状在p<0.001水平上极显著相关的标记。9个标记在两个年份和两种模型中均被检测到与同一性状相关,其中3个株高相关标记(TM10481、PT54964和TM20580)、2个叶数相关标记(TM10846和PT60728)、1个茎围相关标记(TM25276)和1个叶长标记(TM11215)与已报道的QTL定位结果一致,可应用于烟草分子标记辅助选择育种。
[1]王元英,周健.中美主要烟草品种亲源分析与烟草育种[J].中国烟草学报,1995,2(3):11-22.WANG Yuanying,ZHOU Jian.Parentage analysis of major tobacco varieties and tobacco breeding in American and China[J].Acta Tabacaria Sinica,1995,2(3):11-22.
[2]常爱霞,贾兴华,冯全福,等.我国主要烤烟品种的亲源系谱分析及育种工作建议[J].中国烟草科学,2013,34(1):1-6.CHANG Aixia,JIA Xinghua.,Feng Quanfu,et al.Parentage analysis of Chinese flue-cured tobacco varieties and breeding suggestion[J].Chinese Tobacco Science,2013,34(1):1-6.
[3]杜传印,刘洪祥,田纪春.部分烟草种质亲缘关系的AFLP分析[J].作物学报,2006,32(10):1592-1596.DU Chuanyin,LIU Hongxiang,TIAN Jichun.Phylogenetic analysis of partial tobacco germplasm resources with AFLP markers[J].ACTA AGRONOMICA SINICA,2006,32(10):1592-1596.
[4]杨友才,周清明,尹晗琪,等.烟草种质资源遗传多样性及亲缘关系的AFLP分析[J].中国农业科学,2006,39(11):2194-2199.YANG Youcai,ZHOU Qingming,YIN Hanqi,et al.Studies on genetic diversity and relationship in tobacco germplasms by AFLP analysis[J].Scientia Agricultura Sinica,2006,39(11):2194-2199.
[5]梁景霞,祁建民,方平平,等.烟草种质资源遗传多样性与亲缘关系的ISSR聚类分析[J].中国农业科学,2008,41(1):286-294.LIANG Jingxia,QI Jianmin,FANG Pingping,et al.Genetic diversity and genetic relatives analysis of tobacco germplasm based on Inter-simple sequence repeat(ISSR)[J].Scientia Agricultura Sinica,2008,41(1):286-294.
[6]李绪英,肖炳光,高玉龙,等.烟草叶绿体基因组和线粒体基因组SSR位点分析[J].西北植物学报,2011,31(12):2399-2405.LI Xuying,XIAO Bingguang,GAO Yulong,et al.Analysis of SSR loci in chloroplast genome and mitochondria genome of tobacco(Nicotiana)[J].Acta Bot Boreal.–Occident Sin,2011,31(12):2399-2405.
[7]张雪廷,童治军,焦芳婵,等.38份晾晒烟种质资源遗传关系的SSR分析[J].植物遗传资源学报,2013,14(4):653-658.ZHANG Xueting,TONG Zhijun,JIAO Fangchan,et al.Genetic relationship analysis of thirty-eight Sun/Air-Cured tobacco germplasms based on Simple Sequence Repeat(SSR)markers[J].Journal of Plant Genetic Resources,2013,14(4):653-658.
[8]李岩,童治军,焦芳婵,等.基于SSR标记对33份烟草材料的聚类分析[J].西北植物学报,2013,33(10):1972-1980.LI Yan,TONG Zhijun,JIAO Fangchan,et al.Cluster analysis of thirty-three tobacco germplasms based on SSR markers[J].Acta Bot Boreal.–Occident Sin,2013,33(10):1972-1980.
[9]Bindler G,Van der Hoeven R,Gunduz I,et al.A microsatellite marker based linkage map of tobacco[J].Theor Appl Genet,2007,114:341-349.
[10]Bindler G,Plieske J,Bakaher N,et al.A high density genetic map of tobacco(Nicotiana tabacumL.)obtained from large scale microsatellite marker development[J].Theor Appl Genet,2011,123:219-230
[11]Moon H S,Nicholson J S,Lewis R S.Use of transferableNicotiana tabacumL.microsatellite markers for investigating genetic diversity in the genusNicotiana[J].Genome,2008,51:547-559.
[12]Moon H S,Nicholson J S,Heineman A,et al.Changes in Genetics Diversity of U.S.Flue-Cured Tobacco Germplasm over Seven Decades of Cultivar Development[J].Crop Science,2009,49:498-506.
[13]Moon H S,Nifong J M,Nicholson J S,et al.Microsatellite-based Analysis of Tobacco(Nicotiana tabacumL.)Genetic Resources[J].Crop Science,2009,49:2149-2159.
[14]Jessada D,Sornsuda S,Wilasinee S,et al.Determination of Local Tobacco Cultivars Using ISSR Molecular Marker[J].Chiang Mai J.Sci.,2010,37(2):293-303.
[15]Tong Z J,Yang Z M,Chen X J,et al.Large-scale development of microsatellite markers inNicotiana tabacumand construction of a genetic map of flue-cured tobacco[J].Plant Breeding,2012,131:674-680.
[16]Tong Z J,Xiao B G,Jiao F C,et al.Large-scale development of SSR markers in tobacco and construction of a linkage map in fluecured tobacco[J].Breeding Science,2016,66:381-390.
[17]Bradbury P J,Zhang Z,Kroon D E,et al.TASSEL:Software for association mapping of complex traits in diverse samples[J].Bioinformatics,2007,23:2633-2635.
[18]Prichard J K,Stephens M,Rosenberg N A,et al.Association mapping in structured populations[J].American Journal of Human Genetics,2000,67:170-181.
[19]Zhang Z,Ersoz E,Lai C Q,et al.Mixed linear model approach adapted for genome-wide association studies[J].Nature Genetics,2010,42:355-360.
[20]Thornsberry J M,Goodman M M,Doebley J,et al.Dwarf8 polymorphisms associate with variation in fl owering time[J].Nat.Genet.,2001,28(3):286-289.
[21]Ivandic V,Hackett C A,Nevo E,et al.Analysis of simple sequence repeats(SSRs)in wild barley from the fertile crescent:associations with ecology,geography and flowering time[J].Plant Mol.Biol.,2002,48:511-527.
[22]Ivandic V,Thomas W T B,Nevo E,et al.Associations of simple sequence repeats with quantitative trait variation including biotic and abiotic stress tolerance inHordeum spontaneum[J].Plant Breeding,2003,122(4):300-304.
[23]Flint-Garcia S A,Thuillet A C,Yu J M,et al.Maize association population:a high resolution platform for quantitative trait locus dissection[J].Plant J.,2005,44:1054-1064.
[24]Breseghello F,Sorrells M E.Association mapping of kernel size and milling quality in wheat(Triticum aestivumL.)cultivars[J].Genetics,2006,172(2):1165-1177.
[25]Roy J K,Bandopadhyay R,Rustgi S,et al.Association analysis of agronomical important traits using SSR,SAMPL and AFLP markers in bread wheat[J].Curr.Sci.,2006,90(5):683-689.
[26]Agrama H A,Eizenga G C,Yan W.Association mapping of yield and its components in rice cultivars[J].Mol.Breeding,2007,19(4):341-356.
[27]张吉顺,王仁刚,杨春元,等.国内外烤烟品种农艺性状的遗传多样性及与SRAP标记的关联分析[J].作物学报,2012,38(6):1029-1041.ZHANG J S,WANG R G,YANG C Y,et al.Genetic diversity of agronomic traits and association analysis with SRAP markers in Flue-Cured tobacco(Nicotiana tabacum)varieties from China and Abroad[J].ACTA AGRONOMICA SINICA,2012,38(6):1029-1041.
[28]余义文,夏岩石,李荣华,等.烟草种质材料TSNA含量的关联分析[J].中国烟草学报,2014,20(3):48-55.YU Y W,XIA Y S,LI R H,et al.Association analysis of tobacco specific nitrosamines content in tobacco germplasm[J].Acta Tabacaria Sinica,2014,20(3):48-55.
[29]任民,张长静,蒋彩虹,等.基于高密度SSR连锁群的烟草致香物质关联分析[J].中国烟草学报,2014,20(4):88-93.REN M,ZHANG C J,JIANG C H,et al.Association analysis of tobacco aroma constituents based on high density SSR linkage group[J].Acta Tabacaria Sinica,2014,20(4):88-93.
[30]樊文强,孙鑫,杨爱国,等.基于关联分析发掘烟草高钾优异等位变异[J].中国烟草学报,2016,doi:10.16472/j.chinatobacco.2015:327.FAN W Q,SUN X,YANG A G,et al.Exploring high-potassium favorable allele mutation of tobacco based on genome-wide association analysis[J].Acta Tabacaria Sinica,2016,doi:10.16472/j.chinatobacco.2015:327.
[31]王志德,王元英,牟建民.烟草种质资源描述规范和数据标准[M].北京,中国农业出版社,2006:1-91.WANG Z D,WANG Y Y,MOU J M.Descriptors and Data Standard for Tobacco Germplasm[M].Beijing:China Agriculture Press,2006:1-91 .
[32]Tong Z J,Jiao T L,Wang F Q,et al.Mapping of quantitative trait loci conferring resistance to brown spot in flue-cured tobacco(Nicotiana tabacumL.)[J].Plant Breeding,2012,131:335-339.
[33]Rohlf F J.Numerical Taxonomy and Multivariate Analysis System NTSYS-pc version 2.1 User Guide[M].Exeter Software Setauket,New York,2000.
[34]Koichiro T,Daniel P,Nicholas P,et al.MEGA5:Molecular Evolutionary Genetics Analysis Using Maximum Likelihood,Evolutionary Distance,and Maximum Parsimony Method[J].Molecular Biology and Evolution,2011,28:2731-2739.
[35]Pritchard J K.,Stephens M,Donnelly P.Inference of population structure using multilocus genotype data[J].Genetics,2000,155:945-959.
[36]Evanno G,Regnaut S,Goudet J.Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study[J].Mol.Ecol.,2005,14:2611-2620.
[37]Hardy O J,Vekemans X.SPAGeDi:a versatile computer program to analyse spatial genetic structure at the individual or population levels[J].Molecular Ecology Notes,2002,2:618-620.
[38]Kang H M.Efficient control of population structure in model organism association mapping[J].Genetics,2008,178:1709-1723.
[39]Zhang Z,Buckler E S,Casstevens T M,et al.Software engineering the mixed model for genome-wide association studies on large samples[J].Brief Bioinform,2009,10:664-675.
[40]杨志晓,王轶,刘红峰,等.我国主栽烤烟品种亲缘关系及育种[J].中国烟草学报,2013,19(2):34-41.YANG Z X,WANG Y,LIU H F,et al.Genetic relationship in major flue-cured tobacco cultivars in China and its implication in variety breeding[J].Acta Tabacaria Sinica,2013,19(2):34-41.
[41]Soheila G,Reza D,Babak A M,et al.Molecular Characterization and Similarity Relationships among Flue-Cured Tobacco(Nicotiana tabacumL.)Genotypes Using Simple Sequence Repeat Markers[J].Not Bot Horti Agrobo,2012,40(2):247-253.
[42]Šarcˇevic´H,Gunjacˇa J,Budimir A,et al.Long-term Genetic Improvement and Genetic Diversity of Croatian Flue-cured Tobacco(Nicotiana tabacumL.)Cultivars[J]。Crop Sci.,2013,53:112-120.
[43]尹国英,杨小燕,何其波,等.鉴定烟草种质资源SSR核心引物筛选和验证[J].植物遗传资源学报,2013,14(5):960-965.YIN G Y,YANG X Y,HE Q B,et al.Screen and Identi fi cation of SSR Core Primers for Tobacco Germplasm[J].Journal of Plant Genetic Resources,2013,14(5):960-965.
[44]杨柳,汪斌,童治军,等.25 份普通烟草种质资源遗传多样性的SSR 标记分析[J].福建农林大学学报( 自然科学版),2013,42(2):171-175.YANG L,WANG B,TONG Z J,et al.Genetic diversity analysis of 25 Nicotiana tabacum L.germplasm resources based on SSR marker[J].Journal of Fujian Agriculture and Forestry University(Natural ScienceEdition),2013,42(2):171-175.
[45]Fricano A,Bakaher N,Del Corvo M,et al.Molecular diversity,population structure,and linkage disequilibrium in a worldwide collection of tobacco(Nicotiana tabacumL.)germplasm[J].BMC Genetics,2012,13:18.
[46]Harris B P,Stokesbury K D E.The spatial structure of local surficial sediment characteristics on Georges Bank,USA[J].Continental Shelf Research,2010,30(17):1840-1853.
[47]Wang M L,Zhu C S,Barkley N A,et al.Genetic diversity and population structure analysis of accessions in the US historic sweet sorghum collection[J].Theoretical and Applied Genetics,2009,120:13-23.
[48]Kline J B,Moore D J,Clevenger C V.Activation and association of the Tec tyrosine kinase with the human prolactin receptor:Mapping of a Tec/Vav-receptor binding site[J].Molecular Endocrinology,2001,15:832-841.
[49]Gupta P K,Rustgi S,Kulwal P L.Linkage disequilibrium and association studies in higher plants:Present status and future prospects[J].Plant Molecular Biology,2005,57:461-485.
[50]TONG Z J,JIAO F C,WU X F,et al.Mapping of Quantitative Trait Loci Underlying Six Agronomic Traits in Flue-Cured Tobacco(Nicotiana tabacumL.)[J].ACTA AGRONOMICA SINICA,2012,38(8):1407-1415.
[51]Julio E,Denoyes-Rothan B,Verrier J L,et al.Detection of QTLs linked to leaf and smoke properties inNicotiana tabacumbased on a study of 114 recombinant inbred lines[J].Mol.Breeding,2006,18(1):69-91.
[52]Lewis R S,Milla S R,Kernodle SP.Analysis of an introgressed Nicotiana tomentosa genomic region aVecting leaf number and correlated traits inNicotiana tabacum[J].Theor Appl Genet,2007,114:841-854.
[53]Zhou L Y,Li R Y,Fan L J,et al.Mapping epistasis and environment×QTX interaction based on four -omics genotypes for the detected QTX loci controlling complex traits in tobacco[J].The Crop Journal,2013,6:151-159.
[54]肖炳光,卢秀萍,焦芳婵,等.烤烟几种化学成分的QTL初步分析[J].作物学报,2008,34(10):1762-1769.XIAO B G,LU X P,JIAO F C,et al.Preliminary QTL analysis of several chemical components in flue-cured tobacco(Nicotiana tabacumL.)[J].ACTA AGRONOMICA SINICA,2008,34(10):1762-1769.
SSR marker-based analyses on genetic diversity and relevant variations of agronomic traits and chemical composition of 231 flue-cured tobacco germplasm resources
TONG Zhijun,CHEN Xuejun,FANG Dunhuang,ZENG Jianmin,WU Xingfu,XIAO Bingguang*
Yunnan Academy of Tobacco Agricultural Sciences/Key Laboratory of Tobacco Biotechnological Breeding/National Tobacco Genetic Engineering Research Center,Kunming 650021
The aim of this study was to investigate genetic diversity of flue-cured tobacco(Nicotiana tabacum L.)germplasm and to screen molecular markers for agronomic and chemical traits.A total of 231 flue-cured tobacco germplasms were genotyped with 120 simple sequence repeats(SSRs)markers uniformly distributed on 24 linkage groups.Associations of SSR variation with eight agronomic traits and fi ve chemical components were investigated by two contrasting statistical models,i.e.general linear model(GLM)and mixed linear model(MLM).Overall,403 alleles were identi fi ed in all accessions,ranging from 2 to 21 per locus,with an average of 3.36 alleles per locus.Mean polymorphic information content was 0.408,ranging from 0.036 to 0.785.Genetic distance between varieties ranged from 0.003 to 0.371,with a mean of 0.148.Using unweighted pair-group method with arithmetic mean(UPGMA),all accessions were clustered into four groups at a genetic distance of 0.153.Using population genetic structure analysis,the test materials were divided into three subpopulations.Association analysis by GLM_Q and MLM_Q+K models revealed that seventeen and nine markers were signi fi cantly associated with a total of 13 phenotypic traits,respectively.Moreover,nine markers were consistently associated with the same traits at the p < 0.001 level in two years using both models.Speci fi cally,three markers associated with plant height(TM10481,PT54964,and TM20580),two markers with leaf number(TM10846 and PT60728),one marker with stem circumference(TM25276),and one marker with leaf length(TM11215)supported published quantitative trait locus(QTL)mapping results.These seven markers could be used in future molecular marker-assisted selection to improve breeding efficiency.
flue-cured tobacco;genetic diversity;association analysis;agronomic traits;chemical composition

童治军,陈学军,方敦煌,等.231份烤烟种质资源SSR标记遗传多样性及其与农艺性状和化学成分的关联分析[J].中国烟草学报,2017,23(5)
中国烟草总公司项目(110201601027(JY-01),110201603008);中国烟草总公司云南省公司项目(2016YN23,2017YN01,2017YN05)
童治军(1980—),博士,助理研究员,主要从事烟草分子育种研究,Email:tzj861@163.com
肖炳光,博士,研究员,Email:xiaobg@263.net
2017-07-18;< class="emphasis_bold">网络出版日期:
日期:2017-10-19
:TONG Zhijun,CHEN Xuejun,FANG Dunhuang,et al.SSR marker-based analyses on genetic diversity and relevant variations of agronomic traits and chemical composition of 231 flue-cured tobacco germplasm resources[J].Acta Tabacaria Sinica,2017,23(5)
*Corresponding author.Email:xiaobg@263.net
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
今日农业(2020年17期)2020-12-15
湖北农机化(2020年4期)2020-07-24
四川蚕业(2020年4期)2020-02-10
活力(2019年19期)2020-01-06
中国烟草学报(2019年5期)2019-11-14
中国麻业科学(2018年6期)2018-04-09
浙江中西医结合杂志(2017年2期)2017-01-12
声屏世界(2014年6期)2014-02-28
