
小兴安岭原始阔叶红松林与枫桦次生林土壤呼吸及其各组分特征的比较研究
2017-11-09 13:37耿兆鹏毛子军韩营营
植物研究 2017年2期
耿兆鹏 毛子军 黄 唯 韩营营
(东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040)
小兴安岭原始阔叶红松林与枫桦次生林土壤呼吸及其各组分特征的比较研究
耿兆鹏 毛子军*黄 唯 韩营营
(东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040)
原始阔叶红松林是我国温带典型的地带性顶极植被类型,枫桦次生林是其典型的次生林类型之一,对二者土壤呼吸及其各组分特征的研究有助于准确评价该地区的碳平衡。本研究主要测定了2013和2014年2个生长季原始阔叶红松林和枫桦次生林土壤呼吸(RS),并量化了土壤呼吸的各个组分(异养呼吸RH和自养呼吸RA),与此同时测量了土壤10 cm处温度以及土壤含水率。研究结果表明,土壤呼吸及其各组分有着明显的季节变化特性,其大小的变化主要受温度的影响,土壤10 cm处的温度可以解释RS64%~70%、RH56%~65%、RA77%~79%的变异。对于温度的敏感性,原始阔叶红松林土壤呼吸Q10值>枫桦次生林土壤呼吸Q10值,而在单一林型中的比较,RAQ10值>RSQ10值>RHQ10值。此外,总体Q10值随着季节有着明显的变化,且随着温度的升高有降低的趋势。原始阔叶红松林和枫桦次生林RS年平均速率分别为3.92和4.06 μmol·m-2·s-1,RH年平均速率分别为2.97和2.85 μmol·m-2·s-1,RA年平均速率则分别为0.96和1.17 μmol·m-2·s-1。原始阔叶红松林土壤呼吸以及土壤土壤自养呼吸要稍低于枫桦次生林,而原始阔叶红松林异养呼吸则高于枫桦次生林异养呼吸,但差异不显著。原始阔叶红松林和枫桦次生林RS平均年通量分别为942和971 g C·m-2·a-1,RH年通量分别为709和677 g C·m-2·a-1,RA年通量则分别为215和276 g C·m-2·a-1。原始阔叶红松林RS年通量略高于枫桦次生林RS年通量,但差异不显著。我们的实验结果表明,小兴安岭地区枫桦次生林正向演替的过程中, 植被演替变化对土壤呼吸及各组分的影响并不明显,相较于环境因子温度和湿度要小的多。
演替;阔叶红松林;枫桦次生林;土壤呼吸;自养呼吸;异养呼吸
土壤呼吸是陆地生态系统碳素循环的主要环节,也是CO2浓度升高的关键生态学过程[1]。面对潜在的气候变化,土壤呼吸相对微小的改变也能影响生态系统碳源/碳汇的状态[2],显著改变大气中CO2的浓度和土壤碳的累积速率,从而加剧或减缓全球气候变暖[3],因此探究土壤呼吸的控制因素以及量化其对于气候变化的响应已迫在眉睫。森林演替是改变地球生物化学循环的一种基本的生态学过程[4],它对于土壤呼吸的影响是目前生态学研究的热点问题之一。但以往的研究中有关森林演替不同阶段森林土壤呼吸的比较研究鲜见报道[5~7]。尤其是原始阔叶红松林被砍伐后,在进行天然更新的过程中所形成的次生林生态系统是否改变了原来的土壤呼吸特征,进而影响生态系统的碳循环是目前尚未深入探讨的科学问题,有待于进行深入的研究。
在我国东北森林生态系统中,典型的地带性植被阔叶红松林的面积已经非常有限,取而代之的是各种类型的次生群落,枫桦天然次生林是东北阔叶红松林次生演替中期的典型群落类型之一。本研究的目的在于通过对小兴安岭地区阔叶红松林与其被砍伐后所形成的枫桦次生林土壤呼吸作用的比较研究,揭示次生演替群落土壤呼吸及其各组分与原始阔叶红松林的差异及其作用机理,从而为准确评估小兴安岭地区乃至全国森林生态系统碳平衡提供科学参考。
1 研究区域概况及样地选择
研究样地位于小兴安岭南坡人为干扰较少的达里岭支脉东坡的凉水国家级自然保护区。地理位置为东经128°47′8″~128°57′19″,北纬47°6′49″~47°16′10″,总面积为12 133 hm。该地区有明显的温带大陆性季风气候的特征,春秋两季气候多变,春季降水少,常有大风,秋季降温急剧,会出现早霜现象。夏季降水集中,其中6月到8月的降水占到全年的60%以上,冬季气候严寒,多风霜且干旱。无霜期100~120 d,积雪期130~150 d。土壤类型为暗棕壤[8]。在该区域内选择生境相似的原始阔叶红松林和枫桦次生林作为实验林分,在两个林分中分别选择三块面积为20 m×20 m的实验样地,采用常规方法对样地进行调查。其中,两个林分的胸高断面积分别为68.52、26.46 m2·hm-2,比值为2.59∶1,取0~60 cm土样测定土壤有机碳密度。主要树种组成及样地概况见表1,样地土壤有机碳密度参数见表2。

表1 样地概况
注:1.红松;2.五角槭;3.臭冷杉;4.青楷槭;5.糠椴;6.枫桦;7.白桦;8.山杨;9.大青杨;10.暴马丁香;11.鱼鳞云杉;12.榛子
Note:1.Pinuskoraiensis; 2.Acermono; 3.Abiesnephrolepis; 4.Acertegmentosum; 5.Tiliamandshurica; 6.Betulacostata; 7.Betulaplatyphylla; 8.Populusdavidiana; 9.Populusussuriensis; 10.Syringaamurensis; 11.Piceajezoensis; 12.Corylusheterophylla
表2样地0~60cm土层土壤有机碳密度
Table2Organicmattercontentparametersofthesampleplotssoil(kg·m-2)
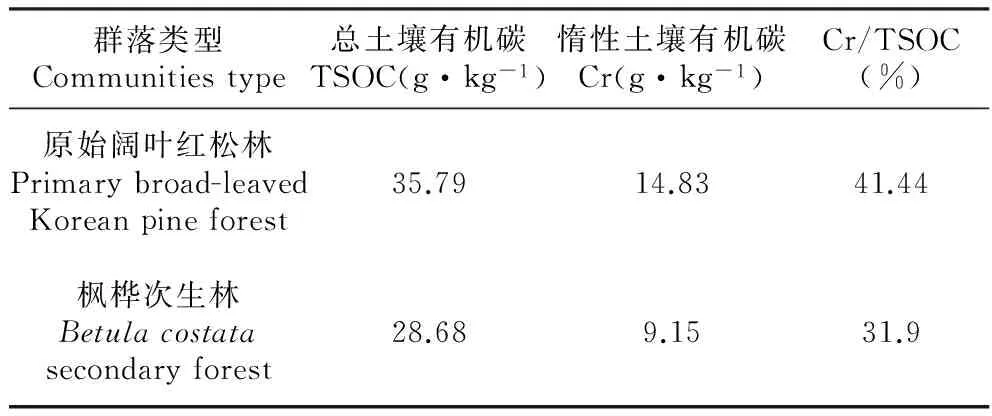
群落类型Communitiestype总土壤有机碳TSOC(g·kg-1)惰性土壤有机碳Cr(g·kg-1)Cr/TSOC(%)原始阔叶红松林Primarybroad-leavedKoreanpineforest35.7914.8341.44枫桦次生林Betulacostatasecondaryforest28.689.1531.9
注:土壤有机碳基础数据来源于本研究组前期研究测定[9]。
Note:Soil organic carbon based data derived from prior research of this team.
2 实验方法
2.1 实验设计
2013年4月底,采用随机区组设计,在阔叶红松林和枫桦次生林的三个固定样地(20 m×20 m)内随机选择3个1 m×1 m的小样方,在周围挖壕至根系分布层以下,用400目的尼龙网纱将小样方围起来用以隔离小样方周围的根系,以防止根系进入小样方,再小心的去除小样方内所有的活体植物,尽量保持土壤原状表层。在每个小样方内按照品字形安置3个内径为10.4 cm,高度为8 cm的PVC土壤环,将PVC环插入土壤的一端削尖,以减少对土壤的压力,土壤环露出地面2 cm,保持土壤环在整个试验周期中不被扰动,然后再在壕沟周围随机安置8个相同的PVC环。在壕沟内的根系在经过近两个月的分解之后进行土壤呼吸的测定。在2013年6~10月和2014年5~10月,每个月测定一次数据,每次测定基本都在9~16时,共测定11次。在每次测定前,仔细检查土壤环,以确保土壤环与土壤的完全接触,且定期清除壕沟内地表植被,尽量保持壕沟内没有活体植物[10]。在整个试验周期,测定土壤呼吸以及瞬时温度和湿度,进行分析。所有这些测定基本涵盖了本样地除冬季以外(冬季封山,无法测定)的土壤呼吸值与土壤温度、土壤湿度的季节动态变化范围。
2.2土壤呼吸、温度、湿度的测定及其Q10的计算
本研究壕沟外土壤呼吸(RS)的测定利用LI-6400同化室连接到LI-6400便携式CO2/H2O测定系统进行测定。在试验中测定一个循环,取所有值,然后算得平均值,作为每次测定的土壤呼吸值(RS)。土壤自养呼吸(RA)和土壤异养呼吸(RH)的测定利用壕沟法间接测定,即壕沟内测定的土壤呼吸为RA,则:
RH=RS-RA
(1)
本研究采用LI-6400同化室连接到LI-6400便携式CO2/H2O测定系统自带的土壤温度探针测定距土壤表面10cm深处的瞬时土壤温度,并与土壤呼吸值的测定保持同步,测定3次,求平均值。同时利用TDR300(Spectrum Technologies,USA)探针测定0~10 cm深度土壤的平均体积含水量。土壤温度、土壤湿度的动态监测数据由安装在样地内的HOBO小型气象站提供。
土壤呼吸以及各组分的呼吸速率与温度之间的关系需要在测得的数据基础之上,利用软件对土壤呼吸采用指数模型模拟[11]。
RS=ROeβT
(2)
式中:RS表示平均土壤呼吸速率,T表示平均土壤温度,RO表示0℃时的呼吸速率,β表示温度反应系数。土壤温度敏感性指数(Q10值)是指温度升高10℃时土壤呼吸速率变化的倍数[12~13]:
Q10=e10β
(3)
式中:β为温度反应系数,由此可以得出Q10值。
2.3 数据处理分析
数据分析采用了Microsoft Excel 2010和SPSS 19统计分析软件,利用Excel 2010和Origin 8来辅助作图。
3 结果和分析
3.1原始阔叶红松林与枫桦次生林土壤总呼吸及其各组分的季节动态及其比较
2013年6月~2014年10月原始阔叶红松林和枫桦次生林的土壤呼吸的测定结果显示,两种林型间差异不显著,但两种林型RS、RH、RA都具有明显的随季节变化性(图1:a),在不同月份RS差异显著(P<0.05),其呼吸值都表现为先升高后降低的趋势,呈单峰曲线趋势。在5~8月,土壤呼吸值逐渐升高,在8月土壤呼吸值达到了最大。其中,原始阔叶红松林RS、RH、RA速率变化范围依次为1.39~7.08 μmol·m-2·s-1,1.07~5.31 μmol·m-2·s-1,0.31~1.78 μmol·m-2·s-1,而枫桦林次生林RS、RH、RA呼吸速率变化范围依次为1.42~7.02 μmol·m-2·s-1,1.01~5.10 μmol·m-2·s-1,0.33~1.92 μmol·m-2·s-1。在8月之后,两种林型的呼吸值开始降低,由于在冬季仪器受限制,土壤呼吸测定到10月份以后停止(图1)。
3.2原始阔叶红松林与枫桦次生林土壤总呼吸及其各组分的与温度之间的关系
分析结果显示,原始阔叶红松林和枫桦次生林在生长季土壤温度差异不显著(P>0.05),两种林型的土壤总呼吸及其各组分的呼吸值都随着温度的升高而增强,温度与土壤总呼吸及其各组分的呼吸值呈明显的指数关系,RS与温度显著相关(P<0.01)(图2)。
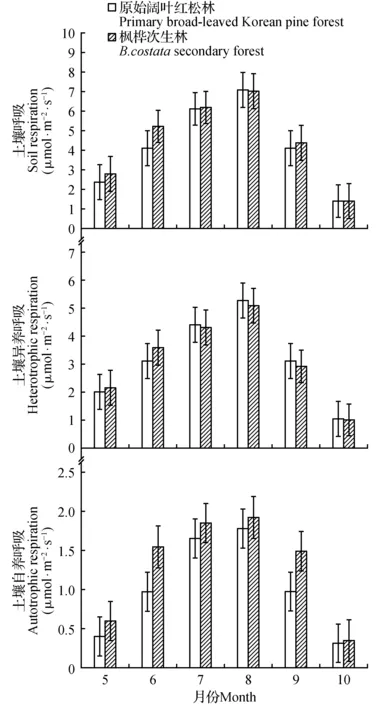
图1 原始阔叶红松林和枫桦次生林土壤呼吸(RS)、土壤异养呼吸(RH)、土壤自养呼吸(RA)季节动态Fig.1 Seasonal variations of soil respiration, heterotrophic respiration and autotrophic respiration at Primary broad-leaved Korean pine forest and B.costata secondary forest
Table3Duncan’smultiple-rangetestforthemeansofsoilrespiration,soiltemperatureandmoistureinPrimarybroad-leavedKoreanpineforestandB.costatasecondaryforest

群落类型Communitiestype土壤呼吸Soilrespiration(μmol·m-2·s-1)土壤温度Soiltemperature(℃)土壤体积含水量Soilvolumetricwatercontent平均值Mean标准差Standarddeviation变异系数Coefficientofvariation平均值Mean标准差Standarddeviation变异系数Coefficientofvariation平均值Mean标准差Standarddeviation变异系数Coefficientofvariation原始阔叶红松林Primarybroad-leavedKoreanpineforest3.921.97509.85.565727.611.224枫桦次生林B.costatasecondaryforest4.061.864610.285.645532.581.75
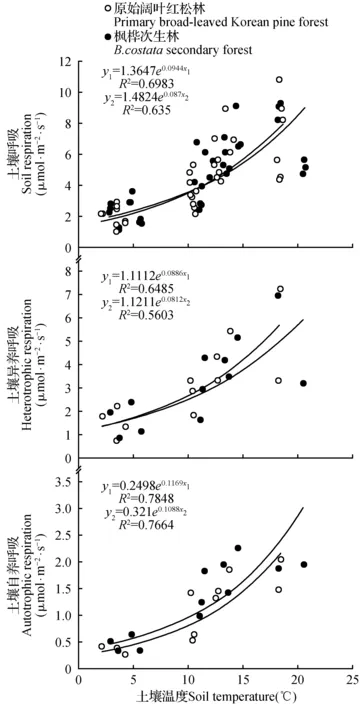
图2 土壤呼吸速率和土壤10 cm温度之间的关系Fig.2 Relationships between soil respiration and soil temperature at the depth of 10 cm
群落类型Communitiestype土壤呼吸RS异养呼吸RH自养呼吸RA原始阔叶红松林Primarybroad-leavedKoreanpineforest2.572.433.22枫桦次生林B.costatasecondaryforest2.392.252.97
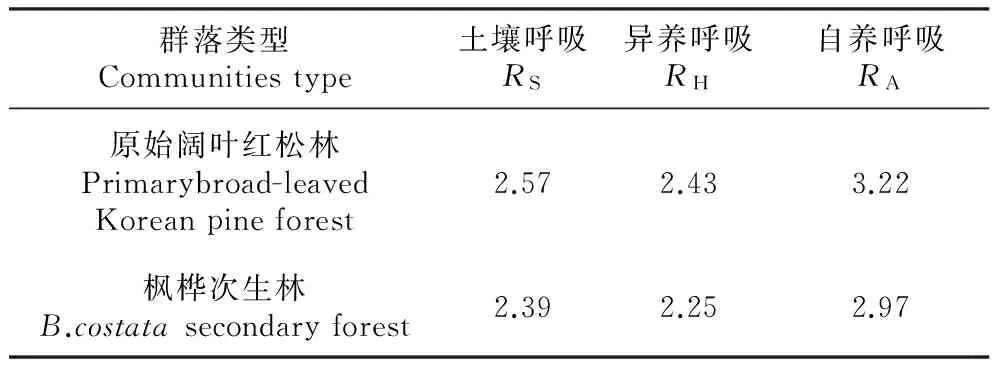
利用公式(2)分析土壤呼吸及其各组分对温度的敏感性发现,原始阔叶红松林RS、RH、RA的Q10值均大于枫桦次生林RS、RH、RA的Q10值(表4),表明原始阔叶红松林土壤呼吸及其各组分对于温度的敏感性要高于枫桦次生林,经斜率差异显著性性检验表明两种林型土壤呼吸及其各组分对于温度的敏感性差异性不显著(P>0.05)。而就单一林型而言,原始阔叶红松林和枫桦次生林土壤呼吸及其各组分Q10值的规律一致,即RAQ10值>RSQ10值>RHQ10值,表明土壤自养呼吸对于温度的反应最为敏感。
为了探究整体Q10值随季节、温度的变化,将温度与土壤呼吸值进行拟合。最先计算5月份总的Q10值,接下来计算5、6月份总的Q10值,以此类推最后计算所有月份总的Q10值。结果显示Q10值随着季节有着明显的变化,且随着温度的升高有降低的趋势,在8月份温度达到最高时,Q10值则处在最小的水平。在整个生长季Q10值随温度的变化趋势与土壤呼吸值随温度变化的趋势截然相反,其曲线呈一条“V”型曲线,在温度最高时达到最低,而温度较低时,土壤呼吸的温度敏感性则较高,土壤呼吸的温度敏感性与温度负相关(图3)。
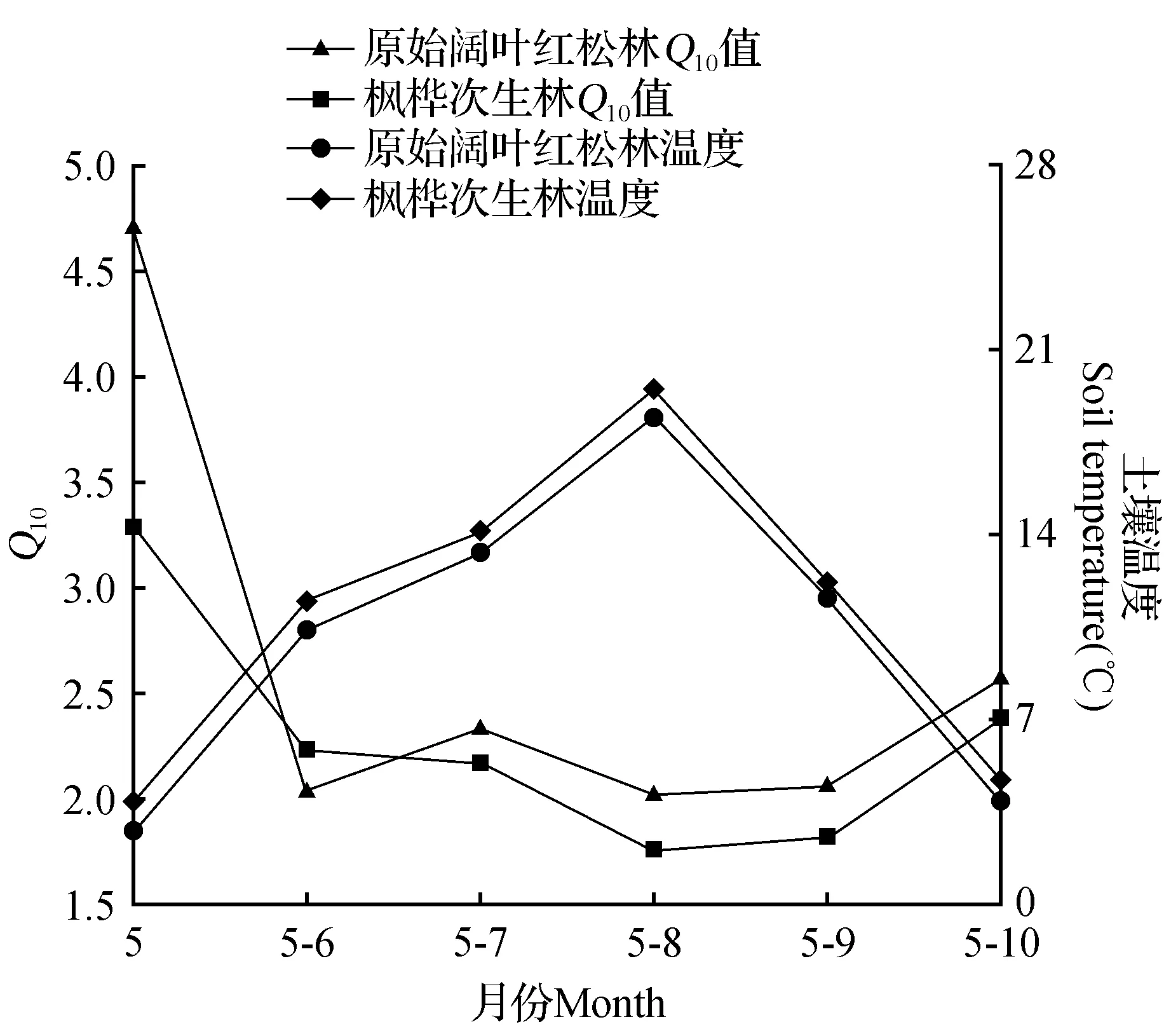
图3 土壤呼吸总体Q10值随季节、温度的变化Fig.3 The overall Q10 value of soil respiration varies when the seasons and the temperature changes
3.3原始阔叶红松林与枫桦次生林土壤总呼吸及其各组分与湿度之间的关系
原始阔叶红松林的土壤含水率总体上低于枫桦次生林,且差异显著(P<0.05),但各月份之间相差不大,而枫桦次生林土壤含水率的在各个月之间差异较大(图4),相关分析结果显示土壤呼吸与土壤含水量之间的相关性不显著(P>0.05),表明土壤湿度并不是影响该地区原始阔叶红松林和枫桦次生林土壤呼吸的关键因子。

图4 原始阔叶红松林和枫桦次生林土壤10 cm处温度和土壤体积含水量的季节性变化特征Fig.4 Seasonal dynamics of soil temperature at the depth of 10 cm and soil volumetric water content in the Primary broad-leaved Korean pine forest and B.costata secondary forest
3.4原始阔叶红松林和枫桦次生林土壤呼吸各组分贡献率及其动态变化
土壤呼吸分别由土壤自养呼吸与土壤异养呼吸共同组成。总体上,2个林型的异养呼吸明显大于自养呼吸,但不同的土壤呼吸组分所占的比重也不尽相同。其中,原始阔叶红松林的土壤呼吸中,异养呼吸所占的比例平均约为77%,而自养呼吸所占的比例仅为23%。枫桦次生林的异养呼吸所占的比例约为71%,而自养呼吸所占的比例则占到了29%(图5),经过检验表明原始阔叶红松林和枫桦次生林之间异养呼吸和自养呼吸所占比例差异显著(P<0.05)。

图5 原始阔叶红松林(a)和枫桦次生林(b)各组分对土壤总呼吸贡献率动态Fig.5 Dynamics of contribution rate of soil respiration components to total soil respiration in the Primary broad-leaved Korean pine forest(a) and B.costata secondary forest(b)
生长季土壤呼吸各组分的贡献在呈动态变化,自养呼吸比例的变化趋势是逐渐升高然后下降,异养呼吸所占比例则逐渐下降然后再升高,自养呼吸最大比例及异养呼吸最小比例出现在7月份。
3.5原始阔叶红松林与枫桦次生林土壤呼吸年通量
通过观测,原始阔叶红松林和枫桦次生林内部小气候环境和土壤水热条件有着些许差异,因此将观测所得两种林型10 cm处土壤在生长季的日温,结合公式(1)算出两种林型每天向大气中释放的CO2的量,然后依次累加,据此可以估算出在生长季2个林分土壤累积向大气中释放的CO2的量(表5),即原始阔叶红松林和枫桦次生林土壤累积向大气释放CO2的量分别为754和777 g C·m-2·a-1。
由于研究地区小兴安岭冬季封山,且LI-6400便携式CO2/H2O测定系统在低温条件下工作困难。所以在11月至来年4月的土壤表面CO2通量的值无法测定,因此需要对此期间的阔叶红松林与枫桦林的土壤表面CO2通量进行估算。根据已有的研究经验,暂且假定生长季RS为全年的RS的80%[14]。通过计算得出原始阔叶红松林的年通量约为942 g C·m-2·a-1,而枫桦次生林的年通量则大约为971 g C·m-2·a-1。由此可以看出,枫桦次生林土壤产生的CO2年通量略大于原始阔叶红松林的CO2年通量,但统计分析表明其差异不显著。

表5 原始阔叶红松林与枫桦次生林各组分土壤呼吸及其年通量
注:字母不同表示同一项目在不同林型之间差异显著(P<0.05)。
Note:Different letters indicate significance(P<0.05)。
4 讨论
4.1原始阔叶红松林与枫桦次生林土壤呼吸的季节动态及其主要环境驱动因子
在本研究中,原始阔叶红松林与枫桦次生林随季节的变化规律与大部分的温带森林生态系统研究得出的结论基本一致[15~17],即两种林型的土壤呼吸作用在夏季8月份最大,而在生长季初期5月份和生长季末期10月份土壤呼吸作用则偏小。此变化趋势正好与温度的变化相一致,相关性分析也显示二者极显著相关(P<0.001)。表明了温度是影响原始阔叶红松林和枫桦次生林土壤呼吸的关键因子。这是由于一定范围内环境温度升高可增强微生物活性,加速土壤中有机质的分解,从而增加土壤中CO2浓度;同时,土壤温度直接影响植物生长和生理活动,从而影响根系呼吸[18~19],导致土壤呼吸速率随着温度的升高而增强。
4.2原始阔叶红松林与枫桦次生林土壤呼吸的温度敏感性
土壤呼吸对环境因子变化的潜在响应可以细化为自养呼吸和异养呼吸对于环境因子的潜在响应,因为二者对土壤温度的响应可能有所不同[20]。通常情况下,土壤呼吸的温度敏感性用Q10来表示,即温度每增加10℃土壤呼吸所增加的倍数,Q10值有着很大的变率,因此不同群落以及同一群落土壤呼吸的不同组分对于温度的敏感性指数Q10可能不尽相同。在本研究中,原始阔叶红松林的土壤呼吸Q10值高于枫桦次生林,这可能与2个林分呼吸底物的多少有关,本研究的2个群落类型的土壤分析显示(表2):阔叶红松林的土壤有机碳含量为35.79 g·kg-2,其中惰性土壤有机碳密度为14.83 g·kg-2,占总有机碳的41.44%,而枫桦次生林的土壤有机碳密度为28.68 g·kg-2,其中惰性土壤有机碳含量为9.15 g·kg-2,占总有机碳的31.9%。由于土壤生物、底物质量和底物供应显著调控土壤呼吸的Q10值,土壤呼吸过程中产生的CO2主要来自于呼吸底物的分解,因此底物质量和底物供应会显著影响土壤呼吸及其温度敏感性[21]。依据热力学原理,分子结构越复杂的底物,即越难以分解的有机物,具有的活化能就越高,对温度的敏感性也越大[22~23]。2个群落土壤呼吸各组分Q10值表现为RA>RS>RH,表明2种群落土壤自养呼吸的温度敏感性要高于异养呼吸的温度敏感性,这与陆彬[23]、韩天丰[8]、Gaumont-Guay[4]以及Ruehr[25]等的结论一致。阔叶红松林自养呼吸的Q10值是异养呼吸Q10值的1.34倍,而枫桦次生林为1.32倍,而原始阔叶红松林自养呼吸Q10值是枫桦次生林自养呼吸Q10值的1.08倍。Boone[26]等的研究发现根呼吸的Q10值是4.6,而土壤微生物呼吸的Q10值仅为2.5,他认为根呼吸如此大的Q10值是根生物量的季节性变化和现存根量对温度变化共同响应的结果。本研究群落阔叶红松林<5 cm细根现存量(5.48 t·hm-2)是枫桦次生林现存量(3.97 t·hm-2)的1.38倍,表明Q10值可能与根系生物量的影响有关。
4.3原始阔叶红松林与枫桦次生林土壤呼吸各组分的贡献率及年通量
在本研究中,土壤呼吸各组分对于土壤呼吸的贡献率为RH>RA。表明在整个土壤呼吸的进行中,RH占据了主导地位。其中原始阔叶红松林的异养呼吸贡献率最大,达到了87%,而枫桦次生林的异养呼吸贡献率则为71%。可能的原因是原始阔叶红松的森林组成与结构更为复杂,经过多年的累积,土壤有机质含量较高,导致微生物组成更为丰富多样、数量更多,从而使异养呼吸所占的比重增加。但枫桦次生林自养呼吸的贡献率要高于原始阔叶红松林的贡献率,表明不同群落类型自养呼吸与异养呼吸存在差异。
在本研究中原始阔叶红松林土壤表面CO2年通量处在831~1 053 g C·m-2·a-1,枫桦次生林土壤表面CO2年通量处在915~1 027 g C·m-2·a-1,此研究结果与杨金艳[27]、韩营营[18]在温带森林做出的结果相近。枫桦次生林土壤表面CO2年通量略高于原始阔叶红松林土壤表面CO2年通量,但统计分析二者差异不显著。有关森林演替过程中土壤碳通量的研究结果很不一致,王旭的结果表明,长白山原始阔叶红松林土壤呼吸大于杨桦林,Wang Jin[28]等对美国弗吉尼亚北部不同演替阶段各种植被类型,包括2个森林群落的土壤CO2年通量的研究则发现,不同演替阶段各群落间土壤年CO2通量没有变化,只是季节动态上有差异,而温度则是最主要的影响因子。我们认同Wang Jin等的观点,本研究中,整个生长季枫桦次生林的土壤温度高于原始阔叶红松林0.48℃,这或许是枫桦次生林土壤碳通量略高于阔叶红松林的原因。
5 结论
通过对小兴安岭地区原始阔叶红松林和枫桦次生林近两个生长季的实验测定结果表明,原始阔叶红松林和枫桦次生林土壤呼吸及其各组分的月动态皆呈现较为一致的单峰曲线,且在温度最高的8月份达到顶峰。土壤温度是影响该地区两种林型土壤呼吸的关键因子,二者显著相关(P<0.01),土壤水分对于两个林型土壤呼吸的影响则不显著(P>0.05)。两种林型的年平均土壤呼吸速率以及年通量差异皆不显著,枫桦次生林要稍大于原始阔叶红松林,但是原始阔叶红松林和枫桦次生林之间自养呼吸以及异养呼吸所占的比例差异显著。此外,在本研究中原始阔叶红松林土壤呼吸及其各组分的Q10值大于枫桦次生林土壤呼吸及其各组分的Q10值,而对于单一林型土壤呼吸及其各组分的Q10值大小关系则为:土壤自养呼吸Q10值>土壤呼吸Q10值>土壤异养呼吸Q10值,表明阔叶红松林土壤呼吸对温度的敏感性高于枫桦次生林。因此,在全球变化的大环境下,相较于其他次生林,作为我国温带典型地带性顶极植被类型得原始阔叶红松林更容易作出响应,从而发生改变。本实验结果表明,小兴安岭地区枫桦次生林正向演替的过程中植被演替变化对土壤呼吸及各组分的影响并不明显,相较于环境因子温度和湿度要小的多。然而,小兴安岭地区原始阔叶红松林和枫桦次生林的土壤呼吸作用受各方生长因子及其周边环境的影响会有所不同,虽然总体呈现出来的土壤呼吸差异不明显,但是在自养呼吸以及异养呼吸所占的比例方面差异明显。因此,在评估小兴安岭地区乃至全国森林生态系统碳平衡,特别是在研究自养呼吸或者异养呼吸时应该进行区别对待,将红松林处在不同演替阶段以及形成的不同林型加以区别,这样才能得到更加准确的数据,从而为全球变化的进一步研究作出相应的贡献。
1.张宪权,王文杰,祖元刚,等.东北地区几种不同林分土壤呼吸组分的差异性[J].东北林业大学学报,2005,33(2):46-47,73.
Zhang X Q,Wang W J,Zu Y G,Zhang Wan li.The Difference between Different Components of Soil Respiration in Several Types of Forests in Northeastern China[J].Journal of Northeast Forestry University.2005,33(2):46-47,73.
2.Wagle P,Kakani V G.Confounding effects of soil moisture on the relationship between ecosystem respiration and soil temperature in switchgrass[J].BioEnergy Research,2014,7(3):789-798.
3.Schlesinger W H,Andrew J A.Soil respiration and the global carbon cycle[J].Biogeochemistry,2000,48(1):7-20.
4.De Kovel C G F,Van Mierlo A E M,Wilms Y J O,et al.Carbon and nitrogen in soil and vegetation at sites differing in successional age[J].Plant Ecology,2000,149(1):43-50.
5.韩天丰,周国逸,李跃林,等.中国南亚热带森林不同演替阶段土壤呼吸的分离量化[J].植物生态学报,2011,35(9):946-954.
Han T F,Zhou G Y,Li Y L,et al.Partitioning soil respiration in lower subtropical forests at different successional stages in southern China[J].Chinese Journal of Plant Ecology,2011,35(9):946-954.
6.Yan J H,Zhang D Q,Zhou G Y,et al.Soil respiration associated with forest succession in subtropical forests in Dinghushan Biosphere Reserve[J].Soil Biology and Biochemistry,2009,41(5):991-999.
7.王旭,周广胜,蒋延玲,等.山杨白桦混交次生林与原始阔叶红松林土壤呼吸作用比较[J].植物生态学报,2007,31(3):348-354.
Wang X,Zhou G S,Jiang Y L,et al.soil respiration in natural mixed(BetulaplatyphyllaandPopu-lusdavidiana) secondary forest and primary broad-leaved korean pine forest[J].Journal of Plant Ecology,2007,31(3):348-354.
8.陈金玲,金光泽,赵凤霞.小兴安岭典型阔叶红松林不同演替阶段凋落物分解及养分变化[J].应用生态学报,2010,21(9):2209-2216.
Chen J L,Jin G Z,Zhao F X.Litter decomposition and nutrient dynamics at different succession stages of typical mixed broad leaved-Korean pine forest in Xiaoxing’ an Mountains,China[J].Chinese Journal of Applied Ecology,2010,21(9):2209-2216.
9.丁一阳,毛子军,张玲,等.小兴安岭原始阔叶红松林和枫桦次生林土壤有机碳库比较研究[J].植物研究,2015,35(4):604-611.
Ding Y Y,Mao Z J,Zhang L,et al.Comparison of Soil Organic Carbon Stock ofBetulacostataSecondary Forest and Mixed Broadleaved Korean Pine Original Forest in Xiaoxing’an Mountains[J].Bulletin of Botanical Research,2015,35(4):604-611.
10.杨金艳,王传宽.东北东部森林生态系统土壤碳贮量和碳通量[J].生态学报,2005,25(11):2875-2882.
Yang J Y,Wang C K.Soil carbon storage and flux of temperate forest ecosystems in northeastern China[J].Acta Ecologica Sinica,2005,25(11):2875-2882.
11.Luo Y Q,Wang S Q,Hui D F,et al.Acclimatization of soil respiration to warming in a tall grass prairie[J].Nature,2001,413(6856):622-625.
12.Davidson E A,Belk E,Boone R D.Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest[J].Global Change Biology,1998,4(2):217-227.
13.Rey A,Pegoraro E,Tedeschi V,et al.Annual variation in soil respiration and its components in a coppice oak forest in central Italy[J].Global Change Biology,2002,8(9):851-866.
14.Bond-lamberty B,Wang C K,Gower S T.A global relationship between the heterotrophic and autotrophic components of soil respiration[J].Global Change Biology,2004,10(10):1756-1766.
15.韩营营,黄唯,孙涛,等.不同林龄白桦天然次生林土壤碳通量和有机碳储量[J].生态学报,2015,35(5):1460-1469.
Han Y Y,Huang W,Sun T,et al.Soil organic carbon stocks and fluxes in different age stands of SecondaryBetulaplatyphyllain Xiaoxing’an Mountain,China[J].2015,35(5):1460-1469.
16.Tang J W,Bolstad P V,Martin J G.Soil carbon fluxes and stocks in a Great Lakes forest chronosequence[J].Global Change Biology,2009,15(1):145-155.
17.Buchmann N.Biotic and abiotic factors controlling soil respiration rates inPiceaabiesstands[J].Soil Biology and Biochemistry,2000,32(11-12):1625-1635.
18.Tang J W,Baldocchi D D,Qi Y,et al.Assessing soil CO2efflux using continuous measurements of CO2profiles in soils with small solid-state sensors[J].Agricultural and Forest Meteorology,2003,118(3-4):207-220.
19.Chen S T,Zou J W,Hu Z H,et al.Global annual soil respiration in relation to climate,soil properties and vegetation characteristics:summary of available data[J].Agricultural and Forest Meteorology,2014,198-199:335-346.
20.陆彬,王淑华,毛子军,等.小兴安岭4种原始红松林群落类型生长季土壤呼吸特征[J].生态学报,2010,30(15):4065-4074.
Lu B,Wang S H,Mao Z J,et al.Soil respiration characteristics of four primary Korean pine communities in growing season at Xiaoxing’an Mountain,China[J].Acta Ecologica Sinica,2010,30(15):4065-4074.
21.杨庆朋,徐明,刘洪升,等.土壤呼吸温度敏感性的影响因素和不确定性[J].生态学报,2011,31(8):2301-2311.
Yang Q P,Xu M,Liu H S,et al.Impact factors and uncertainties of the temperature sensitivity of soil respiration[J].Acta Ecologica Sinica,2011,31(8):2301-2311.
22.Davidson E A,Janssens I A.Temperature sensitivity of soil carbon decomposition and feedbacks to climate change[J].Nature,2006,440(7081):165-173.
23.Bosatta E,Ågren G I.Soil organic matter quality interpreted thermodynamically[J].Soil Biology and Biochemistry,1999,31(13):1889-1891.
24.Gaumont-guay D,Black T A,Barr A G,et al.Biophysical controls on rhizospheric and heterotrophic components of soil respiration in a boreal black spruce stand[J].Tree Physiology,2008,28(2):161-171.
25.Ruehr N K,Buchmann N.Soil respiration fluxes in a temperate mixed forest:seasonality and temperature sensitivities differ among microbial and root-rhizosphere respiration[J].Tree Physiology,2010,30(2):165-176.
26.Boone R D,Nadelhoffer K J,Canary J D,et al.Roots exert a strong influence on the temperature sensitivity of soil respiration[J].Nature,1998,396(6711):570-572.
27.杨金艳,王传宽.东北东部森林生态系统土壤呼吸组分的分离量化[J].生态学报,2006,26(6):1640-1647.
Yang J Y,Wang C K.Partitioning soil respiration of temperate forest ecosystems in Northeastern China[J].Acta Ecologica Sinica .2006,26(6):1640-1647.
28.Wang J,Epstein H E,Wang L X.Soil CO2flux and its controls during secondary succession[J].Journal of Geophysical Research,2010,115(G2):G02005.
This study was fully supported by the National Natural Science Foundation of China(31270494)
introduction:GENG Zhao-Peng(1991—),male,mainly engaged in the research of Forest ecology.
date:2016-05-11
ComparativeStudyontheSoilRespirationandComponentCharacteristicsofPrimaryBroad-leavedKoreanPineForestandBetulacostataSecondaryForestinXiaoxing’anMountatins,China
GENG Zhao-Peng MAO Zi-Jun*HUANG Wei HAN Ying-Ying
(Key Laboratory of Forest Plant Ecology, Ministry of Education,Northeast Forestry University,Harbin 150040)
Primary broad-leaved Korean pine forest is a typical zonal climax vegetation type in the temperate zone of China.Betulacostatasecondary forest is one of its typical secondary forests. The study on soil respiration and the characteristics of the components of the primitive broad-leaved Korean pine forest andB.costatasecondary forest will be helpful in gaining insight into Xiaoxing’an Mountains spatial and temporal distribution characteristics of carbon source and sink as well as its main influence factors. The final end is to accurately evaluate the carbon balance of this region. We tested the soil respiration(RS) of Primary broad-leaved Korean pine forest andB.costatasecondary forest in 2013 and 2014, adopted procedure of quantization in the test of soil respiration components(heterotrophic respirationRH, autotrophic respirationRA), and surveyed the soil temperature and soil moisture at 10 cm of the soil. The soil respiration and its main components feature obvious seasonal variation, which is mainly influenced by temperature. The temperature at the depth of 10 cm of the soil can explain the variation ofRSof 64%-70%,RHof 56%-65%, andRAof 77%-79%. As for the temperature sensitivity, primary broad-leaved Korean pine forest’sQ10value of soil respiration>B.costatasecondary forestQ10value of soil respiration. While in the comparison of a single forest,Q10ofRA>Q10ofRS>Q10ofRH. The generalQ10value presents obvious seasonal changes and the value decreases with the rise in temperature. The average soil respiration ofRSof the primitive broad-leaved Korean pine forest and theB.costatasecondary forest are 3.92 and 4.06 μmol·m-2·s-1, respectively, the average soil respiration ofRHare 2.97 and 2.85 μmol·m-2·s-1, respectively, while the average soil respiration ofRAare 0.96 and 1.17 μmol·m-2·s-1, respectively. The average annual flux ofRSof the primitive broad-leaved Korean pine forest and theB.costatasecondary forest are 942 and 971 m-2·a-1C, respectively; the average annual flux ofRHare 709 and 677 m-2·a-1C, respectively, while the average annual flux ofRAare 215 and 276 m-2·a-1C, respectively. The average annual flux ofRSof Primary broad-leaved Korean pine forest is higher than that ofB.costatasecondary forest, but the differences are not significant. The area of Xiaoxing’an Mountains is in the succession process ofB.costatasecondary forest, vegetation succession influence on soil respiration and its components is not obvious, and compared with environmental factor the temperature and humidity is much smaller.
succession;Primary broad-leaved Korean pine forest;Betula costata secondary forest;soil respiration;heterotrophic respiration;autotrophic respiration
国家自然科学基金(31270494)
耿兆鹏(1991—),男,硕士研究生,从事森林生态学相关研究。
* 通信作者:E-mail:zijunm@nefu.edu.cn
2016-05-11
* Corresponding author:E-mail:zijunm@nefu.edu.cn
S791.247
A
10.7525/j.issn.1673-5102.2017.02.021
猜你喜欢
现代农村科技(2022年5期)2022-11-18
新农业(2022年9期)2022-05-16
小哥白尼(野生动物)(2021年9期)2022-01-17
中国林副特产(2020年5期)2020-10-09
生物工程学报(2020年6期)2020-07-31
现代农业研究(2017年11期)2018-01-12
现代园艺(2017年20期)2017-02-03
农业与技术(2016年6期)2016-05-14
满族文学(2014年5期)2014-09-22
