
华北地区园林树木水分利用特性及影响因子研究
2017-10-25 00:50李少宁鲁绍伟杨新兵
河南农业大学学报 2017年4期
丁 杰,李少宁,鲁绍伟,丁 丽,杨 超,陈 波,杨新兵
(1.河北农业大学林学院,河北 保定 071000;2.北京市农林科学院林业果树研究所,北京 100093;3.北京林果业生态环境功能提升协同创新中心,北京 100093;4.北京燕山森林生态系统定位观测研究站,北京 100093)
华北地区园林树木水分利用特性及影响因子研究
丁 杰1,2,3,李少宁2,3,4,鲁绍伟2,3,4,丁 丽1,杨 超1,2,3,陈 波2,3,4,杨新兵1
(1.河北农业大学林学院,河北 保定 071000;2.北京市农林科学院林业果树研究所,北京 100093;3.北京林果业生态环境功能提升协同创新中心,北京 100093;4.北京燕山森林生态系统定位观测研究站,北京 100093)
以16种园林树木为研究对象,利用盆栽试验方法,采用美国CI-340便携式光合测定系统测定其水分利用特性。结果表明, 1)园林树种于7:00-11:00的WUE明显高于13:00-17:00的WUE,针叶和灌木树种WUE日变化表现出整体下降的趋势,而阔叶和经济林树种均存在“午休”现象。2)经济林树种WUE生长中期最高,为 (4.04±0.20) μmol·mmol-1;阔叶树种WUE为生长季初期最大,为 (3.82±0.56) μmol·mmol-1;灌木树种生长末期最高,为 (2.94±0.05) μmol·mmol-1;针叶树种WUE生长中期最高,为 (2.73±0.25) μmol·mmol-1。3)树种WUE为经济林树种((3.65±0.12) μmol·mmol-1)>阔叶树种((3.33±0.41) μmol·mmol-1)>灌木树种 ((2.90±0.27) μmol·mmol-1)>针叶树种((2.64±0.27) μmol·mmol-1),经济林树种是针叶树种的1.38倍。4)WUE与Pn、Gs呈现出极显著正相关关系(P<0.01),与PAR呈显著性正相关关系(P<0.05),与VPD、t(a)、t(l)呈极显著负相关关系(P<0.01)。研究结果可为水资源的合理利用和园林绿地的科学管理提供理论依据。
园林树种;水分利用率;影响因子
水作为一种重要的自然资源,是促进社会经济发展、推动文明进步的重要因素之一。随着社会经济迅速发展以及城市化建设进程加快,对水资源和环境的需求不断增加,而中国水资源相对匮乏[1-2]。据统计,中国每年因环境污染造成的损失约占GDP的3%~8%[3],为解决环境污染问题,提高生活质量,许多城市开始大范围的建设城市园林绿地,但是园林植物自身巨大的耗水量也不容忽视。有研究发现,平均树高21 m的65种树木中90%的树木单株日耗水量为10~200 kg,加剧了日益增多的城市水资源供需矛盾[4]。因此,园林绿地中选择水分利用率高、抗旱性强的树种并加以科学的管理就显得尤为迫切。研究园林树木的水分利用特性,提高水分利用率具有重要意义。水分利用率(WUE)是树木消耗单位水分所固定同化物质的量,反映了叶片光合过程固定 CO2与蒸腾耗水之间的关系,是评价植物生长适宜程度的综合生理生态指标,也是表征植物抵御干旱胁迫能力的重要依据,一直是国内外干旱、半干旱以及湿润地区农林业、生物学、生态学等多学科研究探讨的热点问题之一[5-11]。赵顺等[12]通过测定不同遮荫处理下臭柏幼苗的光合特性,发现遮荫处理下的WUE和净光合速率(Pn)呈极显著相关性,揭示出臭柏在不同的光照条件通过较高的Pn达到较高的WUE。刘茜等[13]对樟树叶片的蒸腾特征和WUE进行研究发现,叶片WUE与气温(t(a))、叶面温度(t(l))和胞间CO2浓度(Ci)具有较高的相关性。何雅冰等[14]研究了侧柏叶片气孔导度(Gs)对WUE的影响。虽然毛振华等[15]对6种园林地被植物在不同灌溉水平条件下的WUE变化特征进行了比较,但以上研究主要集中在对单个树种或同一种类植物间水分利用特性的研究,对不同种类植物水分利用特性研究较少。研究叶片尺度上的WUE,可以揭示植物内在机制。因此,本试验采用盆栽苗木法,结合手持式CI-340光合测定仪研究叶片层面上不同种类园林树木的水分利用特性及其影响因子。可为园林绿地抗旱树种的筛选和日常管理以及有限水资源的高效利用提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
试验地处北京市农林科学院林业果树研究所的枣种质资源圃内,林果所位于北京市海淀区香山景区,占地40 hm2,西邻北京市西山国家森林公园。该所地理坐标116°13′E,39°59′N,位于华北平原西北部,北温带半湿润大陆性季风气候。太阳辐射量全年平均为112~136 Kcal·cm-1。年均降雨量约483.9 mm,主要集中在夏季6-8月。所内共保存林果种质资源2 400余份,主要有侧柏(Platycladusorientalis)、桧柏(Sabinachinensis)、毛白杨(Populustomentosa)、刺槐(Robiniapseudoacacia)、扶芳藤(Euonymusfortunei)、月季(Rosachinensis)、核桃(Juglansregia)、板栗(Castaneamollissima)、山楂(Crataeguspinnatifida)、桃(Amygdaluspersica)、葡萄(Vitisvinifera)、枣(Ziziphusjujuba)、樱桃(Cerasuspseudocerasus)等。
1.2 试验材料
选取华北地区主要的园林绿化树种(表1)为研究对象,每个树种选择5棵长势相近且生长健壮的苗木放于直径为30 cm高的盆中,取原生土在人工控制条件下盆栽培养,经过2个月的缓苗,对苗木进行蒸腾耗水特性研究。
1.3 研究方法
试验于植物生长季进行,从2016-05-01—2016-10-31,每月选择晴朗无云的天气测定相关数据。试验苗木放置于防雨棚内培养测定,以消除降水的影响。在人工控制条件下(灌足底水后的第2 d),选用手持式仪器CI-340光合测定仪测定树种的蒸腾速率(Tr,mmol·m-2·s-1)、净光合速率(Pn,μmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、胞间CO2质量浓度(Ci,mg·L-1)、水汽压亏缺(VPD,kPa)、叶面温度(t(l),℃)等参数。选取树冠中上部外围向阳的5片完整叶子,每片叶子测定3~5个瞬时值,取平均值,测定时间为7:00-17:00,每隔2 h测定一次。

表1 供试树种基本情况Table 1 Basic information of tested tree species
WUE(水分利用率)=Pn(光合速率)/Tr(蒸腾速率)[16]。
气象因子采用试验地内的米特(Meter)全自动气象站,实时观测并自动记录太阳总辐射(W·m-2)、光合有效辐射(PAR,μmol·m-2·s-1)、气温(t(a),℃)、空气相对湿度(RH,%)、风速(W,m·s-1)、降水(P,mm)等,采集频率为1 h。
1.4 数据处理
试验所用数据为3~5次测定的平均值,相关性分析和画图处理运用SPSS 22.0统计软件与Microsoft Excel 2013。
2 结果与分析
2.1 光合有效辐射、光合速率、蒸腾速率、气孔导度、胞间CO2浓度和水汽压亏缺日动态变化
随着外界条件(光合有效辐射、温度)的规律性变化,Gs、VPD、Ci、Pn、Tr发生改变,水分利用特性同样也具有节律性变化。强烈的太阳辐射是引起t、Tr等日变化发生的重要因素[17]。如图1所示,PAR、VPD、t(a)、t(l)表现为单峰曲线(峰值出现在11:00-13:00),到11:00-13:00上升到最大值,随后依时序呈现出递减趋势。Gs、Pn、Tr为双峰曲线(峰值出现在11:00和15:00),11:00达到最高值后开始下降,到15:00稍有回升后仍呈现出减弱趋势。Ci则显示出双谷趋势,从7:00开始逐渐下降,11:00下降到首个谷值后上升,于13:00又开始降低,到15:00出现第2个谷值,17:00时有所升高。
2.2 不同种类树木水分利用特性
2.2.1 针叶树种水分利用特性 如图2所示,针叶树种盆栽苗木的WUE日动态变化呈现出下降趋势,7:00-9:00WUE达最高值,15:00-17:00最低,早晨WUE整体高于傍晚。这一结果的出现主要是与针叶树种光合作用存在光补偿点和光饱和点有关,蒸腾作用则不存在光限制,因而Pn受限引起变化程度减小,最终导致WUE逐渐减弱。一天中的各个时刻白皮松WUE普遍大于其他树种,WUE日均值为白皮松((3.00±0.62) μmol·mmol-1)>侧柏((2.72±0.78 )μmol·mmol-1)>沙地柏((2.68±0.80) μmol·mmol-1)>桧柏((2.54±0.82) μmol·mmol-1),白皮松抗干旱胁迫能力强于侧柏、沙地柏和桧柏。
针叶苗木的WUE月变化表现出8月明显高于其他月份(图3)。针叶树种中生长时期(7、8月)WUE最高,生长季初期(5月)最低。侧柏WUE月变化为“N”型曲线,8月份WUE达到最大值((2.99±0.65) μmol·mmol-1),9月有所下降,为 (2.51±0.66) μmol·mmol-1,10月份回升为 (2.67±0.69) μmol·mmol-1。桧柏呈现出倒“V”型曲线,即8月WUE到达最大值(2.63±0.87) μmol·mmol-1。白皮松和沙地柏WUE月变化均显示出倒“N”型曲线,白皮松5-7月WUE呈现下降趋势,8月份回升并达到最高((3.16±0.63) μmol·mmol-1)后开始降低。沙地柏WUE月变化为6月份降到最低值 (2.09±0.98) μmol·mmol-1,7月开始上升为 (2.90±0.82) μmol·mmol-1,8月达最高值 (3.05±0.70)μmol·mmol-1,9、10月份呈下降趋势。白皮松5、6月份WUE显著高于其他针叶树种(高28%~29%)。桧柏WUE于7~10月份普遍低于另外3个树种(低11%~17%)。

图1 PAR、Ci、Gs、VPD、T、Tr和Pn日变化Fig.1 The daily changes of PAR、Ci、Gs、VPD、T、Tr and Pn

图2 针叶树种水分利用率日变化Fig.2 Daily changes of coniferous plants’ WUE

图3 针叶树种水分利用率月变化Fig.3 Month changes of coniferous plants’ WUE
2.2.2 阔叶树种水分利用特征 阔叶苗木WUE上午明显高于下午,WUE日变化整体呈降低趋势(图4)。7:00-9:00WUE值最大,13:00出现低谷,15:00有所回升后继续呈现下降趋势,即存在“午休”和“傍晚休”,主要是因为外界环境条件的变化,Pn变化的程度小于Tr变化的程度而引起的WUE改变。除栾树(最高值出现在7:00)外,其他树种最大值出现在9:00,最高值分别为栾树((4.87±1.28) μmol·mmol-1)、银杏((4.33±3.23) μmol·mmol-1)、刺槐((4.30±1.31) μmol·mmol-1)、白蜡((3.76±1.11) μmol·mmol-1)。
由图5可得阔叶树种WUE月变化特征,5、7月份WUE均值高于6、8月份,主要与Tr增加的幅度高于Pn增加的幅度有关。9、10月份最低,10月树种间差异较小,介于(2.30±0.65 ~2.74±0.34) μmol·mmol-1之间,8-10月树种均呈现出下降趋势。阔叶树种在春季和夏季会合成大量的营养物质用于自身生长,因而其消耗单位水分所固定同化物质的量高,WUE相对于秋季(9、10月)大。各个树种WUE月变化特征存在差异,栾树、白蜡表现为倒“N”型特征,7月份出现最大值,分别为(5.23±1.28) μmol·mmol-1、(4.12±0.95) μmol·mmol-1;银杏显示出“M”型特征,最高值出现在7月份((4.26±1.32) μmol·mmol-1);而刺槐则显现出逐渐减弱趋势,即5月份WUE最大((4.19±1.70) μmol·mmol-1),10月份最小((2.30±0.65) μmol·mmol-1)。WUE月均值为栾树((3.83±0.92) μmol·mmol-1)>银杏((3.49±0.50) μmol·mmol-1)>白蜡((3.02±0.61 )μmol·mmol-1)>刺槐((2.99±0.73) μmol·mmol-1),栾树是刺槐的1.28倍,因此栾树具有较强的耐旱能力。

图4 阔叶树种水分利用率日变化Fig.4 Daily changes of borad-leaf plants’ WUE

图5 阔叶树种水分利用率月变化Fig.5 Month changes of borad-leaf plants’ WUE
2.2.3 经济林树种水分利用特征 由图6可知,经济林盆栽苗木WUE日变化规律趋于一致,均表现出“V”型变化,存在水分利用“午休”现象,与中午时分出现的高温高辐射导致的Pn、Tr降低有关,而Pn降低的程度要大于Tr,所以出现了WUE下降的趋势,即出现“午休”现象。7:00WUE最高,随后呈现递减趋势,13:00达到最低值,山楂稍有延迟,在15:00出现最小值,17:00WUE值有所提高。“午休”期间,山楂、核桃的WUE高于樱桃和山杏,值分别为(3.35±0.55) μmol·mmol-1、(3.01±0.51) μmol·mmol-1、(2.85±0.84) μmol·mmol-1、(2.81±0.50) μmol·mmol-1,其中山楂是山杏的1.19倍,说明山楂适应高温高辐射的能力强于其他经济林树种。
经济林树种WUE月变化见图7,各树种间7-9月份WUE波动较大,差值高达34%。山楂、山杏和樱桃月动态呈现倒“N”型趋势,核桃表现为“M”型变化,7-9月份树种WUE相差较大,9、10月份WUE普遍低于其他月份,主要是由于5-8月份果实中生长、积累营养较多。山楂和樱桃均为5月份和8月份WUE较高,山楂WUE值分别为(4.19±1.09)μmol·mmol-1、(4.02±0.59) μmol·mmol-1,樱桃WUE值分别为 (4.21±0.74 )μmol·mmol-1、(5.17±1.12) μmol·mmol-1,山杏为7月份最高((4.38±0.66) μmol·mmol-1),核桃为8月份最高((4.38±0.93) μmol·mmol-1)。经济林树种WUE月均值相差不大,分别为山楂((3.76±0.54) μmol·mmol-1)>樱桃((3.73±0.95) μmol·mmol-1)>核桃((3.59±0.69) μmol·mmol-1)>山杏((3.51±0.74) μmol·mmol-1)。

图6 经济林树种水分利用率日变化Fig.6 Daily changes of economic plants’ WUE
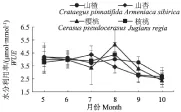
图7 经济林树种水分利用率月变化Fig.7 Month changes of economic plants’ WUE
2.2.4 灌木树种水分利用特征 从图8可以看出,灌木树种苗木WUE日动态曲线表现明显,整体呈现减弱趋势,降幅为33%~38%。7:00-11:00WUE值普遍大于13:00-17:00 的WUE值。从7:00开始,温度T和光合有效辐射PAR持续增强,导致Pn、Tr增强,但灌木树种Pn增加的幅度小于Tr增加的幅度,导致WUE降低,下午时分,T、PAR开始减弱,Pn、Tr也随之下降,而Pn下降的程度要大于Tr下降的程度,所以出现了一天中WUE持续减弱的趋势。黄杨WUE日动态为“W”型曲线,木槿为倒“N”型曲线,柽柳和枸杞为递减曲线,各树种WUE最大值均出现在7:00。木槿仅13:00WUE值低于其他树种,全天的其余时刻均高于黄杨、柽柳、枸杞,说明木槿对高温高辐射的环境条件最敏感。WUE整体显示出木槿((3.15±0.85) μmol·mmol-1)>枸杞((3.06±0.70 )μmol·mmol-1)>黄杨((2.82±0.68) μmol·mmol-1)>柽柳((2.54±0.70) μmol·mmol-1)。
图9为灌木树种WUE月动态变化,灌木树种WUE月波动幅度小,介于(2.36±0.40~3.37±1.20) μmol·mmol-1之间。灌木树种9、10月的WUE相对较其他月份高,与灌木树种需储存较多的营养成分以利于次年的生长发育有关。木槿除10月份以外,其他月份WUE均高于剩余树种,而柽柳则恰恰相反,虽然5-9月均低于另外3个树种,但在10月份有所上升,高出黄杨15%。黄杨WUE月趋势为单峰曲线,峰值 (2.96±0.71) μmol·mmol-1出现在7月份;木槿为双峰曲线,峰值出现在6月份((3.35±0.79) μmol·mmol-1)和9月份((3.37±1.20)μmol·mmol-1);柽柳和枸杞为“N”型曲线,最大值在10月份出现,分别为(2.83±1.03) μmol·mmol-1、(3.27±0.75) μmol·mmol-1。木槿、枸杞抵御干旱胁迫的能力强于黄杨和柽柳。

图8 灌木树种水分利用率日变化Fig.8 Daily changes of shrubs plants’ WUE

图9 灌木树种水分利用率月变化Fig.9 Month changes of shrubs plants’ WUE
综上所述,园林树种在7:00-11:00的WUE普遍高于13:00-17:00的WUE,上午消耗单位水分所固定同化物质的量最大。针叶树种、灌木树种WUE日变化整体呈现递减趋势,即7:00WUE最高,17:00最低。针叶树种7:00WUE高达 (3.85±0.37) μmol·mmol-1,17:00仅有 (1.52±0.27 )μmol·mmol-1,降幅为35%~57%。灌木树种在7:00WUE为 (4.24±0.58) μmol·mmol-1,17:00为 (1.87±0.39 )μmol·mmol-1,7:00WUE是17:00的1.49~1.98倍。阔叶树种和经济林树种WUE均存在“午休”现象,下午有回升,但傍晚时分阔叶树种WUE出现降低趋势,而经济林树种WUE则继续升高。在整个生长季,针叶和经济林树种WUE均为倒“V”型趋势,生长季中期(6、7、8月)WUE最大,分别为 (2.73±0.25) μmol·mmol-1、(4.04±0.20) μmol·mmol-1。针叶树种生长季中期的WUE是初期(5月)和末期(9、10月)的1.12倍和1.04倍,经济林树种则为1.10倍和1.41倍。阔叶树种WUE表现为逐渐降低的趋势,生长季末期占初期的71%。灌木树种呈现出逐渐升高的趋势,升高幅度为5%。WUE年均值为经济林树种((3.65±0.12 )μmol·mmol-1)、阔叶树种((3.33±0.41) μmol·mmol-1)高于灌木树种((2.90±0.27 )μmol·mmol-1)和针叶树种((2.64±0.27) μmol·mmol-1),主要是因为消耗单位水分经济林树种果实的养分积累较多,阔叶树种年生长量较大。
2.3 相关性分析
树木的生长离不开水,而树木对水分的利用效率受到多方面因素的综合影响[18],WUE为Pn与Tr的比值,因此所有对Pn、Tr有影响的因子都会在不同程度上对WUE产生影响。表2为不同树种不同时间段的水分利用特性与各个影响因子之间的相关性综合分析。Pn与PAR、Gs、Tr呈极显著正相关关系(P<0.01),与Tl呈显著正相关关系,与Ci呈极显著负相关关系。Tr与t(a)、t(l)、PAR、Gs呈极显著正相关关系,与VPD呈显著正相关关系,与Ci呈极显著负相关关系。WUE与Pn、Gs呈极显著正相关关系(P<0.01),与PAR呈显著正相关关系(P<0.05),与VPD、t(a)、t(l)呈极显著负相关关系(P<0.01)。光合有效辐射PAR强,气孔导度Gs增加,光合速率Pn增大,WUE高。温度T逐渐升高,Tr、VPD加强,为减少水分散失气孔关闭,Pn降低,WUE低。
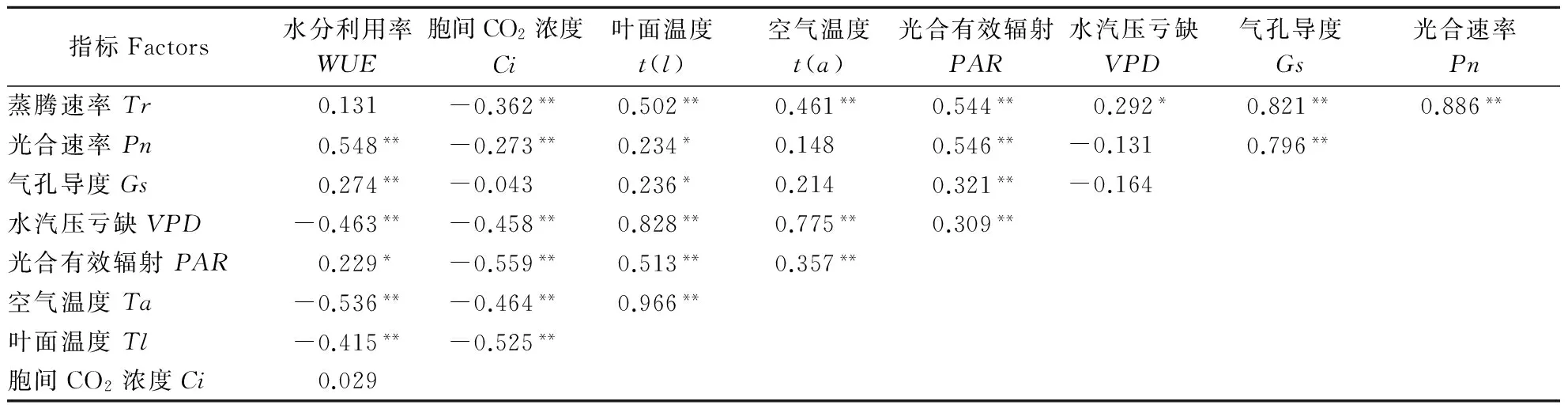
表2 不同树种不同时间段水分利用特性与影响因子相关性Table 2 Correlation analysis of water use characteristics and influencing factors of different tree species in different time periods
注:**在 0. 01 水平上显著相关,*在 0. 05 水平上显著相关。
Note:** denotes significant difference atP=0.01 probability level,* denotes significant difference atP=0.05 probability level.
3 结论与讨论
3.1 园林树种WUE的差异
植物的生长是多方面因素共同作用的结果,水又是最重要的要素之一。WUE体现了植物的生长状况,是反应植物耐旱性的一个重要指标。一般在相同的条件下,WUE越高植物耗水量相对越少,生成供自身生长的有机物质越多,释放的氧气越多,抗旱能力越强[19-20]。本研究中,不同种类树种WUE均表现为7:00-11:00明显高于13:00-17:00,这与张向峰等[20]对苦竹WUE的研究结果相一致。原因主要是清晨t(a)较低,光照强度较弱,Tr较低使得WUE较高,随着太阳辐射和t(a)的逐渐升高,Pn、Tr加强,光合作用存在光饱和和光抑制现象[21],使得Pn增加的速度相对于Tr增加的程度要小,因而出现了WUE降低的趋势。
针叶树种WUE日变化规律呈现出降低趋势,7:00-9:00时WUE为1 d中的最高值((3.85±0.37) μmol·mmol-1),15:00-17:00时WUE减弱到最低值((1.52±0.27 )μmol·mmol-1)。WUE整体表现为生长季中期((2.73±0.25 )μmol·mmol-1)>生长季末期((2.61±0.09 )μmol·mmol-1)>生长季初期((2.44±0.49) μmol·mmol-1)。阔叶树种WUE值日变化趋势表现出双峰曲线趋势,存在“午休”和“傍晚休”现象,即13:00-15:00最低,仅有 (1.96±0.44) μmol·mmol-1,7:00-9:00时WUE值最高,可达(4.87±1.28 )μmol·mmol-1。抗旱能力表现为生长季初期(3.82±0.56 μmol·mmol-1)>生长季中期((3.59±0.44) μmol·mmol-1)>生长季末期((2.70±0.23) μmol·mmol-1)。其中,刺槐WUE月变化则显现出逐渐减弱的趋势,王翼龙等[22]对刺槐WUE的研究也得出了相同的日变化趋势,但其测得的日均WUE为:7 月>10 月>6 月>8 月,与该研究得出的6月((3.31±0.56 )μmol·mmol-1)>7月((3.22±1.04) μmol·mmol-1)>8月((2.53±1.15 )μmol·mmol-1)>10月((2.30±0.65) μmol·mmol-1)略有不同,可能是由于试验树种年龄的不同所致。
经济林树种抗旱性表现为生长季初期((4.05±0.20) μmol·mmol-1)>生长季中期((4.04±0.20) μmol·mmol-1)>生长季末期((2.87±0.41) μmol·mmol-1)。WUE变化幅度较小,位于(2.81±0.50~4.47±0.84) μmol·mmol-1范围内,日动态显示出“V”型变化,即7:00WUE最高,存在水分利用“午休”现象(时间出现在13:00-15:00),这一结果与池永宽等[23]、巴合提牙尔·克热木等[24]的研究结果相同。灌木树种WUE日变化动态整体呈现下滑趋势,7:00出现最大WUE值,达到(4.24±0.58) μmol·mmol-1。抗旱能力表现为生长季末期((2.94±0.05) μmol·mmol-1)>生长季中期((2.92±0.07) μmol·mmol-1)>生长季初期((2.80±0.32) μmol·mmol-1)。此次试验得出,WUE表现为阔叶树种((3.32±0.43 )μmol·mmol-1)>针叶树种((2.74±0.19) μmol·mmol-1),与胡红玲等[25]得出的阔叶树种WUE低于针叶树种的结果不同,因此还有待于进一步的研究。
3.2 WUE的影响因子
影响植物叶片WUE的因子主要包括内因(光合途径、气孔等)和外因(太阳辐射、温度等),WUE是Pn和Tr的比值,WUE依赖于光合作用和蒸腾作用,因此,所有影响植物WUE的Pn和Tr的因素,都会影响植物WUE[26]。本次试验得出,树种WUE与Pn、Gs、VPD、t(a)、t(l)相关性极显著(P<0.01),与PAR相关性显著(P<0.05)。
本研究中,园林树种WUE与VPD、t(a)、t(l)呈极显著负相关关系(P<0.01),这与夏江宝等[27]对美国凌霄受环境因子影响的研究结果一致,其中WUE与VPD呈负相关关系,与刘贤赵等[18]对番茄单叶WUE的研究结果相同,但与其气孔阻力增加WUE提高的结果不同,主要是由于光照强度(遮荫)不同导致的。光照对WUE的影响主要是通过作用于光合作用和蒸腾作用来实现的,因为光是2者的驱动力,但影响方式不同。光对光合作用仅产生瞬时影响,而对蒸腾作用的影响是可在时空上累加的。温度t主要通过气孔关闭和光合作用羧化酶活性来影响叶片光合作用和蒸腾作用,进而影响WUE[26]。喻方圆等[28]、樊廷录等[29]、NICOTRA等[30]在研究针叶树、小米、天竺过程中,也得出了WUE与t呈极显著的负相关关系(P<0.01),但PEARMAN等[31]、刘晓宏等[32]的研究得出了正好相反的结果,即WUE和温度t呈正相关关系,因此关于温度t对WUE的影响,还有待进一步的研究。薛雪等[33]研究5 个常绿园林树种的夏季水分利用特性得出,WUE与Pn呈极显著正相关关系(P<0.01),与该试验的研究结果相同,即可通过提高植物的光合作用来增加植物的WUE,但与Gs呈极显著负相关关系与本研究不同,可能是由于试验观测期的长短不同引起的。根据影响因素的强弱不同以及各种类树木的生长特性,阔叶树种、经济林树种在生长季末期,针叶和灌木树种于生长季初期可通过提高光合速率和降低水汽压亏缺来提高水分利用率。
[1] 王熹,王湛,杨文涛,等.中国水资源现状及其未来发展方向展望[J].环境工程,2014(7):1-5.
[2] 陈国荣.园林树木的水分利用率研究概述[J].辽宁林业科技,2016(4):34-36.
[3] ZUO T Y. Recycling Economy and Sustainable Development of Materials, 3th China Conference on Membrane Science and Technology [R]. Beijing: Beijing University of Technology, 2007.
[4] WULLSCHLEGER S D, MEINZER F C, VERTESSY R A. A review of whole-plant water use studies in tree [J].Tree Physiology Tree ,1998,18(8/9):499-512.
[5] RICHARDS R A, REBETZKE G J, CONDON A G, et al. Breeding opportunities for increasing the efficiency of water use and crop yield intemperate cereals[J]. Crop Science, 2002,42(1):111-121.
[6] 吴俊文,刘珊,李吉跃,等.干旱胁迫下广东石漠化地区造林树种光合和耗水特性[J].生态学报,2016,36(11):3429-3440.
[7] MORRISON J L. Intercellular CO2 concentration and stomatal response to CO2 [S]//Zeiger, ed. Stomatal Function. Stanford: Stanford University press, 1987:229-251.
[8] MOTT K A. Sensing of atmospheric CO2 by plants [J]. Plant, Cell & Environment, 1990, 13(7): 731-737.
[9] AUGE R M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis [J]. Mycorrhiza, 2001, 11(1): 3-42.
[10] HETHERINGTON A M, Woodward F I. The role of stomata in sensing and driving environmental change [J]. Nature, 2003, 424(6951):901-908.
[11] 李机密,黄儒珠,王健,等.陆地植物水分利用率[J].生态学杂志, 2009, 28(8):1655-1663.
[12] 赵顺,黄秋娴,李玉灵,等.遮荫处理对臭柏幼苗光合特性的影响[J].生态学报,2014, 34(8):1994-2002.
[13] 刘茜.樟树叶片蒸腾特性及其与生理生态因子关系分析[J].中南林业科技大学学报,2009,29(1):1-5.
[14] 何雅冰.北京山区典型树种不同CO2浓度下水分利用效率研究[D].北京:北京林业大学,2016.
[15] 毛振华.不同灌溉量条件下六种地被植物的耗水特性[D].呼和浩特:内蒙古农业大学,2011.
[16] 胡化广,张振铭,吴生才,等.植物水分利用效率及其机理研究进展[J].节水灌溉,2013(3):11-15.
[17] 鲁绍伟,杨新兵,陈丽华,等.华北土石山区典型人工林耗水规律研究[M].北京:中国林业出版社,2008.
[18] 刘贤赵,康绍忠.遮荫对番茄单叶水分利用效率的影响研究[J].中国生态农业学报,2003,11(11):23-26.
[19] SOBRADO M A. Relation of water transport to leaf gas exchange properties in three mangrove species [J]. Trees, 2000, 14(5):258-262.
[20] 张向峰,王玉杰,王云琦,等.苦竹叶片光合及水分利用率特性[J].水土保持通报, 2012,32(1):122-126.
[21] SUKHBIR S, KENNETH J B, SANGAMESH V A, et al. Estimating water balance, evapotranspiration and water use efficiency of spring safflower using the CROPGRO model [J]. Agricultural Water Management, 2017, 185: 137-144.
[22] 王翼龙.黄土高原半干旱区两典型林分主要树种光合耗水特性及影响因素研究[D].北京:中国科学院研究生院,2010.
[23] 池永宽,熊康宁,王元素,等.西南石漠化地区两种经济林木光合日动态特征[J].经济林研究,2015,33(1):45-49.
[24] 巴合提牙尔·克热木,郭靖,徐业勇,等.南疆地区2个引种大樱桃品种光合特性对比研究[J].西北林学院学报,2016,31(2):86-91.
[25] 胡红玲,张健,万雪琴,等.巨桉与5种木本植物幼树的耗水特性及水分利用效率的比较[J]. 生态学报,2012,32(12):3873-3882.
[26] 韩若霜.呼和浩特几种园林树木水分生理特性及耐旱性评价[D].呼和浩特:内蒙古农业大学,2014.
[27] 夏江宝,张景宽,张光灿.水分胁迫对美国凌霄单叶水分利用效率的影响[J].西北植物学报,2009,29(1)0154-0159.
[28] 喻方圆,徐锡增, ROBERT D G. 水分和热胁迫处理对4种针叶树苗木气体交换和水分利用效率的影响[J].林业科学,2004,40(2):38-44.
[29] 樊廷录,宋尚有,徐银萍,等.旱地冬小麦灌浆期冠层温度与产量和水分利用效率的关系[J].生态学报,2007,27(11):4491-4497.
[30] NICOTRA A B, COSGROVE M J, COWLING A, et al. Leaf shape linked to photosynthetic rates and temperature optima in South African Pelargonium species [J]. Oecologia, 2008, 154(4):625-635.
[31] PEARMAN G I, FRANCEY R J, FRASER P J B. Climatic implications of stable carbon isotopes in tree rings [J]. Nature, 1976, 260 (5554):771-773.
[32] 刘晓宏, 赵良菊, MENASSIE G, 等.东非大裂谷埃塞俄比亚段内C3植物叶片δ13C和δ15N及其环境指示意义[J].科学通报,2007,52(2) :199-206.
[33] 薛雪,李娟娟,郑云峰,等.5个常绿园林树种的夏季光合蒸腾特性[J].林业科学,2015,51(9):150-155.
StudyonwaterusecharacteristicsandinfluencefactorsoflandscapeplantsinNorthChina
DING Jie1,2,3,LI Shaoning2,3,4,LU Shaowei2,3,4,DING Li1,YANG Chao1,2,3,CHEN Bo2,3,4, YANG Xinbing1
(1.College of Forestry, Agricultural University of Hebei, Baoding 071000, China; 2.Forestry and Pomology Institute, Beijing Academy of Agriculture and Forestry Sciences, Beijing 100093, China; 3.Beijing Collaborative Innovation Center for Eco-environmental Improvement with Forestry and Fruit Trees, Beijing 100093, China; 4.Beijing Yanshan Forest Ecosystem Research Station, Beijing 100093, China)
The water use properties of sixteen landscape plants were measured using CI-340 photosynthetic with the method of potted plants. Results show: 1) Landscape plants’WUEat 7:00-11:00 were obviously higher than that at 13:00-17:00. The daily changes of coniferous and shrub trees’WUEshowed the overall downward trend, and that of the broadleaf and economic tree species had “lunch break” phenomenon. 2) Economic tree species’WUEwas the highest at vigorous growth period. It was (4.04±0.20)μmol·mmol-1. Broad leaved tree species’WUEwere the biggest (3.82±0.56μmol·mmol-1) at early growing season. Shrub tree species’WUEwere the highest (2.94±0.05μmol·mmol-1) at the end of growing season. Coniferous tree species’WUEwas the highest at vigorous growth period. It was 2.73±0.25μmol·mmol-1. 3) TheWUEorder was economic tree species ((3.65±0.12)μmol·mmol-1) > deciduous tree species ((3.33±0.41)μmol·mmol-1) > shrub tree species ((2.90±0.27)μmol·mmol-1) > coniferous tree species ((2.64±0.27)μmol·mmol-1), and economic tree species’WUEwas the 1.38 times of the coniferous tree species’. 4) TheWUEpresented a very significant positive correlation (P<0.01) with Pn and Gs, and showed a significant positive correlation (P<0.05) with PAR, and a very significant negative correlation (P<0.01) withVPD,t(a) andt(l). The results of the study provided theoretical basis for the rational use of water resources and the scientific management of landscape greenbelt.
landscape plants; water use efficiency; influence factors
2017-03-18
北京市农林科学院青年基金项目(QNJJ201532);创新能力建设专项项目(KJCX20150205); 2011协同创新中心(PXM2017_014207_000043);北京市农林科学院科技创新团队项目(JWKST201609)
丁 杰(1992-),女,河北固安人,硕士研究生,从事水土保持方面的研究。
鲁绍伟(1969-),男,河北抚宁人,研究员,博士;杨新兵(1978-),男,河北涉县人,副教授,博士。
1000-2340(2017)04-0487-08
S 714.7
A
(责任编辑:李 莹)
猜你喜欢
林业科技(2020年3期)2021-01-21
意林·少年版(2020年17期)2020-10-12
农民致富之友(2020年17期)2020-06-19
发明与创新·中学生(2020年4期)2020-04-17
山东林业科技(2019年2期)2019-06-03
红土地(2018年7期)2018-09-26
现代园艺(2018年2期)2018-03-15
现代农业研究(2017年2期)2017-04-01
科学与财富(2016年26期)2016-12-01
环球人文地理·评论版(2014年5期)2014-08-12
