
不同连作年限怀牛膝根际土壤理化性质及微生物多样性
2017-10-13 12:32王娟英许佳慧吴林坤吴红淼孔露霏林文雄
生态学报 2017年17期
王娟英,许佳慧, 吴林坤, 吴红淼, 朱 铨, 孔露霏, 林文雄,*
1 福建农林大学生命科学学院, 福州 350002 2 福建农林大学生物农药与化学生物学教育部重点实验室, 福州 350002 3 福建农林大学农业生态过程与安全监控重点实验室, 福州 350002
不同连作年限怀牛膝根际土壤理化性质及微生物多样性
王娟英1,2,3,许佳慧2,3, 吴林坤1,2,3, 吴红淼1,2,3, 朱 铨1,2,3, 孔露霏1,2,3, 林文雄1,2,3,*
1 福建农林大学生命科学学院, 福州 350002 2 福建农林大学生物农药与化学生物学教育部重点实验室, 福州 350002 3 福建农林大学农业生态过程与安全监控重点实验室, 福州 350002
以不同连作年限怀牛膝根际土壤为材料,采用土壤农化分析技术和磷脂脂肪酸生物标记法,对土壤理化性质以及根际微生物群落结构进行分析。结果表明:怀牛膝连作会使大部分大量元素(全磷、碱解氮、速效磷和速效钾)含量上升,而全氮、全钾的变化并无明显趋势。全量钙、全量镁、全量锰和有效锌的含量也会随着连作年限的增加而增加。研究发现,怀牛膝连作1a的土壤与对照相比,根际土壤微生物群落结构差别不大,但是与连作10a和30a的土壤相比微生物多样性却发生了明显变化。土壤中总PLFAs、细菌、革兰氏阴性菌Gram(-)及真菌含量都随连作年限增加而明显上升,微生物多样性增加,且革兰氏阳性菌Gram(+)/革兰氏阴性菌Gram(-)的比值随着连作年限的增加而下降。此外,土壤中一些嗜热解氢杆菌和真菌等参与土壤物质循环和木质素降解的微生物类群数量明显增加,假单胞菌等有益微生物类群也显著增多,这是牛膝连作促进作用的生物学基础。研究用药用植物怀牛膝证明了植物根际微生物结构和功能多样性对植物连作促进或连作抑制作用的产生与控制的生态学意义。
怀牛膝;连作促进;磷脂脂肪酸;根际微生物
Abstract: In this study, the rhizosphere soils ofAchyranthesbidentata, which was grown in a monoculture for multiple years, were investigated for analyzing the major elements, trace elements and rhizosphere microbial community structure by means of agricultural chemistry and phospholipid fatty acid (PLFA) analysis techniques. The results showed that consecutive monocultures ofA.bidentataincreased the contents of major elements (total phosphorus, alkaline hydrolysis nitrogen, available phosphorus, and available potassium). However, the ability of consecutive monocultures to change the total nitrogen content and total potassium was not obvious. In addition, total calcium, total magnesium, total manganese, and available zinc contents also increased with the increasing years of monoculture. Further analysis of PLFA profiles showed that there were no obvious differences in the rhizosphere microbial community structures between the control and newly plantedA.bidentata. Besides, different results were depicted by 10 years and 30 years of consecutive monoculture, which showed that the total PLFA, bacteria (especially G-), and fungal content increased significantly, and the G+/G-ratio decreased with increasing years of monoculture. Moreover, some microbes involved in material cycling and lignin degradation, such asHydrogenobacter, fungi (PLFA18:2ω6,9), and some beneficial bacteria includingPseudomonasincreased significantly with increasing years of monoculture. These results suggest the important biological basis for the promotion mechanism ofA.bidentataunder a monoculture system. The case study exemplifies the ecological significance of rhizosphere microbial structure and its functional diversity in monoculture promotion or monoculture inhibition.
KeyWords:Achyranthesbidentata; continuous cropping promotion; phospholipid fatty acid; rhizosphere microorganisms
怀牛膝(AchyranthesbidentataBl.)为苋科牛膝属多年生草本植物,主产于我国河南温县、武陟、博爱及沁阳等地,是著名的“四大怀药”之一。根据《中华人民共和国药典》记载,牛膝性味甘苦、酸平,具有补肝肾、散淤血、填精补髓、益阴活血之功效[1]。在长期的生产实践过程中,人们发现大约有70%以上的药用植物连续在同一块田地上种植后,会出现土壤微生态环境恶化,自身生长发育不良,产量与药用品质下降等现象,即连作障碍(continuous monoculture problem)。根据文献报道,大部分块根类药材(如地黄、人参、三七、当归等)常会出现这种现象,因而称之为忌连作药用植物[2- 5]。然而,与之相反的是,怀牛膝是一种非常耐连作的道地药材。近些年来作者在调查中发现,头茬怀牛膝品质较差,而连作5a后其品质明显改善。当连作10a以上时,怀牛膝地下部即药用部位生长良好,主根又长又粗,须根和侧根少,产量高、品质好[6-7]。
有研究表明,药用植物连作障碍的产生与其根系分泌物介导下,根际微生物结构变异,病原菌增多,有益菌下降有关[8]。而连作促进的产生及其机制至今仍是个迷。李娟等[9]运用HPLC等技术测定牛膝主要药用活性成分甾酮和皂苷含量,结果显示麦茬地中首次种植的牛膝药材中甾酮含量(0.5401 mg/g)和皂苷含量(1.0462 mg/g)均低于多年怀牛膝-小麦复种连作地中牛膝的甾酮(0.7493 mg/g)和皂苷含量(2.4381 mg/g),因而推断连作促进可能与连作引起土壤中药用活性成分的累积有关。然而,这些物质能否有效促进药用植物本身的生长发育,未见直接的试验证据报道。此外,有研究结果表明,植物根系分泌物进入土壤后易被土壤微生物降解,很难达到实验室条件下的有效作用浓度[10],因而,产生直接化感(direct allelopathy)促进或抑制作用的可能性不大[11]。近年来,越来越多的研究结果认为,植物化感作用(包括连作化感促进或连作化感抑制作用)是由植物根系分泌物介导而间接产生的化学生态学现象。换一句话说,植物化感作用是植物与土壤微生物相互作用的结果。在这一过程中,根际环境作为土壤微生物活动的一个复杂的、动态的微生态系统,在植物生长、吸收、分泌过程中扮演着重要角色。诸多研究表明,根际土壤微生物多样性参与根际生态系统的物质转化和循环,在维持土壤健康方面扮演着越来越重要的角色[12-13]。因此,从植物-微生物根际互作角度,探讨怀牛膝连作促进的根际生态学效应是一项亟待解决的问题。
研究植物根际土壤微生物的群落结构变化是植物-微生物根际互作机制的重要内容之一。通常可采用可培养微生物法、磷脂脂肪酸生物标记法(PLFAs)和BIOLOG技术、限制性片段长度多态性(T-RFLP)、变性梯度凝胶电泳(DGGE)、同位素标记等方法[14]。磷脂脂肪酸生物标记法(PLFA)主要根据不同类群微生物的指示PLFAs不同,具有高度专一性,能够快速、直接提供土壤微生物的生物量和群落结构的信息,且实验步骤要求较低,因此,该方法仍是目前较为常用的和有效的一种微生物群落结构分析方法[15-16]。马琨等[17]采用PLFAs方法研究发现,连作与间作处理下,马铃薯根际土壤微生物群落结构差异大,具体表现在马铃薯与蚕豆间作下,其根际效应比马铃薯与玉米间作的明显。李奇松等[18]研究也发现在无隔和网隔处理(即存在间作作物地下部根系互作)时,玉米和花生间作不仅是产量高而且根际微生物总PLFAs、细菌、真菌都高于全隔(即不存在间作作物地下部根系互作)处理。可见PLFAs至今仍是经济有效的方法。据此,本研究采用PLFAs技术,以不同连作年限牛膝根际土壤为研究对象,探讨牛膝根际土壤微生物群落结构随连作年限增加的动态变化,为阐明牛膝的连作促进机理提供依据。
1 材料和方法
1.1 材料
以“核桃纹”牛膝为供试品种,同一时间种植在不同连作年限的土壤中,在其块根膨大期,采用五点取样法收集不同土壤样本,即收集以下4个处理的土壤样本:(1)对照土,即对应于各处理,且只种前作作物小麦的土壤(CK);(2)连作牛膝1a的土壤;(3)连作10a的土壤;(4)连作30a的土壤。田地种植模式采用不同年限的复种连作,即前茬种植小麦,收获后种植牛膝。
怀牛膝播种期为7月20日,花期7—9月,果期9—10月。播种前,施有机复合肥1200kg/hm2,翻地约50cm左右再整地播种。之后在整个牛膝生长发育期间均采用相同的田间管理措施。本试验于怀牛膝块根膨大期(9月20日)取样,土壤样品采用五点取样法,牛膝根际土壤取样方法如下:首先用铲子去除怀牛膝地上部表层土壤,然后用铲子挖出地下部块根,轻轻抖去根围土,收集块根表面根际土。收集的土壤样品过2mm孔径筛子后分成两部分,一部分风干用于测定土壤理化性质,另一部分用于提取土壤微生物磷脂脂肪酸。
1.2 土壤理化性质测定方法
1.2.1 土壤中大量元素的测定
称取风干后土壤1.000g土装入开氏瓶中,加入5mL浓硫酸,电压调制360V,消煮3h至土变白。待其稍冷却后,加入500μL过氧化氢,将电压调制230V,以后每隔5min加少于500μL 过氧化氢,直至上清液变澄清后取出开氏瓶。加入20mL双蒸水,过滤,定容100mL。滤液保存用于全N、全P和全K的测定。碱解N的测定:采用碱解扩散法。速效P的测定:采用(碳酸氢钠浸提)钼锑抗比色法。速效K的测定:采用醋酸铵浸提原子吸收法[19]。
1.2.2 土壤中微量元素的测定
全量微量元素测定:将土壤风干后过孔径为2mm的塑料筛,称取0.1000g土样,加入5mL浓硝酸和1mL的30%过氧化氢,经微波消解,微波仪工作条件如表1。

表1 微波仪工作条件
待冷却后,将液体转移至10mL离心管,10000r/min离心3min。将上清液倒出,用超纯水定容至25mL,最后过滤备用,同时做空白实验。测定时采用WFX- 130A原子吸收分光光度计来测量土壤中全量Fe、Mn、Zn、Cu、Ca、Mg的含量[20](表2)。
有效态铜、锌、铁、锰的测定:采用二乙三胺五乙酸(DTPA)浸提法[21],称取风干后怀牛膝根际土壤10.00g于50mL离心管中,加入新鲜配制的DTPA浸提液20mL,盖好瓶盖,置于25℃,180r/min条件的摇床上摇3h,立即过滤。滤液立即用原子吸收分光光度计测定。除了标曲用DTPA浸提液配置外,浓度及仪器工作条件与全量元素测定方法相同。
1.3 土壤微生物群落PLFA生物标记分析
土壤微生物群落结构分析:采用磷脂脂肪酸(PLFA)生物标记法进行测定[22]。操作步骤为:将20mL的0.2mol/L的KOH甲醇溶液和4g的新鲜土样加到50mL的离心试管中,混合均匀,在37℃下温育1h(磷脂脂肪酸释放,并甲脂化,样品10min摇匀1次)。加入3mL 1.0mol/L的醋酸溶液(中和pH值),充分混匀后加10mL正己烷(使PLFAs转到有机相中),600rpm/min离心15min,然后将上层正己烷转到干净试管中,用冷冻干燥仪挥发掉溶剂。最后将PLFAs溶解在1mL体积比为1∶1的正己烷∶甲基丁基醚溶液中,再加入10μmol/L的内标19∶0,过滤后用作GC分析。

表2 WFX-130A原子吸收分光光度计火焰法工作条件
采用美国Agilent 6890N型气相色谱仪,条件:二阶程序升高柱温,70℃起始1min,接着20℃/min升至170℃,持续2min,尔后5℃/min升温至280℃,维持5min,最后以40℃/min升温至300℃,1.5min后结束。进样量1μL,进样分流比100∶1。
1.4 数据分析
方差分析采用Excel和DPS 3.0结合,数据均采用单因子方差,多重比较方法选用Tukey法(P<0.05)来分析。当分析不同连作年限牛膝土壤的4个样本之间土壤微生物PLFAs的分布特性时,将数据对数转化,以欧氏距离为聚类尺度,用(WPGMA)进行系统聚类。并采用Canoco软件进行主成分(PCA)分析。
2 结果与分析
2.1 土壤理化性质分析
2.1.1 土壤中大量元素分析
从表3可见,除了全氮、全钾并无明显的变化趋势外,牛膝连作下,土壤中全磷、碱解氮、速效磷和速效钾的含量均上升。进一步分析发现,在连作10a的土壤中这些营养成分的含量已经有明显的增加,而在连作30a的土壤中最高。可见,怀牛膝连作会使土壤大部分大量元素(特别是速效的元素)含量上升。
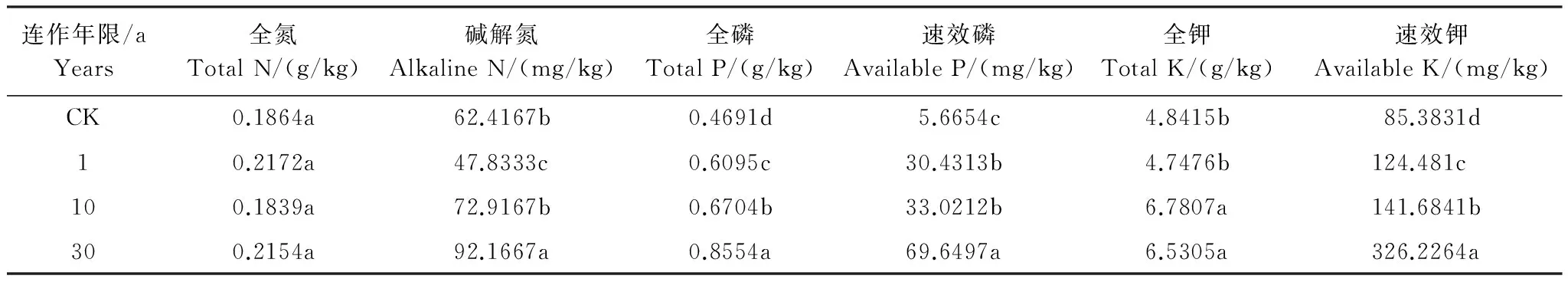
表3 不同连作年限怀牛膝根际土壤中大量元素的含量
CK:表示对照土壤CK stands for control soil; 同一列不同字母表示不同样品差异水平达 0. 05
2.1.2 土壤中微量元素分析
试验分析发现:除了土壤的全量钙、全量镁、全量锰含量随着连作年限的增加而增加外,其它3种元素(如全量铁、全量锌和全量铜)的含量并无明显增加的趋势,有的甚至还呈现下降的趋势(表4)。对土壤有效态微量元素分析还发现,除了土壤有效锌含量随着连作年限的增加而增加外,土壤中有效铁、有效锌和有效锰含量都呈先下降后上升变化的规律(表5),这说明怀牛膝连作会影响土壤中全量微量元素的变化,但是对于土壤有效态的微量元素却影响不显著。

表4 不同连作年限怀牛膝根际土壤中全量微量元素的含量
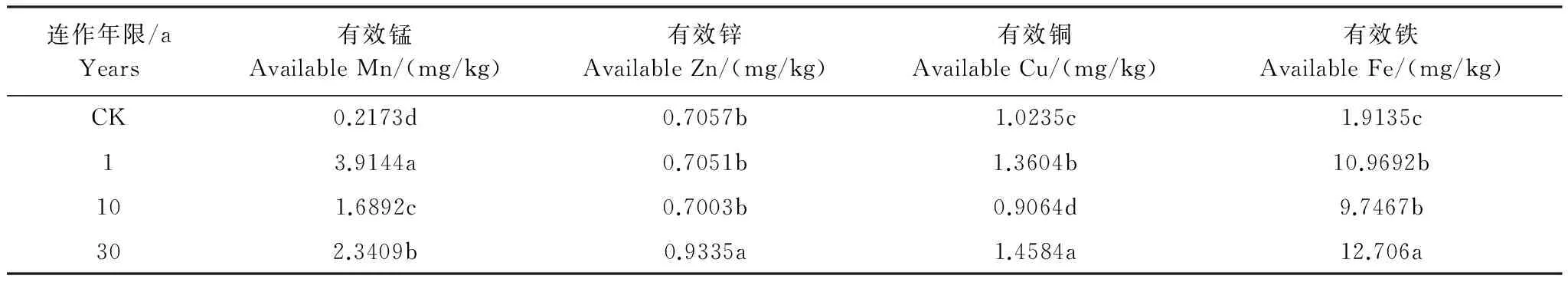
表5 不同连作年限怀牛膝根际土壤中有效态微量元素的含量
2.3 不同连作年限怀牛膝根际土壤微生物群落PLFAs检测
磷脂脂肪酸结果显示,共检测到15种PLFAs的生物标记(表6)。包括细菌的PLFAs生物标记,如12:0、16:0;革兰氏阳性细菌PLFAs生物标记,如i14:0、a15:0、a17:0;革兰氏阴性细菌PLFAs生物标记,如16:1ω9t、16:1 ω 7c、cy19:0;放线菌PLFAs生物标记,如10Me18:0;真菌PLFAs生物标记,如18:2 ω6,9、18:1 ω9c;嗜热解氢杆菌和原生生物的PLFAs生物标记,分别是18:0和20:4 ω6c(6,9,12,15),种类丰富。除了在1a的根际土壤里未检测到原生生物和13:0的PLFAs标记外,其他磷脂脂肪酸在所有土壤里均有分布。且种植过牛膝的根际土壤PLFAs丰富度大部分都高于对照土壤。进一步分析还发现,特别是16:0这类磷脂脂肪酸的含量随着连作年限的增加而增加,且增长幅度非常明显。而i14:0的含量则呈下降趋势。
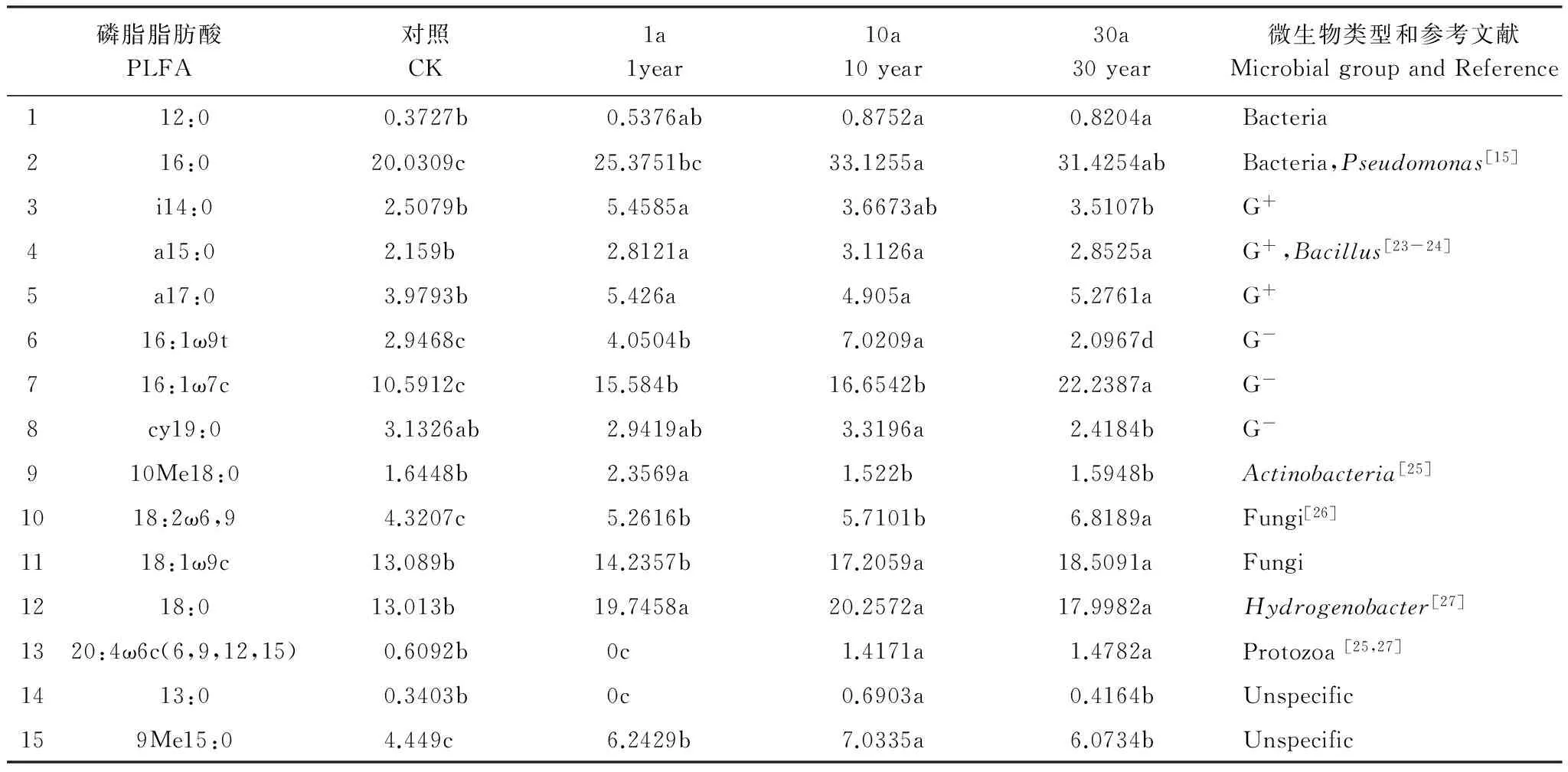
表6 不同连作年限牛膝根际土壤中各PLFAs的浓度
浓度单位 concentration unit:(nmol/g d.m. soil);1a表示种植牛膝第1年土壤 1 year refering to the rhizospheric soils ofAchyranthesbidentatamonocultured for one year;10a表示种植牛膝第10年土壤 10 year refering to the rhizospheric soils ofA.bidentatamonocultured for ten years,30a表示种植牛膝第30年土壤 30 year refering to the rhizospheric soils ofA.bidentatamonocultured for thirty years
2.4 不同连作年限怀牛膝根际土壤PLFAs种类和含量变化
从表7可以看出,不同连作年限怀牛膝根际土壤磷脂脂肪酸(PLFAs)生物标记种类和含量差异明显。但是不同连作年限的怀牛膝土壤微生物种类和含量变化趋势相似,都呈现出细菌>革兰氏阴性细菌G->真菌>革兰氏阳性细菌G+的趋势。并且发现与连作1a的土壤相比,除了G+的含量下降之外,连作10a和30a的土壤中其他的微生物(包括总PLFAs、细菌、G-、真菌)含量都明显上升。另外,G+/G-的比值随着连作年限的增加而下降,而真菌/细菌得比值表现出相反的变化趋势。
表7不同连作年限牛膝根际土壤中总PLFAs和各类菌所占百分比
Table7ThetotalPLFAsandpercentageofdifferentspecificmicrobialPLFAstototalonesinrhizosphericsoilsofAchyranthesbidentatamonoculturedforthedifferentyears

项目Item对照CK1a10a30a总PLFATotalPLFA83.1863c110.0285b126.5162a123.5279a细节Bacteria54.933a56.504a57.4441a57.1649aG+10.3961b12.4356a9.2338b9.4127bG-20.0886a20.5265a21.3383a21.6644a真菌Fungi20.9642a17.7198b18.1142b20.5061a细菌/真菌G+/G-/%51.8839b60.5722a43.2946c43.48cFungi/Bacteria/%38.1893a31.367b31.5483b35.9248ab
Total PLFA浓度单位concent unit:(nmol/g d.m. soil);其它单位用“%”表示the other units indicated in “%”
2.5 基于PLFAs生物标记的不同连作年限怀牛膝聚类及主成分分析
将所有土壤样品(6个处理,3次重复)进行主成分分析(图1A),发现牛膝连作对根际微生物群落结构有明显影响。主成分1(PC1)和主成分2(PC2)分别解释总方差变量的44.3%和74.2%。对照和连作1a的怀牛膝根际土壤距离相近,均分布于主成分1的正端,连作10a与连作30a的距离较近,均分布于主成分1的负端。如图1B所示,当欧式距离为14.23时,不同连作年限怀牛膝根际土壤可聚为两大类,第一类包括CK,连作1a怀牛膝根际土壤,而连作10a和连作30a的怀牛膝根际土壤则聚在第二类中,说明这些土壤微生物多样性相类似,生物量相近。这结果与主成分分析的结果相类似。综合主成分与聚类分析结果可以看出,连作年限短的土壤样品(如连作1a)明显区别于连作年限长的土壤样品(如连作10a、连作30a)。

图1 不同连作年限牛膝根际土壤中PLFAs生物标记主成分分析(A)和聚类分析(B)Fig.1 Classification of PLFAs biomarks in rhizospheric soils of Achyranthes bidentata monocultured for different years by principal component (A) and WPGMA cluster analysis (B)
2.6 不同连作年限牛膝根际土壤中各类PLFAs和大量元素冗余分析(RDA)
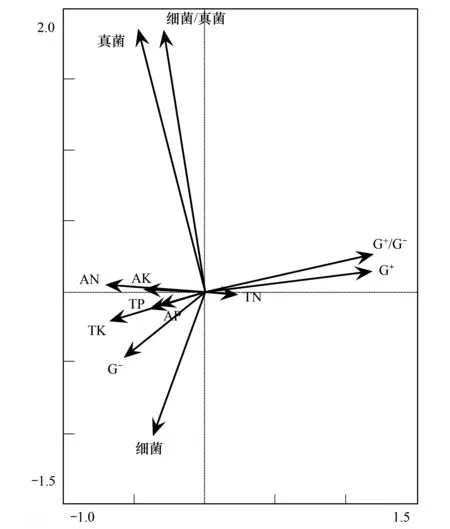
图2 不同连作年限牛膝根际土壤中各类PLFAs和大量元素冗余分析Fig.2 Redundancy analysis (RDA) between group-specific phospholipid fatty acid (PLFAs) and soil nutrients variables in rhizospheric soils of Achyranthes bidentata monocultured for different years
从RDA(图2)结果发现各类PLFAs以及营养元素均分布在3个象限内。进一步分析表明G+/G-和G+和绝大多数环境变量和各类微生物菌群呈负相关,而与全氮呈正相关关系。细菌以及革兰氏阴性菌的含量和绝大多数的元素(包括全磷、速效氮、速效磷、全钾、速效钾)含量密切相关。而真菌以及真菌/细菌的比值与元素含量关系不大。
3 讨论
前人研究表明,在单一化种植模式下,农作物减产的原因可大致总结为以下3点:(1)土壤养分匮乏或失调[28-29];(2)根系分泌物中化感物质的自毒效应[30];(3)连作土壤微生态结构失衡[31-32]。对于这3个原因,不同的学者有不同研究结论与看法。孙冰玉[33]研究认为,连作下,东北烤烟根际土壤肥力发生了很大变化,土壤有机质含量、供钾和供磷水平明显不足,且必需元素等都显著降低。但是,韩春丽等[34]发现连作大豆土壤中微量元素锌、锰、铁、铜含量的变化在不同年限中并无明显下降变化规律。近年来,不少学者研究认为同一种植物长时间在同一田块上种植,会造成了土壤中有益微生物数量的减少及有害病原菌数量的增长,在此消彼长的过程中影响植物体的正常功能、生长发育等生命活动。华菊玲等[35]发现,连作下芝麻根际土壤有益菌(芽孢杆菌Bacillus)的菌群数量明显低于新种植地,而尖孢镰孢菌(Fusariumoxysporum)、青枯劳尔氏菌(Ralstoniasolanacearum)的含量显著高于新种植地,正是这样的微生物区系失衡导致了芝麻的连作障碍产生,因而导致减产降质,其结果充分说明了土壤微生物群落结构与功能多样性的重要性。
本研究结果提供了一个作物连作促进的特例。研究发现怀牛膝连作会使得土壤中有效态的大量元素含量增加,微量元素则相反。然而,对全量大量元素影响却不明显。郝慧荣等[36]和Chen等[6]研究结果也认为,与牛膝连作2a土壤相比,连作20a的土壤有机质、全氮、全磷、全钾、速效氮(铵态氮)和速效磷均有所增加,尤其以全磷、速效氮(铵态氮)和速效磷增加较为显著,说明怀牛膝连作一定程度上改善了根际土壤环境,提高了土壤的保肥能力和补给能力,不仅有利于植株的生长,还为根际微生物活动提供营养和能源。据此,本研究从土壤微生物角度入手,利用磷脂脂肪酸生物标记法,研究连作下怀牛膝根际土壤微生物的变化情况。结果表明,在连作条件下,怀牛膝根际土壤微生物群落结构明显发生了改变,即连作年限越长,微生物群落结构差异越大,且土壤中细菌特别是革兰氏阴性细菌的数量呈上升趋势,革兰氏阳性细菌的数量变化不大,所以G+/G-比值明显下降。前人研究认为该比值下降,说明土壤中可利用养分逐渐增多,佐证了我们对牛膝土壤有效养分随连作年限延长而增多的结论[37]。进一步分析发现,随连作年限的延长,土壤中一些嗜热解氢杆菌、放线菌和真菌等一些参与土壤物质循环和木质素降解菌[25- 27]的微生物类群的数量也明显提高,这是促进牛膝连作土壤养分有效化不断增强的生物学基础。此外,本研究还发现,随连作年限的延长,土壤中一些有益微生物类群如假单胞菌Psudomonas(PLFA16:0)[15]芽孢杆菌Bacillus(PLFAa15:0)[23-24]也显著增多(表6)。作者所在的课题组郝慧荣等[36]和Chen等[6]在对牛膝根际土壤微生物群落结构分析时发现,连作20a牛膝土壤微生物对氨基酸类的代谢活性较高,对多聚物、酚酸类、羧酸类的代谢活性较低,放线菌和原生动物的PLFA相对含量比连作2年分别高出1.4%和2.3%;T-RFLP分析发现,一些促生根际菌(PGPR,以芽孢杆菌属Bacillus和类芽孢杆菌属Paenibacillus为主),包括假单胞菌属Pseudomonas、枯草芽孢杆菌Bacillussubtilis、浸麻类芽孢杆菌(Paenibacillusmacerans)、环状芽孢杆菌(Bacilluscirculans)、球形芽孢杆菌(Bacillussphaericus)等和一些改善土壤质地、防治植物病害的功能菌(如链霉菌属Streptomycetes等)随着连作年限的增加而不断增加,而以支原体属(Mycoplasma)为主的病原菌则不断减少,这是牛膝连作促进的重要根际生物学特性。
综上所述,植物根际微生物结构与功能多样性对植物的健康生长有着极其重要的作用。近年来,有学者[38]将植物根际微生物组称为植物的第二基因组学,已引起国内外学者的普遍关注。众所周知,植物根系分泌物是介导根际微生物变化的诱导因子。不同的植物以及植物在其生长发育的不同阶段,其根系会分泌各种次生代谢产物,而这些植物根系分泌物能产生间接的生态效应,并有效抑制某些类群微生物的生长,同时也会有效促进另一些类群微生物的生长,从而影响根际微生物的区系结构和功能多样性,最终导致土壤微生物群落的选择性效应[39- 41]。其中最为典型的就是豆科植物与根瘤菌的共生关系。豆科植物根系会分泌一种类黄酮物质,从而诱导根瘤菌在其根系表面定殖,从而形成共生关系,这就说明根系分泌物与根际微生物之间存在着密切关系。覃逸明等[42]采用高效液相色谱法(HPLC)对药用植物丹凤(PaeoniaostiiT.)自毒物质进行检测,发现根系分泌物的浓度(包括阿魏酸、肉桂酸、香草醛、香豆素和丹皮酚)随着连作年限的增加而上升,进而影响丹凤地下部根长、根系活力以及地上部植株的生物量。Zolla等[43]在研究干旱胁迫对拟南芥植株生长的影响时发现,种植在同域土壤中的植物生物量要高于非同域土壤的,这是由于同域土壤中存在许多根际促生菌的原因。本研究结果发现随着连作年限增加,总PLFAs生物量和细菌总量特别是革兰氏阴性细菌和一些有益菌含量都在增加。这可能就是因为根系分泌物介导下,植物根际促生菌(PGPR)在土壤中产生富集作用的结果。然而,究竟是何种牛膝根系分泌物,且如何介导连作牛膝根际土壤微生物差异变化及其作用的内在机制尚需深入研究。
[1] 孟大利, 侯柏玲, 汪毅, 张鹏, 李铣. 中药牛膝中的植物甾酮类成分. 沈阳药科大学学报, 2006, 23(9): 562- 564, 576- 576.
[2] 郭兰萍, 黄璐琦, 蒋有绪, 吕冬梅. 药用植物栽培种植中的土壤环境恶化及防治策略. 中国中药杂志, 2006, 31(9): 714- 717.
[3] 吴林坤, 黄伟民, 王娟英, 吴红淼, 陈军, 秦贤金, 张重义, 林文雄. 不同连作年限野生地黄根际土壤微生物群落多样性分析. 作物学报, 2015, 41(2): 308- 317.
[4] 李勇, 刘时轮, 黄小芳, 丁万隆. 人参(Panaxginseng)根系分泌物成分对人参致病菌的化感效应. 生态学报, 2009, 29(1): 161- 168.
[5] 陆晓菊, 官会林, 张正芸, 马永存, 唐蜀昆. 三七连作根际土壤微生物区系的16S rRNA系统遗传多样性. 微生物学报, 2015, 55(2): 205- 213.
[6] Chen T, Li J, Wu L K, Lin S, Wang J H, Li Z F, Zhang Z Y, Lin W X. Effects of continuous monoculture ofAchyranthesbidentataon microbial community structure and functional diversity in soil. Allelopathy Journal, 2015, 36(2): 197- 211.
[7] 李振方. 怀牛膝连作促进效应及其分子机理研究[D]. 福州: 福建农林大学, 2008.
[8] Wu L K, Wang J Y, Huang W M, Wu H M, Chen J, Yang Y Q, Zhang Z Y, Lin W X. Plant-microbe rhizosphere interactions mediated byRehmanniaglutinosaroot exudates under consecutive monoculture. Scientific Reports, 2015, 5: 15871.
[9] 李娟, 黄剑, 李萍, 张重义. 连作对牛膝中甾酮及三萜类物质含量的影响. 时珍国医国药, 2010, 21(10): 2433- 2434.
[10] Lin R Y, Wang H B, Guo X K, Ye C Y, He H B, Zhou Y, Lin W X. Impact of applied phenolic acids on the microbes, enzymes and available nutrients in paddy soils. Allelopathy Journal, 2011, 28(2): 225- 236.
[11] Wu H M, Wu L K, Wang J Y, Zhu Q, Lin S, Xu J H, Zheng C L, Chen J, Qin X J, Fang C X, Zhang Z X, Azeem S, Lin W X. Mixed phenolic acids mediated proliferation of pathogensTalaromyceshelicusandKosakoniasacchariin continuously monoculturedRadixpseudostellariaerhizosphere soil. Frontiers in Microbiology, 2016, 7: 335.
[12] Jacobsen C S, Hjelmsø M H. Agricultural soils, pesticides and microbial diversity. Current Opinion in Biotechnology, 2014, 27: 15- 20.
[13] Rutigliano F A, Castaldi S, D′Ascoli R, Papa S, Carfora A, Marzaioli R, Fioretto A. Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment. Applied Soil Ecology, 2009, 43(1): 40- 46.
[14] 吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望. 植物生态学报, 2014, 38(3): 298- 310.
[15] 张秋芳, 刘波, 林营志, 史怀, 杨述省, 周先冶. 土壤微生物群落磷脂脂肪酸PLFA生物标记多样性. 生态学报, 2009, 29(8): 4127- 4137.
[16] Wu Z Y, Haack S E, Lin W X, Li B L, Wu L K, Fang C X, Zhang Z X. Soil microbial community structure and metabolic activity ofPinuselliottiiplantations across different stand ages in a subtropical area. PLoS One, 2015, 10(8): e0135354.
[17] 马琨, 杨桂丽, 马玲, 汪春明, 魏常慧, 代晓华, 何文寿. 间作栽培对连作马铃薯根际土壤微生物群落的影响. 生态学报, 2016, 36(10): 2987- 2995.
[18] Li Q S, Wu L K, Chen J, Khan M A, Luo X M, Lin W X. Biochemical and microbial properties of rhizospheres under maize/peanut intercropping. Journal of Integrative Agriculture, 2016, 15(1): 101- 110.
[19] 王昭艳, 左长清, 曹文洪, 杨洁, 徐永年, 秦伟, 张京凤. 红壤丘陵区不同植被恢复模式土壤理化性质相关分析. 土壤学报, 2011, 48(4): 715- 724.
[20] 黄明堦, 魏道智, 陈燕丹, 蔡碧琼, 游秀花. 荷叶无机营养元素含量及溶出率. 河南科技大学学报: 自然科学版, 2010, 31(5): 80- 82, 86- 86.
[21] 杜静静. 不同种植年限果园土壤理化性质与酶活性研究[D]. 临汾: 山西师范大学, 2013.
[22] Wu L K, Li Z F, Li J, Khan M A, Huang W M, Zhang Z Y, Lin W X. Assessment of shifts in microbial community structure and catabolic diversity in response toRehmanniaglutinosamonoculture. Applied Soil Ecology, 2013, 67: 1- 9.
[23] 刘国红. 芽孢杆菌的分类鉴定及其相关属的分类系统演变研究[D]. 福州: 福建农林大学, 2009.
[24] 刘贯锋. 芽胞杆菌分类学特征及其生防功能菌株筛选[D]. 福州: 福建农林大学, 2011.
[25] 刘波, 胡桂萍, 郑雪芳, 张建福, 谢华安. 利用磷脂脂肪酸(PLFAs)生物标记法分析水稻根际土壤微生物多样性. 中国水稻科学, 2010, 24(3): 278- 288.
[26] 郁红艳. 农业废物堆肥化中木质素的降解及其微生物特性研究[D]. 长沙: 湖南大学, 2007.
[27] 付学琴, 黄文新. 不同树龄南丰蜜橘根际土壤微生物群落多样性分析. 园艺学报, 2014, 41(4): 631- 640.
[28] 秦舒浩, 曹莉, 张俊莲, 师尚礼, 王蒂. 轮作豆科植物对马铃薯连作田土壤速效养分及理化性质的影响. 作物学报, 2014, 40(8): 1452- 1458.
[29] 简在友, 王文全, 孟丽, 王丹, 游佩进, 张子龙. 人参连作土壤元素含量分析. 土壤通报, 2011, 42(2): 369- 371.
[30] 杨瑞秀. 甜瓜根系自毒物质在连作障碍中的化感作用及缓解机制研究[D]. 沈阳: 沈阳农业大学, 2014.
[31] Yu J Q, Matsui Y. Extraction and identification of phytotoxic substances accumulated in nutrient solution for the hydroponic culture of tomato. Soil Science and Plant Nutrition, 1993, 39(4): 691- 700.
[32] Yu J Q, Matsui Y. Phytotoxic substances in root exudates of cucumber (CucumissativusL.). Journal of Chemical Ecology, 1994, 20(1): 21- 31.
[33] 孙冰玉. 连作对烟田土壤酶活性、微生物种群数量及土壤理化性质的影响[D].哈尔滨: 东北林业大学, 2010.
[34] 韩春丽, 刘娟, 肖春华, 张旺锋, 刘梅, 黄建军. 新疆绿洲连作棉田土壤微量元素含量的时空变化研究. 土壤学报, 2010, 47(6): 1194- 1201.
[35] 华菊玲, 刘光荣, 黄劲松. 连作对芝麻根际土壤微生物群落的影响. 生态学报, 2012, 32(9): 2936- 2942.
[36] 郝慧荣, 李振方, 熊君, 陈慧, 张重义, 林文雄. 连作怀牛膝根际土壤微生物区系及酶活性的变化研究. 中国生态农业学报, 2008, 16(2): 307- 311.
[37] 张圣喜, 陈法霖, 郑华. 土壤微生物群落结构对中亚热带三种典型阔叶树种凋落物分解过程的响应. 生态学报, 2011, 31(11): 3020- 3026.
[38] Berendsen R L, Pieterse C M J, Bakker P A H M. The rhizosphere microbiome and plant health. Trends in Plant Science, 2012, 17(8): 478- 486.
[39] 王建花, 陈婷, 林文雄. 植物化感作用类型及其在农业中的应用. 中国生态农业学报, 2013, 21(10): 1173- 1183.
[40] Haichar F Z, Marol C, Berge O, Rangel-Castro J J, Prosser J I, Balesdent J, Heulin T, Achouak W. Plant host habitat and root exudates shape soil bacterial community structure. The ISME J, 2008, 2(12): 1221- 1230.
[41] Hartmann A, Schmid M, Van Tuinen D, Berg G. Plant-driven selection of microbes. Plant and Soil, 2009, 321(1/2): 235- 257.
[42] 覃逸明, 聂刘旺, 黄雨清, 王千, 刘欣, 周科. 凤丹(PaeoniaostiiT.)自毒物质的检测及其作用机制. 生态学报, 2009, 29(3): 1153- 1161.
[43] Zolla G, Badri D V, Bakker M G, Manter D K, Vivanco J M. Soil microbiomes vary in their ability to confer drought tolerance toArabidopsis. Applied Soil Ecology, 2013, 68: 1- 9.
AnalysisofphysicochemicalpropertiesandmicrobialdiversityinrhizospheresoilofAchyranthesbidentataunderdifferentcroppingyears
WANG Juanying1,2,3, XU Jiahui2,3, WU Linkun1,2,3, WU Hongmiao1,2,3, ZHU Quan1,2,3, KONG Lufei1,2,3, LIN Wenxiong1,2,3 *
1Collegeoflifescience,FujianAgricultureandForestryUniversity,Fuzhou350002,China2KeyLaboratoryofBiopesticideandChemicalBiology,MinistryofEducation,FujianAgricultureandForestryUniversity,Fuzhou350002,China3FujianProvincialKeyLaboratoryofAgroecologicalProcessingandSafetyMonitoring,Fuzhou350002,China
国家自然科学基金(81303170,81573530,31401950,U1205021);闽台作物特色种质创制与绿色栽培协同创新中心(2015- 75)
2016- 06- 07; < class="emphasis_bold">网络出版日期
日期:2016- 12- 27
10.5846/stxb201606071097
*通讯作者Corresponding author.E-mail: wenxiong181@163.com
王娟英,许佳慧, 吴林坤, 吴红淼, 朱铨, 孔露霏, 林文雄.不同连作年限怀牛膝根际土壤理化性质及微生物多样性.生态学报,2017,37(17):5621- 5629.
Wang J Y, Xu J H, Wu L K, Wu H M, Zhu Q, Kong L F, Lin W X.Analysis of physicochemical properties and microbial diversity in rhizosphere soil ofAchyranthesbidentataunder different cropping years.Acta Ecologica Sinica,2017,37(17):5621- 5629.
猜你喜欢
世界科学技术-中医药现代化(2022年9期)2023-01-17
中国土壤与肥料(2021年5期)2021-12-02
中成药(2018年12期)2018-12-29
中成药(2018年6期)2018-07-11
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
中学生阅读(初中版)(2016年11期)2017-01-13
中药与临床(2015年5期)2015-12-17
中国中医药现代远程教育(2014年11期)2014-08-08
中医研究(2014年4期)2014-03-11
