
金镶玉竹、黄秆乌哺鸡竹和黄纹竹在北京地区冬季的耐寒生理和叶片结构比较
2017-10-11 09:30:33国际竹藤中心北京000紫竹院公园北京00048
福建农林大学学报(自然科学版) 2017年5期
李 娟, 高 健, 范 蕊(.国际竹藤中心,北京 000;.紫竹院公园,北京 00048)
金镶玉竹、黄秆乌哺鸡竹和黄纹竹在北京地区冬季的耐寒生理和叶片结构比较
李 娟1, 高 健1, 范 蕊2
(1.国际竹藤中心,北京 100102;2.紫竹院公园,北京 100048)
对移栽到北京6 a的金镶玉竹、黄秆乌哺鸡竹和黄纹竹冬季越冬期的叶片结构、光合荧光特性、相对电导率等进行研究.结果表明:(1)3个竹种叶片上表皮无显著变化,下表皮黄秆乌哺鸡竹和黄纹竹的机械损伤程度大于金镶玉竹,黄秆乌哺鸡竹和黄纹竹气孔器宽度下降幅度显著小于金镶玉竹,黄纹竹叶片致密度、气孔密度下降幅度最大;(2)黄秆乌哺鸡竹净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)变化幅度大于金镶玉竹,而黄纹竹的却小于金镶玉竹,Fv/Fm值降幅表现为金镶玉竹<黄秆乌哺鸡竹<黄纹竹;(3)金镶玉竹和黄秆乌哺鸡竹叶片的相对电导率变化幅度差异不显著,黄纹竹的叶片和鞭根相对电导率的变化幅度差异显著.
金镶玉竹; 黄秆乌哺鸡竹; 黄纹竹; 冬季; 北京
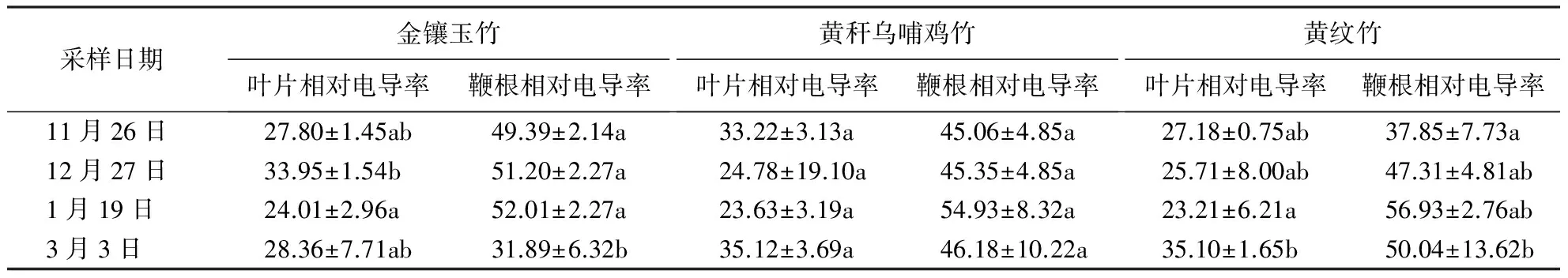
Abstract: For wide application of bamboo species, growth mechanism of 3 kinds of bamboo planted in Beijing, includingPhyllostachysaureosulacataf.spectabilis, Ph. vivaxf. aureocaulis,Ph.vivaxf.huangwenzhu, was compared in terms of leaf ultrastructure, photosynthetic characteristics and fluorescence, electric conductivity after winter. The results showed that there were no significant variations in upperepidermis among 3 species, but physical damages were more serious in underepidermis ofPh.vivaxf.aureocaulis, Ph. vivaxf.huangwenzhuthanPhyllostachysaureosulacataf.spectabilis.Fluctuation in stomatal width was smaller inPh.vivaxf.aureocaulisandPh.vivaxf.huangwenzhuthanPhyllostachysaureosulacataf.spectabilis. Compared withPhyllostachysaureosulacataf.spectabilis, net photosynthetic rate (Pn) and transpiration rate (Tr) stomatal conductance (Gs) varied more significantly inPh.vivaxf.aureocaulis, but less inPh.vivaxf.huangwenzhu. The decline rate ofFv/Fmin an ascending order wasPhyllostachysaureosulacataf.spectabilis Keywords:Phyllostachysaureosulacataf.spectabilis;Ph.vivaxf.aureocaulis;Ph.vivaxf.huangwenzhu; Beijing 温度是限制植物分布的主要因素,温度过高或过低对植物的正常生长均产生不利影响,其中低温对植物的危害最大.多年生竹亚科植物主要分布在长江以南地区,北方地区分布数量少,分布面积小[1].从上世纪80年代开始,山东、河北、北京和陕西等地对部分观赏价值高的竹种进行耐寒选育和引种工作[2-11],研究结果显示:刚竹属的早竹(Ph.praecox)、黄秆京竹(Ph.aureosulacataf.aureocaulis)、黄秆乌哺鸡竹(Ph.vivaxf.aureocaulis)、黄纹竹(Ph.vivaxf.huangwenzhu)等竹种,地被竹铺地竹(Sasaargenteastriatus)、菲白竹(S.fortunei)等能忍受-10 ℃以下的低温,在引种地安全越冬并生长良好.近年来,学者对自然低温条件下黄秆乌哺鸡、黄条金刚竹(Pleioblastuskongosanensis)、麻竹(Dendrocalamuslatiflorus)等竹种的生理指标和形态指标进行研究,结果[12-15]显示乔木状竹种耐寒性高于灌木状竹种,耐寒竹种在低温胁迫下通过可溶性蛋白和可溶性糖的升高来提高细胞的保水能力,同时维持较高的酶活性以减轻低温下膜脂过氧化水平.黄秆乌哺鸡竹、黄纹竹属于刚竹属,是乌哺鸡竹的2个栽培变型,主要分布在江苏和浙江等地,是不可多得的观赏及笋用竹种[1].对这2个竹种进行引种研究,结果显示2个竹种均是较耐寒的品种[16-21].金镶玉竹原产北京、江苏[1]等地,耐寒性高.本研究将3个竹种在在北京越冬期的多个指标进行综合比较,探讨3个竹种的耐寒机制,为耐寒竹种在北方地区种质繁育和竹类植物在北方地区的推广应用研究提供参考. 试验地位于北京市房山区良乡,地处平原地带,年平均气温11.6 ℃,最高气温达43.5 ℃,最低气温达到-26 ℃;年平均降水量687 mm,最大降水量1 322 mm,最小降水量277 mm;年平均无霜期185 d,土壤以黄棕壤为主.供试竹种为金镶玉竹、黄秆乌哺鸡竹和黄纹竹,于2004年由南京林业大学竹种园母竹移栽引入,每个竹种的引种面积均为25 m2,于国际竹藤中心良乡试验站种植6 a,所处环境一致且相邻,地形平坦. Kestrel 4000小气候仪由Nielsen-Kellerman公司提供;Li-COR6400便携式光合仪由Li-COR公司提供;IMAGR-PAM便携式调制叶绿素荧光仪由Walz公司提供;DDSJ-308A电导仪由上海仪电科学仪器有限公司提供;Free Zone冷冻干燥机由Labconco公司提供;Quanta 200HV环境扫描电镜由Philips公司提供. 利用小气候仪观测并记录竹林每日的最高、最低气温.分别在11月26日,12月27日,1月19日和3月3日上午、中午、下午测定小气候的温度、湿度、风速等,每次测定3次,结果取平均值.采用直角地温计测定竹种地下10、20、40 cm的温度,测定3个重复,结果取平均值. 选取1年生竹枝上部受光较一致的功能叶3~5片进行标记,于9:00—11:30分别利用便携式光合仪和便携式调制叶绿素荧光仪测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和PSⅡ最大光化学效率(Fv/Fm).测定时控制叶室温度为25 ℃,光强为800 μmol·m-2·s-1,参比室CO2浓度为380 mol·L-1.测定3株重复植株,每株选取叶片3~5片,测定结果取平均值. 分别在11月26日、12月27日、1月19日和3月3日上午10:00—12:00,从长势良好的植株冠层中部的向阳面剪取1年生竹叶,地下20 cm 挖取鞭根,取回实验室分析其相对电导率和竹叶超微结构.用自来水洗净叶片表面,再用去离子水冲洗2次.将3个竹种叶片和鞭根均称取5 g,切成小块,放入洁净10 mL的试管中,加入去离子水,于25 ℃浸泡1 h,其间多次摇动试管.利用电导仪测定浸出液相对电导率(S1),测毕,将试管盖塞封口,置沸水浴30 min,以杀死植物组织;冷却至室温后平衡10 min,摇匀测绝对电导率(S2),并计算相对电导率.相对电导率/%=(S1/S2)×100.每个竹种测定3次. 竹叶经冷冻干燥机冷冻抽干48 h,以保证材料解剖结构的稳定性.将竹叶用导电胶粘于样品台上进行喷金处理,然后置于环境扫描电镜中观测竹叶超微结构. 利用Digiminizr软件测量竹叶超微结构的气孔器长度和气孔器宽度,采用Excel 2007和SPSS 14.0软件对数据进行统计分析.采用单因素(one-way ANOVA)和LSD法进行方差分析和多重比较(α=0.05),数据为平均值±标准差.不同耐寒指标综合比较采用模糊数学隶属函数法. 从2010年11月26日越冬初期(以后简称“初期”)到2011年3月3日越冬末期(以后简称“末期”)的气温变化情况,如图1所示.从11月到翌年3月,最低温为-14 ℃(2011年1月14日);1月份温度最低,平均气温为-9.06~-0.3 ℃.2010年12月温度波动较大,温度3次降到-10 ℃左右;2011年2月测得的温度仍然较低,最低温度仍然保持在-5 ℃左右,3月份后,气温逐渐升高. 图1 2010年11月—2011年3月北京市房山区的最高温度、最低温度变化Fig.1 Changes in the highest and lowest temperatures from November 2010 to March 2011 in Fangshan district in Beijing 从表1可以看出,从初期到末期,研究区竹种小气候温度与该区域一致.测定日期11月26日和3月3日,温度在0 ℃以上;12月27日和1月19日温度在0 ℃以下,差异显著.11月26日湿度较低,显著低于1月19日和3月3日这2个测定日.研究地在测定日风速均较小,差异不显著.同时根据观察,研究地在冬季1—2月,有风日占98%,白天平均风速1.18 m·s-1,最大风速3.4 m·s-1.地下温度比地上温度高,越冬期地下温度均在0 ℃以上,11月26日地下10 cm温度比地下40 cm高1.17 ℃.其他3个测定日则相反,土层越深温度越高,4个测定日3个土层温度差异均不显著. 表1 研究地越冬期小气候和地下温度的变化1)Table 1 Changes in microclimate and subsurface temperature in experimental plot during wintering 1)相同小写字母表示差异不显著,不同小写定母表示差异显著(P<0.05). 如图2所示,金镶玉竹、黄秆乌哺鸡竹、黄纹竹的叶片结构相似,上表皮平整,基本没有附属结构,可以分辨脉带与脉间带.上表皮(近轴面)由长细胞、短细胞、硅质细胞及少量气孔器组成.下表皮(远轴面)由脉间带和脉带组成,分布有乳突、气孔器、微毛和刺毛(少量).金镶玉竹脉间带微毛分布较少,其他2个竹种分布较多.气孔器主要分布在下表皮,分散排列较为均匀,分布方向与叶长轴一致,由副卫细胞和保卫细胞共同组成.经过越冬期,2个竹种上表皮无明显变化,下表皮发生机械损伤,细胞壁细胞膜系统受破坏,黄纹竹损伤大于其他2个竹种. 如表2所示,3个竹种经过越冬期后,叶片末期与初期相比,上表皮无显著变化,下表皮乳突密度、微毛密度、气孔密度、气孔器长度、气孔器宽度均下降.金镶玉竹、黄秆乌哺鸡竹和黄纹竹乳突密度分别下降了53.11%、44.33%、66.90%,微毛密度分别下降了43.21%、41.92%、48.12%,气孔密度分别下降了15.56%、18.60%、72.18%.末期与初期相比,黄纹竹的变化差异显著,说明黄纹竹机械损伤较严重.金镶玉竹气孔器长度和宽度在初期均较大,经过越冬期,其气孔器长度和气孔器宽度分别下降了56.31%和76.54%;而黄秆乌哺鸡竹和黄纹竹的气孔器长度分别下降了61.13%、56.78%;宽度分别下降了8.33%、17.87%.金镶玉竹气孔器宽度变化差异显著. A.11月26日金镶玉竹上表皮;B.3月3日金镶玉竹上表皮;C.11月26日金镶玉竹下表皮;D.3月3日金镶玉竹下表皮;E.11月26日黄秆乌哺鸡竹上表皮;F.3月3日黄秆乌哺鸡竹上表皮;G.11月26日黄秆乌哺鸡竹下表皮;H.3月3日黄秆乌哺鸡下表皮;I.11月26日黄纹竹上表皮;J.3月3日黄纹竹上表皮;K.11月26日黄纹竹下表皮;L.3月3日黄纹竹下表皮.图2 3个竹种越冬期叶片超微结构Fig.2 Changes in leaf ultrastructure for 3 bamboo species during wintering 表2 3个竹种越冬期叶片结构特征1)Table 2 Changes in leaf structure in 3 bamboo species during wintering 1)相同小写字母表示差异不显著,不同小写定母表示差异显著(P<0.05). 如图3所示,越冬末期3个竹种的光合指标与初期相比均发生较大变化.3个竹种在初期Pn值为正值,且差异显著;末期降为负值,差异不显著,金镶玉竹、黄秆乌哺鸡竹和黄纹竹的降幅分别为126.10%、174.07%和144.12%.3个竹种Tr值在初期为负值,金镶玉竹Tr与黄纹竹差异不显著,两者Tr均与黄秆乌哺鸡竹差异显著.末期温度升高,太阳辐射使得Tr值升为正值,金镶玉竹、黄秆乌哺鸡竹和黄纹竹Tr的增幅分别是104%、207%和120%.3个竹种气孔导度Gs在初期为负值.初期金镶玉竹Gs与黄纹竹差异不显著,两者Gs均与黄秆乌哺鸡竹差异显著.末期Gs为正值,黄秆乌哺鸡竹和黄纹竹在末期Gs显著增大,增幅分别是272.40%和329.68%,两者差异不显著,而金镶玉的增幅仅211.36%.3个竹种PSⅡ最大光化学效率(Fv/Fm)值在初期差异不显著,但在越冬末期,降幅分别是17.63%、19.88%、27.07%,金镶玉竹Fv/Fm显著低于其他2个竹种. Ⅰ.金镶玉竹;Ⅱ.黄杆乌哺鸡竹;Ⅲ.黄纹竹.图3 3个竹种越冬期光合荧光特性的变化Fig.3 Changes in photosynthetic characteristics and fluorescence for 3 bamboo species during witering 如表3所示,末期叶片相对电导率与初期相比,金镶玉竹、黄秆乌哺鸡竹和黄纹竹升高了2.00%、5.72%和29.14%;金镶玉竹与黄秆乌哺鸡竹在越冬期不同测定时间变化差异均不显著;黄纹竹在末期与其他测定时间差异显著.3个竹种初期鞭根相对电导率大于叶片相对电导率,末期金镶玉鞭根相对电导率下降了35.43%,而黄秆乌哺鸡竹和黄纹竹的相对电导率分别提高了29.14%和32.21%;末期与初期相比,金镶玉竹与黄纹竹差异显著,与黄秆乌哺鸡竹差异不显著. 通过计算3个竹种的乳突密度、微毛密度、气孔器长度、气孔器宽度、气孔密度、Fv/Fm、Pn、Tr、Gs和相对电导率10个指标隶属函数值,评价研究竹种的耐寒性,结果表明金镶玉竹的耐寒性隶属函数指标中乳突密度、气孔器宽度、光合相关指标均高于其它2个竹种,计算得到的耐寒性隶属函数均值表现为金镶玉竹(0.51)>黄秆乌哺鸡竹(0.49)>黄纹竹(0.38). 表3 3个竹种越冬期相对电导率的变化1)Table 3 Changes in electric conductivity in 3 bamboo species during wintering % 1)相同小写字母表示差异不显著,不同小写字母表示差异显著(P<0.05). 研究结果显示,耐寒性表现为金镶玉竹>黄秆乌哺鸡竹>黄纹竹,黄秆乌哺鸡竹和黄纹竹能在北京地区安全越冬,这与他人研究结果[8-9,11,16-17, 20-21]一致,但仅对3个竹种在寒冷胁迫下外观形态和少部分生理指标进行比较,对3个竹种的耐寒机制未作深入分析.本文通过分析发现,金镶玉竹叶片结构致密,气孔调节能力强于其他2个竹种;黄纹竹在越冬期光合能力下降,叶片冻伤严重,耐寒能力较弱. 金镶玉竹叶片组织结构紧密度相对较大,表皮乳突密度大于其他2个竹种,气孔密度偏小,这种结构可以减少因冻害导致细胞收缩产生的机械损伤[22].金镶玉的Pn、Tr降幅最小,推测其表面结构降低了其与外界环境进行气体交换的能力[22],有助于防止水分过度蒸散[22-23].金镶玉竹在越冬期结束后气孔宽度下降最大.说明其气孔调节能力较强,防止水分的过分蒸腾,同时提高水分调节能力[24],使其对环境具有较强的适应能力.黄秆乌哺鸡竹和黄纹竹叶片组织结构紧密度小于金镶玉竹,胁迫初期2个竹种乳突密度、微毛密度、气孔密度差异不显著,末期黄纹竹降幅大于黄秆乌哺鸡竹.说明黄纹竹因冻害产生的机械损伤大于黄秆乌哺鸡竹.黄秆乌哺鸡竹Pn、Tr变幅最大,但Fv/Fm降幅小于黄纹竹,表明其在低温逆境中通过降低光合速率、呼吸消耗和调整Gs提高恢复能力[25],增强植株抗逆性.黄纹竹气孔密度较大,在初期光合能力强于金镶玉竹,但是其组织调节能力弱于其他2个竹种.末期黄纹竹的Gs增幅最大,同时Fv/Fm值降幅最大,机械损伤比其他2个竹种严重. 在整个越冬期,3个竹种叶片相对电导率和鞭根相对电导率均出现上升、下降反复变化的过程,说明各竹种在遭遇低温胁迫时,其体内的防御机制慢慢发生作用,从而提高其自我保护能力[26-27].在寒害低温下,耐寒性强的植物,膜透性增加较慢且变化可逆转,易恢复正常;反之,耐寒性弱的植物膜透性增大,速度较快且不可逆转,常不能恢复正常,从而导致死亡[28].末期与初期相比,黄纹竹叶片相对电导率和鞭根相对电导率均显著增强,表明其受到的冻害大于其他2个竹种,这个结果也验证了上述3个竹种叶片结构变化的结果. 本研究3个竹种在北京地区适宜的小环境均可生长较好,如果遇到寒冷年份,应提前对黄纹竹进行修剪或适宜越冬保护.由于本研究母竹移栽自南京林业大学竹种园,然而在不同生长环境选择不同的种源十分重要,需要进一步研究.同时,有学者认为不耐寒的竹种的原因是在低温条件下其导管中产生严重的气穴栓塞[29],说明竹类植物耐寒性是受多种因素的影响,有待今后进一步研究. [1] 耿伯介,王正平.中国植物志(第九卷,第一分册)[M].北京:科学出版社,1996. [2] 孙守家.山东蒙山竹类引种试验初报[J].山东林业科技,2002(4):8-11. [3] 张玉龙,吴建功,马荣田.淡竹引种栽培试验初报[J].山西林业科技,2003(1):44-45. [4] 卢义山,李荣锦,倪竞德.竹类植物种质资源圃抗寒性竹种的选择[J].江苏林业科技,2007,34(2):12-17. [5] 李作军.6种观赏竹在关中地区引种实验[D].杨凌:西北农林科技大学,2013. [6] 刘国华,邹秀华,王福升,等.山东省引种栽培竹种生长情况调查及对比研究[J].山东农业大学学报(自然科学版),2006,37(4):586-590. [7] 刘西岭,刘汉柱,辛华.青岛引种的5个竹种与当地竹种的抗寒性比较研究[J].中国农学通报,2010,26(8):129-131. [8] 王金革,陈进勇.北京植物园竹亚科植物耐寒性评价[J].世界竹藤通讯,2012,10(2):1-8. [9] 赵康,冯小虎,欧小平,等.北京地区竹类引种试验初报[J].世界竹藤通讯,2006,4(4):15-19. [10] 张玲.引种地被竹抗旱抗寒性研究[D].泰山:山东农业大学,2009. [11] 范卓敏,冯小虎,翟敬宇.观赏竹在北京地区园林绿化中应用的调查与分析[J].世界竹藤通讯,2011,9(3):1-7. [12] 刘国华,栾以玲,张艳华.自然状态下竹子的抗寒性研究[J].竹子研究汇刊,2006,25(2):10-14. [13] 林树燕,丁雨龙.电导法对7 种观赏竹的抗寒性测定[J].西北林学院学报,2008,23(1):34-38. [14] 张玮,黄树燕,吴继林,等.低温胁迫对麻竹叶片和根系抗性生理指标的影响[J].生态学杂志,2012,31(3):513-519. [15] 高贵宾,钟浩,吴志庄,等.不同混生地被竹光合和荧光特征比较[J].福建农林大学学报(自然科学版),2016,45(5):515-521. [16] 郑蓉,黄耀华,连巧霞,等.刚竹属13个竹种叶绿素荧光特性比较[J].江西农业大学学报,2008,30(4):263-267. [17] 孙飞飞.观赏竹的耐寒性研究及园林应用[D].临安:浙江农林大学,2015. [18] 胡爱群,黄术,张胜华.黄秆乌哺鸡竹引种研究[J], 湖南林业科技,2015,42(5):83-86. [19] 周成敏,宋艳冬,何林.黄纹竹驯化试验[J].浙江林业科技,2016,36(1):64-67. [20] 孙耀清,李文杨,张桂玲.引种观赏竹生态适应性研究[J].河南农业科学,2015,44(5):125-128. [21] 孙艳.百竹园26种竹种生态适应性初步研究[D].福州:福建农林大学,2014. [22] 刘国华,林树燕,王福升,等.4种地被竹生理指标对低温的响应[J].竹子研究汇刊,2011,30(4):6-10. [23] 杨明博,杨吉力,杨九艳,等.鄂尔多斯高原不同生境条件下中间锦鸡儿植物叶片表皮特征及遗传多样性变化分析[J].植物生态学报,2007,31(6):1 181-1 189. [24] WOOLEY J T. Water relations of soybean leaf haiers[J]. Agronomy Journal, 1964,56(6):569-571. [25] 武辉,戴海芳,张巨松,等.棉花幼苗叶片光合特性对低温胁迫及恢复处理的响应[J].植物生态学报,2014,38(10):1 124-1 134. [26] 李迎春,陈双林,郭子武,等,模拟低温或自然低温条件下牡竹属3种类抗寒性及生理指标的比较[J].植物资源与环境学报,2012,21(3):13-19. [27] 秦宇.山东省引种观赏竹的抗寒性研究[D].泰山:山东农业大学,2014. [28] CHAN H J. Electrolyte leakage and ethylene production induced by chilling injury of papayas[J]. Hmtscience, 1985,2(6):1 070-1 072. [29] 王福升,田新立,丁雨龙,等.用根压法研究竹子的耐旱、耐寒性[J].林业科学,2011,47(8):176-181. (责任编辑:叶济蓉) ChangesinphysiologicalindicesandleafstructureofPhyllostachysaureosulacataf.spectabilis,Ph.vivaxf.aureocaulis,Ph.vivaxf.huangwenzhuduringwinterinBeijing LI Juan1, GAO Jian1, FAN Rui2 S718.51 A 1671-5470(2017)05-0527-07 10.13323/j.cnki.j.fafu(nat.sci.).2017.05.008 2017-02-13 2017-05-02 国际竹藤中心基本科研业务费资助项目(1632017017);国家科技支撑计划项目(2015BAD04B03). 李娟(1981-) ,女,助理研究员.研究方向:植物生理生态.Email:ljgx2003@126.com.1 材料与方法
1.1 试验地概况
1.2 主要仪器
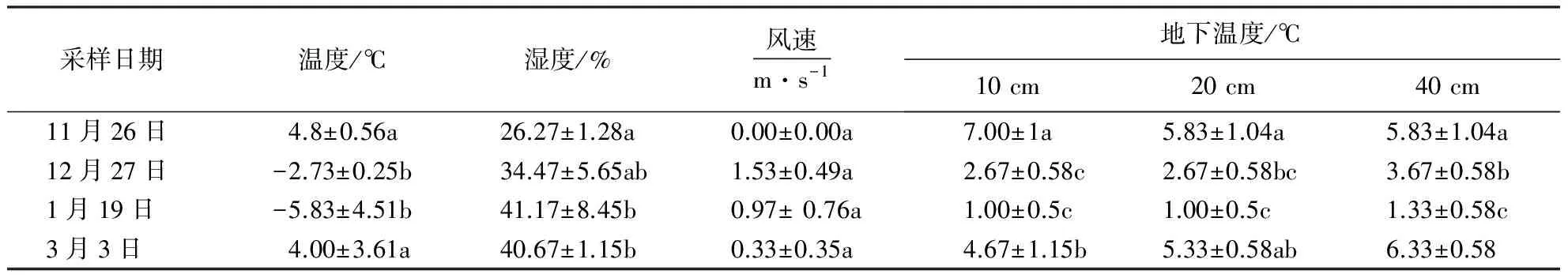
1.3 小气候测定
1.4 光合、叶绿素荧光参数测定
1.5 相对电导率和超微结构测定
1.6 数据分析
2 结果与分析
2.1 研究地小气候变化
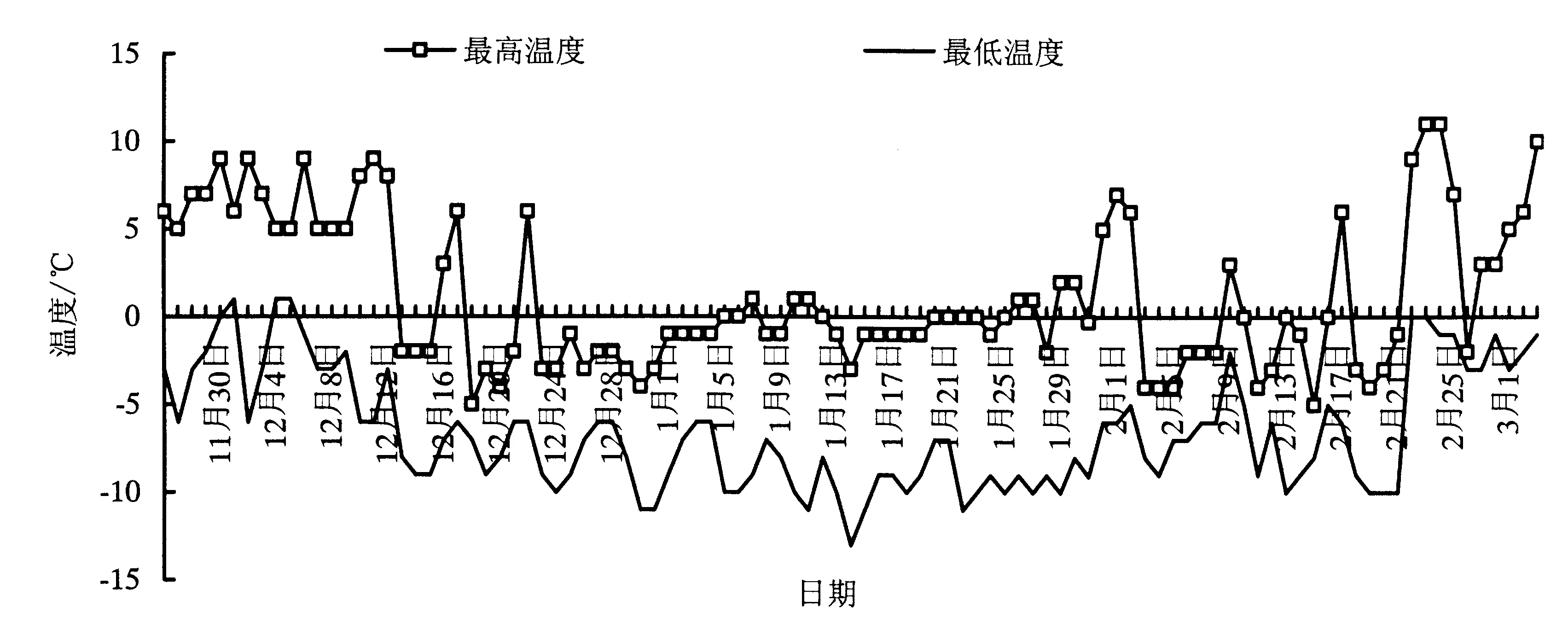
2.2 竹种叶片超微结构

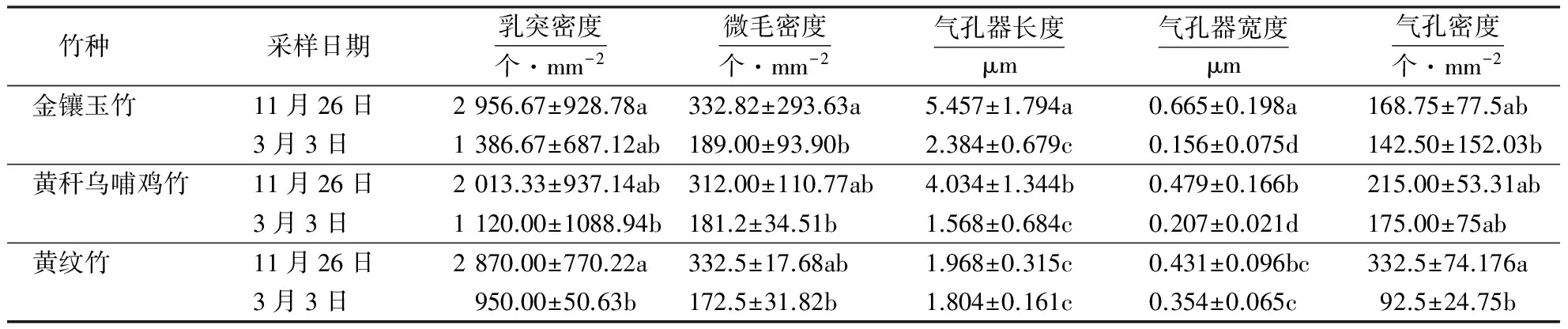
2.3 竹种光合荧光特性
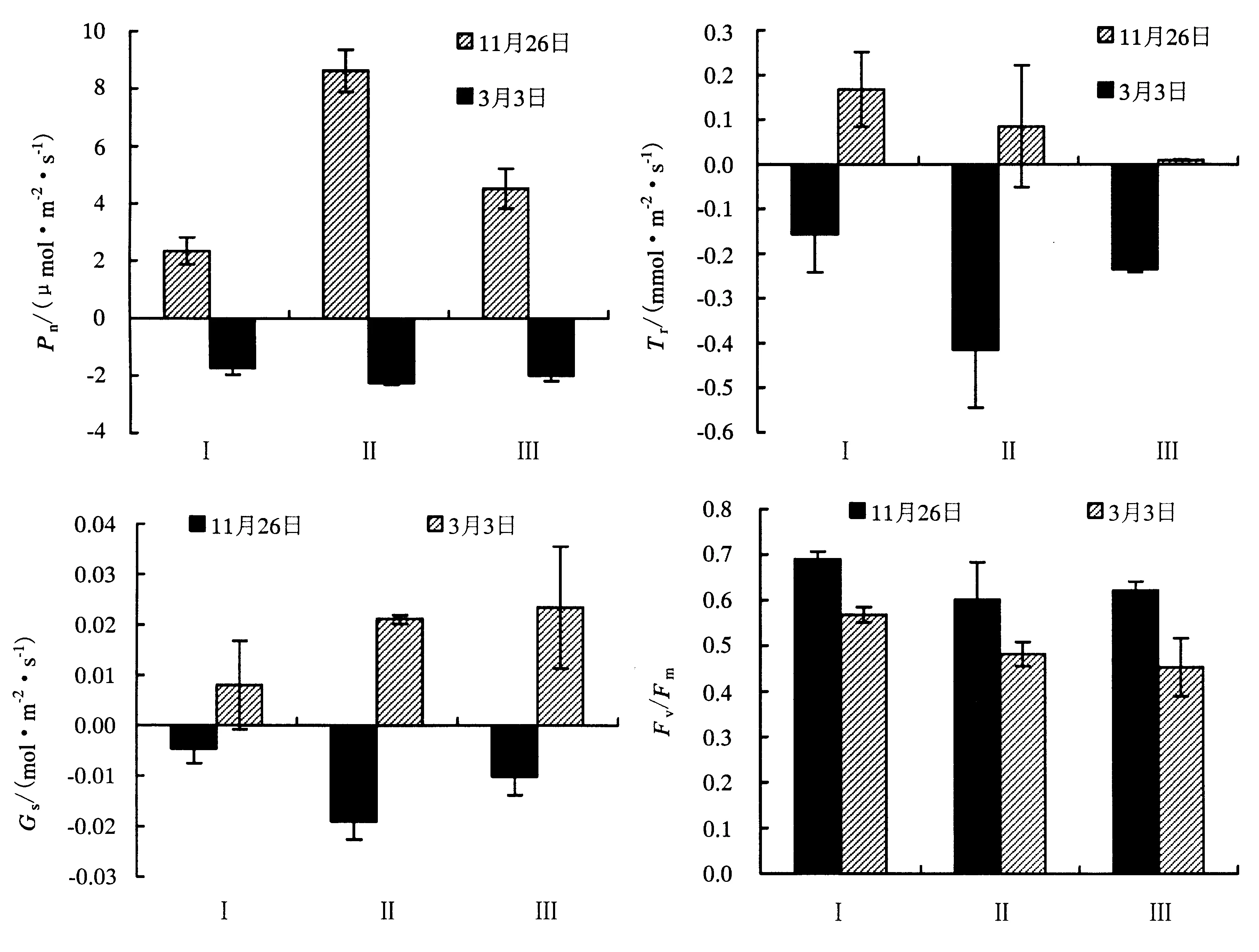
2.4 竹种相对电导率的变化
2.5 竹种耐寒性综合评价
3 小结与讨论
(1.International Centre for Bamboo and Rattan, Beijing 100102, China; 2.Zizhuyuan Park, Beijing 100048, China)
猜你喜欢
广东农业科学(2016年8期)2016-10-08 05:05:19
中国宝玉石(2016年1期)2016-09-29 04:00:44
农业与技术(2015年24期)2016-01-26 03:30:27
世界竹藤通讯(2015年4期)2015-12-29 10:46:38
世界竹藤通讯(2015年4期)2015-12-29 10:46:36
西部资源(2015年4期)2015-02-15 02:19:42
作物研究(2014年6期)2014-03-01 03:38:59
世界竹藤通讯(2010年2期)2010-08-15 00:50:49
世界竹藤通讯(2010年1期)2010-05-30 07:26:02
中学生英语·阅读与写作(2008年8期)2008-12-19 05:28:50
