
赖氨酸对奶牛乳腺上皮细胞内乳脂肪合成相关基因和蛋白表达的影响
2017-09-16 05:59赵艳丽郭晓宇史彬林闫素梅
动物营养学报 2017年9期
陈 璐 赵艳丽 郭晓宇 史彬林 闫素梅
(内蒙古农业大学动物科学学院,呼和浩特010018)
赖氨酸对奶牛乳腺上皮细胞内乳脂肪合成相关基因和蛋白表达的影响
陈 璐 赵艳丽 郭晓宇 史彬林 闫素梅*
(内蒙古农业大学动物科学学院,呼和浩特010018)
本试验旨在研究赖氨酸(Lys)对奶牛乳腺上皮细胞(BMECs)内乳脂肪合成相关基因和蛋白表达的影响,探讨Lys影响乳脂肪合成的机理。将第3代BMECs随机分为6组,每组6个重复,每个重复1个培养孔。各组培养基中Lys的浓度分别为0.5(基础培养基,对照)、1.0、2.0、4.0、8.0和16.0 mmol/L,37 ℃、5% CO2培养48 h后测定BMECs甘油三酯(TAG)含量、乳脂肪合成相关基因和蛋白的表达量。结果表明:BMECs内TAG含量(P=0.013)以及脂肪酸结合蛋白3(FABP3,P=0.001)、脂蛋白脂酶(LPL,P=0.096)、脂肪酸合成酶(FASN,P=0.003)、乙酰甘油磷酸脂酰转移酶6(AGPAT6,P=0.038)和甘油-3-磷酸酰基转移酶(GPAM,P=0.022)基因表达量对Lys呈显著或趋于显著的浓度依赖效应。FABP3基因表达量以2.0、4.0、8.0、16.0 mmol/L组和LPL基因表达量以1.0、2.0、4.0、8.0、16.0 mmol/L组显著高于0.5 mmol/L组(P<0.05);FASN基因表达量以2.0 mmol/L组最高,显著高于16.0 mmol/L组(P<0.05);硬脂酰辅酶A去饱和酶1(SCD1)基因表达量以2.0、4.0 mmol/L组显著高于其他组(P<0.05);磷脂酸磷酸酯酶1(LPIN1)、嗜乳脂蛋白亚家族1成员1(BTN1A1)和黄嘌呤脱氢酶(XDH)基因表达量均以1.0、2.0、4.0、8.0 mmol/L组显著高于0.5 mmol/L组(P<0.05);过氧化物酶体增殖物激活受体γ(PPARγ)基因及蛋白表达量均以2.0、4.0 mmol/L组显著高于0.5和8.0、16.0 mmol/L组(P<0.05);固醇调节元件结合蛋白1(SREBP1)基因表达量以1.0、2.0、4.0 mmol/L组显著高于其他组(P<0.05),蛋白表达量以1.0 mmol/L组显著高于其他组(P<0.05)。但高浓度Lys抑制AGPAT6和GPAM的基因表达,AGPAT6基因表达量以2.0、4.0、8.0、16.0 mmol/L组显著低于0.5、1.0 mmol/L组(P<0.05),GPAM基因表达量以16.0 mmol/L组显著低于0.5、1.0、2.0、4.0 mmol/L组(P<0.05)。可见,Lys对BMECs的乳脂肪合成具有显著的促进效果,但高浓度的Lys抑制了乳脂肪合成相关基因的表达。本试验条件下,培养基中Lys适宜浓度为2.0~4.0 mmol/L。
奶牛;乳腺上皮细胞;赖氨酸;乳脂肪
乳脂肪是牛奶的主要成分,是评价牛奶质量的重要指标之一。氨基酸(AA)是乳蛋白合成的重要前体物,但AA在影响乳蛋白合成的同时,也影响着乳脂肪合成与组成[1]。因此,深入研究AA对乳脂肪合成的影响及机理,对调控乳腺内乳成分的合成和改善乳品质具有重要意义。赖氨酸(Lys)是动物的必需氨基酸,Giallongo等[2]向奶牛灌注过瘤胃赖氨酸(RPLys)后发现,Lys在促进乳蛋白合成的同时,对乳脂肪的合成具有促进效果。Xu等[3]研究发现,向泌乳母猪饲喂Lys和缬氨酸(Val)后,适宜的Lys和Val比例能够使母猪背部脂肪损失降到最低,说明Lys和Val能够影响乳脂肪的合成。韩慧娜[4]研究发现,秸秆饲粮条件下奶牛阴外动脉灌注AA能促进乳腺内短链脂肪酸的摄取。可见,Lys在一定程度上影响了乳脂肪的合成,然而国内外的研究报道多集中在向奶牛瘤胃内及阴外动脉灌注Lys影响乳蛋白和乳脂肪合成的方面,但以奶牛乳腺上皮细胞(BMECs)为模型,研究不同浓度Lys对乳脂肪合成相关基因和蛋白表达的影响的国内外报道较少,此方面机理有必要继续进行研究和探索。鉴于此,本研究以BMECs为模型,研究不同浓度Lys对乳脂肪合成相关基因和蛋白表达的影响,为进一步探讨Lys对BMECs内乳脂肪合成的影响机理提供理论基础,为调控奶牛Lys营养水平、提高生产性能提供理论依据。
1 材料与方法
1.1主要试剂与仪器
Ⅱ型胶原酶、DMEM/F12培养基、胰岛素转铁蛋白硒钠、胎牛血清(FBS)、0.25%胰蛋白酶/乙二胺四乙酸(EDTA)购自Gibco公司;Lys(L8662)、氢化可的松、表皮生长因子、催乳素、油红O、琼脂糖和兔抗过氧化物酶体增殖物激活受体γ(PPARγ)抗体购自Sigma公司(AV32880);两性霉素购自Amresco公司;细胞培养用双抗购自Corning公司;磷酸盐缓冲液(PBS)和Tris缓冲液(TBS)购自HyClone公司;RNAiso PLUS、PrimeScript RT Master Mix和SYBR Premix ExTaqTMⅡ购自TaKaRa公司;放射免疫沉淀测定(RIPA)裂解液、2,2-联喹啉-4,4-二甲酸二钠(BCA)蛋白质浓度试剂盒、Western一抗稀释液、Western二抗稀释液、十二烷基硫酸钠(SDS)-聚丙烯酰胺凝胶电泳(PAGE)电泳液、Western转膜液、ECL化学超敏显色液购自北京碧云天公司;鼠抗固醇调节元件结合蛋白1(SREBP1)抗体购自Abcam公司(ab3259);兔抗磷酸甘油醛脱氢酶(GAPDH)抗体购自Proteintech公司(10494-1-AP);辣根过氧化物酶(HRP)标记山羊抗兔二抗购自KPL公司(04-15-06);HRP标记山羊抗鼠二抗购自天津三箭生物技术有限公司(LK2003)。
主要仪器包括:二氧化碳恒温培养(Forma-311,Thermo公司);生物安全柜(MSC-ADVANTAGE,Thermo公司);倒置显微镜(Olympuse公司);全自动酶标仪(Synergy H4 BioTek,BioTek公司);细胞计数仪(Cytorecon,GE公司);荧光定量PCR仪(ABI-7500,ABI公司);蛋白质电泳仪、蛋白质转膜仪、蛋白质成像系统(Bio-Rad公司)。
1.2原代BMECs体外培养
BMECs用胶原酶消化法获得。在内蒙古呼和浩特市北亚清真屠宰场选取3~5岁经产的健康泌乳中期的高产荷斯坦奶牛乳腺组织。去除组织表层,于深层取约1 cm3的组织块放入4 ℃预冷的3×双抗PBS中。随后在超净台内分别用3×PBS、75%酒精和1×PBS清洗。再去除组织块的表层,选腺泡丰富的组织剪成糊状。加入等体积0.5%的Ⅱ型胶原酶溶液,37 ℃和5% CO2条件下消化1 h。之后用80目滤网过滤,收集细胞滤液,179×g离心5 min,弃上清;PBS冲洗细胞,179×g离心3 min,重复冲洗2次。加入完全培养基,吹打均匀后接种于25 cm2细胞培养瓶中,在37 ℃和5% CO2条件下培养。当原代细胞贴壁率达到80%~90%后用0.25%胰蛋白酶/EDTA对细胞进行纯化和传代[5]。
1.3试验设计
试验用第3代的BMECs,按照试验要求的细胞密度接种于不同的细胞培养板上,在37 ℃和5% CO2条件下培养24 h。采用单因子随机试验设计,试验中使用的DMEM/F12培养基中Lys的浓度为0.5 mmol/L,参考李喜艳[6]和高海娜[7]的研究结果并通过噻唑蓝(MTT)法测定细胞的增殖率确定Lys的浓度,各组Lys的浓度分别为0.5(基础培养基,对照)、1.0、2.0、4.0、8.0和16.0 mmol/L,每组6个重复,每个重复1个培养孔。细胞贴壁率达到80%~90%时,换为饥饿培养基,培养12 h后按照试验设计要求换成不同浓度Lys的培养基,37 ℃和5% CO2培养48 h。
1.4测试指标与方法
1.4.1 细胞活力与TAG的含量
细胞活力采用MTT法检测,以490 nm吸光度值(OD490 nm)计算细胞相对增殖率(RGR),公式如下:
RGR(%)=(试验组OD490 nm/对照组OD490 nm)×100。
TAG含量参考Ramírez-Zacarías等[8]的方法测定,将细胞以5×104个/mL的密度接种于24孔培养板上,按试验设计培养结束后,弃掉培养基,PBS清洗2遍,每孔加入4%多聚甲醛溶液0.2 mL固定细胞1 h,PBS清洗2遍,用0.5 mL油红O工作液避光浸染2 h。用PBS清洗3遍,晾干培养板后,加入0.3 mL异丙醇萃取30 min,用全自动酶标仪测定波长为510 nm吸光度值(OD510 nm),用以表示TAG含量。
1.4.2 BMECs内乳脂肪合成相关基因的表达
将细胞以2×105个/mL的密度接种于6孔培养板,按试验设计培养结束后,细胞总RNA按照Trizol法提取,用酶标仪检测总RNA的纯度与浓度,260和280 nm吸光度值比值(OD260 nm/OD280 nm)在1.8~2.2表示总RNA纯度较好。完整性用2%琼脂糖凝胶电泳来检测。将总RNA反转录成cDNA,采用PrimeScript RT Master Mix试剂盒的方法,反转录体系为10 μL。基因表达量采用SYBY Premix Ex TaqTMⅡ试剂盒的方法进行测定,反应体系为20 μL。以GAPDH为管家基因,对乳脂肪合成相关基因[脂肪酸合成酶(FASN)、乙酰辅酶A羧化酶(ACACA)、PPARγ、SREBP1、硬脂酰辅酶A去饱和酶1(SCD1)、脂肪酸结合蛋白3(FABP3)、脂蛋白脂酶(LPL)、乙酰甘油磷酸脂酰转移酶6(AGPAT6)、甘油-3-磷酸酰基转移酶(GPAM)、磷脂酸磷酸酯酶1(LPIN1)、嗜乳脂蛋白亚家族1成员1(BTN1A1)、黄嘌呤脱氢酶(XDH)]进行相对定量,引物序列见表1。实时荧光定量PCR的反应程序为:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火34 s,72 ℃延伸20 s,进行40个循环反应;95 ℃ 5 s,60 ℃ 30 s,95 ℃ 15 s,51个循环,绘制熔解曲线。基因的相对表达量采用2-△△Ct表示。

表1 乳脂肪合成相关基因的引物序列

续表1基因GenesGenBank登录号GenBankaccessionNo.引物序列Primersequences(5'—3')长度Length/bp参考文献References嗜乳脂蛋白亚家族1成员1BTN1A1M35551F:AGGACGGACTGGGCAATTGR:GAACCCATTCTCGGGAGTCAT81Bionaz等[11]黄嘌呤脱氢酶XDHBC102076F:GATCATCCACTTTTCTGCCAATGR:CCTCGTCTTGGTGCTTCCAA100自行设计
F:上游引物;R:下游引物。
F: forward primer; R: reverse primer.
1.4.3 BMECs内PPARγ和SREBP1的蛋白表达量
采用蛋白免疫印迹的方法测定,以GAPDH作为内参蛋白。将细胞以1×106个/mL的密度接种于25 cm2细胞培养瓶中,按试验设计培养结束后,弃掉上清,用PBS清洗细胞2遍,每瓶加入250 μL含0.1%苯甲基磺酰氟(PMSF)的RIPA细胞裂解液,4 ℃裂解5 min后刮下细胞并收集细胞悬液,4 ℃ 15 455×g离心10 min,取上清用于检测蛋白的表达量。取适量样品用BCA法测定样品的蛋白质浓度,随后分别向60 μg的每种样品中添加5×蛋白质上样缓冲液,按照4∶1的比例混合,100 ℃加热5 min使蛋白质热变性,然后进行电泳,浓缩胶80 V电泳30 min,分离胶120 V电泳120 min。电泳结束后,将分离的蛋白质转移到聚偏二氟乙烯(PVDF)膜上(100 V,4 ℃,50 min)。转膜后,吐温-TBS(TBST)漂洗3次,每次5 min,室温摇床上封闭1 h,TBST漂洗3次,每次5 min。然后将其分别与250倍稀释的兔抗PPARγ一抗稀释液和50倍的鼠抗SREBP1一抗稀释液结合,于4 ℃孵育过夜,取出PVDF膜后TBST漂洗3次,每次5 min。随后分别与稀释1 000倍的山羊抗兔二抗稀释液和稀释500倍的山羊抗鼠二抗稀释液结合,室温摇床孵育1 h,TBST漂洗3次,每次10 min。用ECL显色法试剂盒进行显色,在蛋白质成像仪上照相分析。图片用Quantity one软件进行灰度值分析,PPARγ和SREBP1蛋白表达量采用各组灰度值与0.5 mmol/L组灰度值的比值表示。
1.5数据处理
所有数据通过Excel进行整理,采用SAS 9.0软件的方差分析程序(ANOVA)进行显著性检验,同时用回归统计程序进行一次线性与二次曲线回归分析,P<0.05表示组间的差异或回归关系显著,0.05≤P<0.10表示组间的差异或回归关系趋于显著,P>0.10表示组间的差异或回归关系不显著。
2 结 果
2.1Lys对BMECs细胞活力、TAG含量及乳脂肪合成相关基因表达的影响
由表2可知,随着Lys浓度的增加,BMECs RGR呈显著的一次线性下降(P<0.001),TAG含量呈显著的二次曲线升高(P=0.013),FABP3和LPL基因表达量分别呈显著和趋于显著的一次线性增加(P=0.001和P=0.096),其中,4.0、8.0、16.0 mmol/L组的RGR显著低于0.5、1.0、2.0 mmol/L组(P<0.05),FABP3基因表达量以2.0、4.0、8.0、16.0 mmol/L组显著高于0.5、1.0 mmol/L组(P<0.05),LPL基因表达量以1.0、2.0、4.0、8.0、16.0 mmol/L组显著高于0.5 mmol/L组(P<0.05)。随着Lys浓度的增加,FASN基因表达量呈显著的二次曲线上升(P=0.003),最高值出现在2.0 mmol/L组,显著高于16.0 mmol/L组(P<0.05)。2.0、4.0 mmol/L组的SCD1基因表达量显著高于其他组(P<0.05)。2.0、4.0 mmol/L组PPARγ基因表达量显著高于0.5和8.0、16.0 mmol/L组(P<0.05)。1.0、2.0、4.0 mmol/L组的SREBP1基因表达量显著高于其他各组(P<0.05)。AGPAT6和GPAM基因表达量随Lys的增加均呈显著的一次线性降低(P=0.038和P=0.022),AGPAT6基因表达量以2.0、4.0、8.0、16.0 mmol/L组显著低于0.5、1.0 mmol/L组(P<0.05),GPAM基因表达量以16.0 mmol/L组显著低于0.5、1.0、2.0、4.0 mmol/L组(P<0.05)。LPIN1基因表达量以1.0、2.0、4.0、8.0 mmol/L组较高,显著高于0.5 mmol/L组(P<0.05)。BTN1A1和XDH基因表达量也以1.0、2.0、4.0、8.0 mmol/L组较高,显著高于其他各组(P<0.05)。
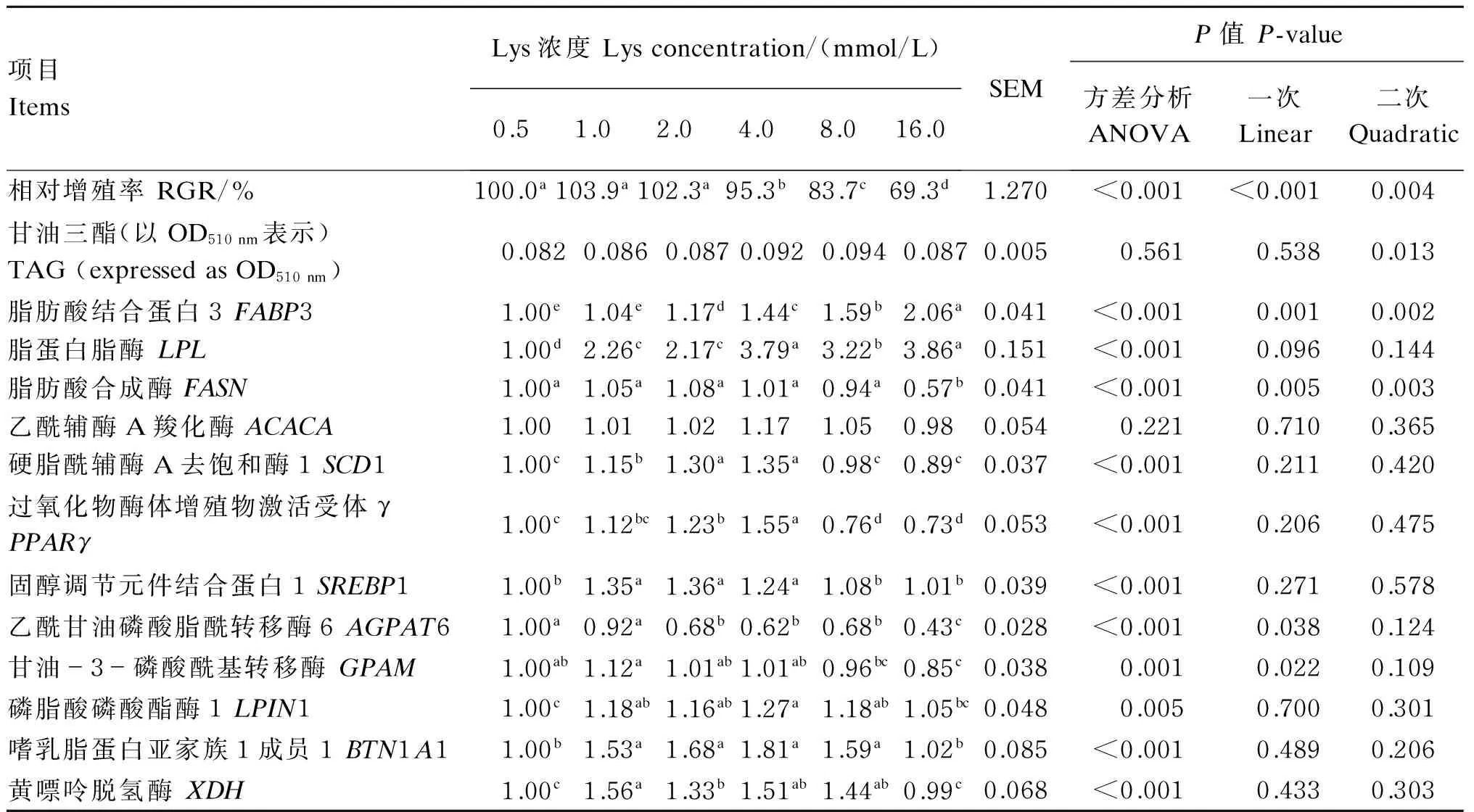
表2 Lys对BMECs细胞活力、TAG含量和乳脂肪合成相关基因表达量的影响
同行数据肩标相同或无字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。P<0.05表示回归关系显著;0.05≤P<0.10表示回归关系趋于显著;P>0.10表示回归关系不显著。下表同。
Values in the same row with the same or no letter superscripts mean no significant difference (P>0.05), while with different letter superscripts mean significant difference (P<0.05).P<0.05 means significant regression, 0.05≤P<0.10 means regression tend to be significant, andP>0.10 means no significant regression. The same as below.
2.2Lys对BMECsPPARγ和SREBP1蛋白表达的影响
由表3和图1可知,PPARγ蛋白表达量以1.0、2.0、4.0 mmol/L组显著高于其他组(P<0.05),SREBP1蛋白表达量以1.0 mmol/L组显著高于其他各组(P<0.05)。

表3 Lys对BMECs PPARγ和SREBP1蛋白表达的影响
3 讨 论
乳脂肪是衡量乳品质的重要指标之一[13],大约98%为TAG,其中,中短链脂肪酸(SMCFA)的含量占乳脂肪的2/3以上[14],奶牛乳汁中有50%的C16∶0脂肪酸以及几乎全部的C4∶0~C14∶0是由乳脂肪合成前体物(乙酸、β-羟丁酸)在乳腺从头合成的[15]。TAG在粗面内质网膜的表面合成,合成后以脂滴的形式积累,积累的脂滴通过质膜的包裹再由细胞分泌出去[16]。因此,BMECs内TAG含量和脂滴的累积能直接反映BMECs内乳脂肪的合成。研究表明,AA不仅能影响乳蛋白的合成,也能影响乳脂肪的合成[1]。李珊珊[17]研究表明,在BMECs中添加必需氨基酸显著促进乳蛋白的合成的同时,可能通过SREBP1和PPARγ调节乳脂肪的合成。然而,Lys对BMECs内的TAG合成的影响及其机理研究尚未见报道。本试验结果得出,随着Lys浓度增加,BMECs内TAG含量呈显著的二次曲线升高,说明Lys对TAG含量的影响呈浓度依赖关系,关于其影响机理尚不清楚,需要进一步研究。
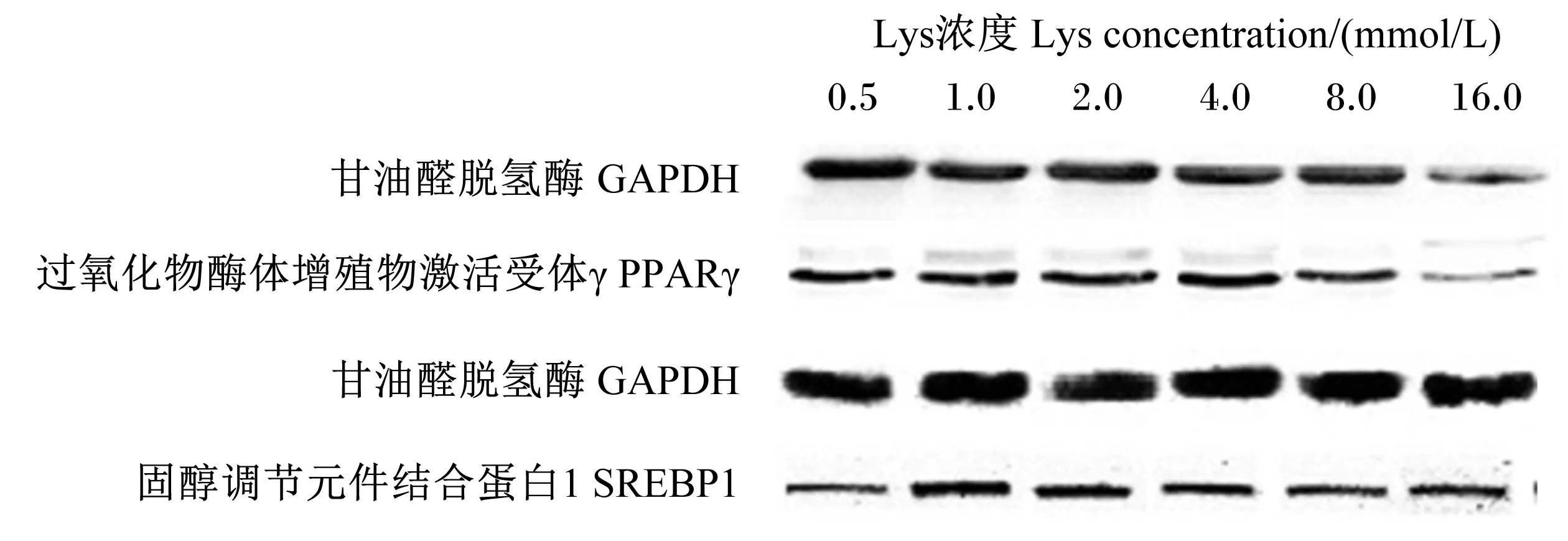
图1 Lys对BMECs PPARγ和SREBP1蛋白表达的影响
与细胞内的从头合成、长链脂肪酸(LCFA)的摄取与转运、脂滴的形成等过程有关的基因均会影响TAG的合成。哺乳动物组织中摄取和LCFA的主要基因是LPL和FABP3[18]。LPL能催化TAG分解为脂肪酸和甘油,为组织供能及贮存能量[19]。FABP3与细胞内LCFA的转运有关,可将LCFA从细胞膜上运送到TAG和磷脂的合成位点[20]。赵艳丽等[21]研究表明,添加适宜浓度的蛋氨酸对BMECs内FABP3和LPL基因表达具有显著的促进作用,说明蛋氨酸可以促进BMECs内LCFA的摄取与转运。本研究结果得出,2.0、4.0、8.0、16.0 mmol/L组FABP3基因表达量、1.0、2.0、4.0、8.0、16.0 mmol/L组LPL基因表达均显著高于0.5 mmol/L组,且二者均随着Lys浓度的增加呈显著和趋于显著的一次线性增加,说明Lys对BMECs内LCFA的摄取与转运可能具有一定的促进效果,提示Lys对乳脂肪合成的促进效果呈浓度依赖关系。
FASN是一种多功能的酶系统,参与脂肪酸合成的整个过程,FASN是奶牛乳脂肪酸从头合成过程中的关键基因[11],泌乳期奶牛乳腺内SMCFA(C4~C16)的合成受FASN编码的蛋白调控[22]。并且,FASN催化乙酰辅酶A和丙二酰辅酶A而生成LCFA。硬脂酰辅酶A去饱和酶(SCD)是Δ9去饱和酶中一种主要的酶,是细胞中单不饱和脂肪酸合成的限速酶[20]。Li等[23]研究发现,添加不同比例的含Lys的必需氨基酸可上调BMECs内LPIN1、FASN和SCD的基因表达量。本试验结果表明,Lys对FASN基因表达的促进效果呈显著的浓度依赖效应,尤以2.0 mmol/L组最高,显著高于16.0 mmol/L组;Lys也促进SCD1基因表达,以2.0、4.0 mmol/L组显著高于其他组。这些研究结果部分地解释了Lys对乳脂肪合成具有促进效果的原因。
此外,GPAM、AGPAT6和LPIN1参与催化TAG的合成[24],是参与乳脂肪合成的关键酶。GPAM催化脂酰辅酶A与甘油-3-磷酸的sn-1位点结合形成溶血磷脂酸;AGPAT6催化第2个脂酰辅酶A与甘油-3-磷酸的sn-2位点结合形成磷脂酸(PA);LPIN能转移磷酸基团,将PA转变成二酰甘油。研究发现敲除了泌乳小鼠的AGPAT6基因后不能合成乳脂肪[25]。本研究结果发现,Lys对GPAM和AGPAT6基因表达量的作用效果呈显著的浓度依赖效应,随着Lys浓度的增加抑制了它们的表达,AGPAT6以2.0、4.0、8.0、16.0 mmol/L组显著低于0.5、1.0 mmol/L组,GPAM以16.0 mmol/L组显著低于0.5、1.0、2.0、4.0 mmol/L组;同时,Lys显著地促进了LPIN1基因表达,以1.0、2.0、4.0、8.0 mmol/L组较高。这些研究结果说明,高浓度的Lys抑制了乳脂肪合成相关基因的表达,进一步解释了Lys对乳脂肪合成的促进效果呈浓度依赖效应的原因。
BTN1A1和XDH是调控脂滴形成的主要蛋白[26]。固醇调节元件结合蛋白(SREBP)是核转录因子家族成员之一,过氧化物酶体增殖物激活受体(PPAR)属于核激素受体家族中的配体激活受体,PPARγ可调控SREBP1基因的表达,同时LPL和ACACA等是PPARγ的标靶基因[27]。Kadegowda等[28]用PPARγ激活剂处理BMECs后发现,上调了ACACA、FASN、SREBF1、SCD和LPIN1基因的表达。本研究结果显示,BTN1A1和XDH基因表达量均以1.0、2.0、4.0、8.0 mmol/L组较高;PPARγ基因及蛋白表达量均以2.0、4.0 mmol/L组显著高于0.5和8.0、16.0 mmol/L组;SREBP1基因表达量以1.0、2.0、4.0 mmol/L组较高,蛋白表达量以1.0、2.0 mmol/L组较高,进一步从转录因子和脂滴形成的角度解释了Lys对乳脂肪合成具有促进效果的原因。FABP3、LPL、FASN、GPAM、AGPAT6等是PPARγ和SREBP1的靶基因,在本研究只对PPARγ和SREBP1的蛋白表达量进行了探讨研究,而对FABP3、LPL、FASN、GPAM、AGPAT6等蛋白表达量尚未开展研究,有必要今后继续研究探讨,以更好地阐明Lys对乳脂肪合成的影响机理。
生冉[29]研究指出,雷帕霉素抑制雷帕霉素靶蛋白(mTOR)信号通路后,SREBP1、PPARγ、ACACA和SCD1的基因表达量显著下降;Soliman等[30]的试验研究也得出了相似的结果,雷帕霉素抑制了人乳腺外植体中SREBP1及其靶基因ACACA、FASN和SCD1的基因表达,说明mTOR信号通路通过转录因子SREBP1和PPARγ调控乳脂肪的合成。Lys是否通过mTOR信号通路间接地调控乳脂肪的合成尚未见资料报道,本试验并未对mTOR信号通路相关基因进行检测,需要进一步深入研究。
综合脂肪酸摄取与转运、从头合成、TAG的合成及脂滴形成的相关基因表达结果可以看出,Lys对BMECs内的乳脂肪合成具有一定的促进效果,呈浓度依赖效应,以2.0~4.0 mmol/L的Lys添加浓度为宜,且Lys对BMECs细胞活力的影响也呈显著浓度依赖效应。然而目前关于添加Lys对奶牛乳脂肪合成及其调控机理的研究甚少,并且本试验得出的结果还没有在体内进一步验证,因此,需要进一步深入研究。
4 结 论
① Lys对BMECs的乳脂肪合成具有显著的促进效果,但高浓度的Lys抑制了乳脂肪合成相关基因的表达。
② 本试验条件下,培养基中Lys适宜浓度为2.0~4.0 mmol/L。
[1] MAXIN G,RULQUIN H,GLASSER F.Response of milk fat concentration and yield to nutrient supply in dairy cows[J].Animal,2011,5(8):1299-1310.
[2] GIALLONGO F,HARPER M T,OH J,et al.Effects of rumen-protected methionine,lysine,and histidine on lactation performance of dairy cows[J].Journal of Dairy Science,2016,99(6):4437-4452.
[3] XU Y T,ZENG Z K,XU X,et al.Effects of the standardized ileal digestible valine:lysine ratio on performance,milk composition and plasma indices of lactating sows[J].Animal Science Journal,2016,doi:10.1111/asj.12753.
[4] 韩慧娜.秸杆日粮条件下阴外动脉灌注乳脂和乳蛋白前体物对奶牛乳腺内短链脂肪酸摄取规律的影响[D].硕士学位论文.呼和浩特:内蒙古农业大学,2015.
[5] SHENG R,YAN S M,QI L Z,et al.Effect of the ratios of acetate and β-hydroxybutyrate on the expression of milk fat- and protein-related genes in bovine mammary epithelial cells[J].Czech Journal of Animal Science,2015,60(12):531-541.
[6] 李喜艳.奶牛乳腺上皮细胞中赖氨酸蛋氨酸配比模式对酪蛋白合成的影响及机理研究[D].硕士学位论文.北京:中国农业科学院,2011.
[7] 高海娜.亮氨酸、组氨酸、赖氨酸和蛋氨酸对奶牛乳腺上皮细胞中酪蛋白合成的影响及调控机理研究[D].硕士学位论文.兰州:甘肃农业大学,2016.
[9] ZHOU Y,AKER R M, JIANG H.Growth hormone can induce expression of four major milk protein genes in transfected MAC-T cells[J].Journal of Dairy Science,2008,91(1):100-108.
[10] QI L Z,YAN S M,SHENG R,et al.Effects of saturated long-chain fatty acid on mRNA expression of genes associated with milk fat and protein biosynthesis in bovine mammary epithelial cells[J].Asian-Australasian Journal of Animal Sciences,2014,27(3):414-421.
[11] BIONAZ M,LOOR J J.Gene networks driving bovine milk fat synthesis during the lactation cycle[J].BMC Genomics,2008,9:366.
[12] 张养东.脂多糖对泌乳奶牛乳脂肪和乳蛋白影响及其机理研究[D].博士学位论文.哈尔滨:东北农业大学,2011.
[13] HARVATINE K J,BOISCLAIR Y R,BAUMAN D E.Recent advances in the regulation of milk fat synthesis[J].Animal,2009,3(1):40-54.
[14] JENSEN R G.The composition of bovine milk lipids:January 1995 to December 2000[J].Journal of Dairy Science,2002,85(2):295-350.
[15] HARFOOT C G,HAZLEWOOD G P.Lipid metabolism in the rumen[M]//HOBSON P N,STEWART C S.The rumen microbial ecosystem.Netherlands:Springer,1997.
[16] SOLIMAN G A.The integral role of mTOR in lipid metabolism[J].Cell Cycle,2011,10(6):861-862.
[17] 李珊珊.必需氨基酸调节奶牛乳腺合成乳蛋白和乳脂肪的作用机制[D].博士学位论文.杭州:浙江大学,2015.
[18] LRHNER R,KUKSIS A.Biosynthesis of triacylglycerols[J].Progress in Lipid Research,1996,35(2):169-201.
[19] BONNET M,LEROUX C,CHILLIARD Y,et al.Rapid communication:nucleotide sequence of the ovine lipoprotein lipase cDNA[J].Journal of Animal Science,2000,78(11):2994-2995.
[20] 胡菡,王加启,李发弟,等.奶牛乳腺脂肪酸合成相关基因研究进展[J].生物技术通报2009(10):34-39.
[21] 赵艳丽,陈璐,史彬林,等.亮氨酸对奶牛乳腺上皮细胞内乳脂合成相关基因和蛋白表达的影响[J].动物营养学报,2017,29(4):1319-1326.
[22] WAKIL S J.Fatty acid synthase,a proficient multifunctional enzyme[J].Biochemistry,1989,28(11):4523-4530.
[23] LI S S,HOSSEINI A,et al.Essential amino acid ratios and mTOR affect lipogenic gene networks and miRNA expression in bovine mammary epithelial cells[J].Journal of Animal Science and Biotechnology,2016,7:44.
[24] COLEMAN R A,LEE D P.Enzymes of triacylglycerol synthesis and their regulation[J].Progress in Lipid Research,2004,43(2):134-176.
[25] BEIGNEUX A P,VERGNES L,QIAO X,et al.AGPAT6-a novel lipid biosynthetic gene required for triacylglycerol production in mammary epithelium[J].Journal of Lipid Research,2006,47(4):734-744.
[26] MCMANAMAN J L,RUSSEL T D,SCHAACK J,et al.Molecular determinants of milk lipid secretion[J].Journal of Mammary Gland Biology and Neoplasia,2007,12(4):259-268.
[27] DESVERGNE B,MICHALIK L,WAHLI W.Transcriptional regulation of metabolism[J].Physiological Reviews,2006,86(2):465-514.
[28] KADEGOWDA A,BIONAZ M,PIPEROVA L,et al.Lipogenic gene expression in MAC-T cells is affected differently by fatty acids and enhanced by PPAR-gamma activation[J].Journal of Dairy Science,2008,91:678.
[29] 生冉.乙酸参与奶牛乳腺上皮细胞内乳脂肪和乳蛋白合成的调控机理研究[D].博士学位论文.呼和浩特:内蒙古农业大学,2015.
[30] SOLIMAN M,KIMURA K,AHMED M,et al.Inverse regulation of leptin mRNA expression by short- and long-chain fatty acids in cultured bovine adipocytes[J].Domestic Animal Endocrinology,2007,33(4):400-409.
*Corresponding author, professor, E-mail: yansmimau@163.com
(责任编辑 王智航)
Effects of Lysine on Expressions of Genes and Proteins Involved in Milk Fat Synthesis in Bovine Mammary Epithelial Cells
CHEN Lu ZHAO Yanli GUO Xiaoyu SHI Binlin YAN Sumei*
(Collage of Animal Science, Inner Mongolia Agriculture University, Hohhot 010018, China)
This study was to detect the effects of lysine (Lys) on expressions of genes and proteins involved in milk fat synthesis in bovine mammary epithelial cells (BMECs) and discuss the mechanism of Lys regulating milk fat synthesis. The 3rd passage BMECs were divided into six groups with six replicates per group and one pore per replicate. Cells were cultured in medium containing 0.5 (basal medium, control), 1.0, 2.0, 4.0, 8.0 and 16.0 mmol/L Lys, respectively. The triglyceride (TAG) content, expressions of genes and proteins involved in milk fat synthesis were detected after 48 h cultivation at 37 ℃ and 5% CO2. The results showed as follows: TAG content (P=0.013) and gene expressions of fatty acid-binding protein 3 (FABP3,P=0.001), lipoprotein lipase (LPL,P=0.096), fatty acid synthase (FASN,P=0.003), 1-acylglycerol-3-phosphate O-acyltransferase 6 (AGPAT6,P=0.038) and glycerol-3-phosphate acyltrandferase (GPAM,P=0.022) acted dose-dependent on Lys at significant level or significant tendency. Compared with 0.5 mmol/L group, 2.0, 4.0, 8.0 and 16.0 mmol/L groups significantly increased gene expression ofFABP3 (P<0.05), and 1.0, 2.0, 4.0, 8.0 and 16.0 mmol/L groups significantly increased gene expression ofLPL(P<0.05). Gene expression ofFASNin 2.0 mmol/L group was the highest, which was significantly higher than that in 16.0 mmol/L group (P<0.05). Gene expression of stearoyl-CoA desaturase 1 (SCD1) in 2.0 to 4.0 mmol/L groups was significantly higher than that in other groups (P<0.05). Gene expressions of phosphatidic acid phosphatase 1 (LPIN1), butyrophilin subfamily 1 member A1 (BTN1A1) and xanthine dehydrogenase (XDH) in 1.0, 2.0, 4.0 and 8.0 mmol/L groups were significantly higher than those in 0.5 mmol/L group. Compared with the 0.5, 8.0 and 16.0 mmol/L groups, 2.0 and 4.0 mmol/L groups significantly increased gene and protein expression of peroxisome proliferator-activated receptor-γ (PPARγ) (P<0.05). Gene expression of sterol regulatory element binding protein 1 (SREBP1) in 1.0, 2.0 and 4.0 mmol/L groups was significantly higher than that in the other groups (P<0.05), and protein expression ofSREBP1 in 1.0 mmol/L group was significantly higher than that in the other groups (P<0.05). High dose of Lys decreased gene expressions ofAGPAT6 andGPAM. Compared with 0.5 and 1.0 mmol/L groups, 2.0, 4.0, 8.0 and 16.0 mmol/L groups significantly decreased gene expression ofAGPAT6 (P<0.05). Compared with 0.5, 1.0, 2.0 and 4.0 mmol/L groups, 16.0 mmol/L groups significantly decreased gene expression ofGPAM(P<0.05). In conclusion, Lys significantly promote milk fat synthesis in BMECs, but high dose of Lys inhibits gene expressions related in milk fat synthesis. Under the conditions in the present study, 2.0 to 4.0 mmol/L of Lys is an optimal level in culture medium.[ChineseJournalofAnimalNutrition,2017,29(9):3366-3374]
dairy cow; bovine mammary epithelial cells; lysine; milk fat
10.3969/j.issn.1006-267x.2017.09.042
2017-02-24
国家奶业“973计划”项目(2011CB1008003)
陈 璐(1990—),女,山西襄汾人,硕士研究生,从事奶牛营养研究。E-mail:1510560671@qq.com
*通信作者:闫素梅,教授,博士生导师,E-mail: yansmimau@163.com
S823
:A
:1006-267X(2017)09-3366-09
猜你喜欢
今日农业(2022年3期)2022-11-16
今日农业(2021年10期)2021-07-28
中国生殖健康(2020年5期)2021-01-18
小天使·一年级语数英综合(2019年12期)2019-01-13
中国生殖健康(2019年5期)2019-01-06
中国生殖健康(2018年5期)2018-11-06
妈妈宝宝(2017年2期)2017-02-21
中国医学科学院学报(2015年5期)2015-03-01
中国洗涤用品工业(2015年8期)2015-02-28
小天使·三年级语数英综合(2014年9期)2014-09-12
