
番茄环纹斑点病毒Gn蛋白的原核表达及多抗血清制备
2017-09-16 02:35郑宽瑜尚卫娜张洁郑雪董家红
生物技术通报 2017年9期
郑宽瑜尚卫娜张洁郑雪董家红
(1. 云南省农业科学院生物技术与种质资源研究所 云南省农业生物技术重点实验室 农业部西南作物基因资源与种质创新重点实验室,昆明650223;2. 浙江大学生命科学研究院,杭州 310058)
番茄环纹斑点病毒Gn蛋白的原核表达及多抗血清制备
郑宽瑜1尚卫娜2张洁1郑雪1董家红1
(1. 云南省农业科学院生物技术与种质资源研究所 云南省农业生物技术重点实验室 农业部西南作物基因资源与种质创新重点实验室,昆明650223;2. 浙江大学生命科学研究院,杭州 310058)
通过生物信息学预测,番茄环纹斑点病毒(Tomato zonate spot virus,TZSV)Gn蛋白为跨膜蛋白,有3个跨膜结构域,抗原表位预测发现非跨膜区包含有较高的抗原指数区域,因此选择Gn蛋白的非跨膜区进行原核表达及多抗血清制备。用特异性引物,通过RT-PCR从感染TZSV的番茄中扩增出部分Gn基因片段,将获得的片段构建到原核表达载体pET-30a中,获得原核表达载体pET-30a-Gn,转化大肠杆菌BL21(DE3),用IPTG诱导表达。SDS-PAGE电泳分析显示,高效诱导表达了40 kD融合蛋白。用回收纯化的融合蛋白免疫家兔,获得多抗血清。间接ELISA测定多抗血清的效价为1/16 384。利用制备好的抗血清对感染TZSV的病样进行Western blotting 检测,能检测到特异性条带。结果表明该抗血清特异性良好,可用于TZSV Gn相关的免疫反应实验。
番茄环纹斑点病毒;Gn蛋白;原核表达;抗血清
番茄环纹斑点病毒(Tomato zonate spot virus,TZSV)属于布尼亚病毒科(Bunyaviridae),番茄斑萎病毒属(Tospovirus),自2005年发现以来,一直是云南番茄、辣椒、烟草等经济作物的重要病原[1],由蓟马传毒为害[2]。TZSV与其它番茄斑萎病毒属病毒类似,基因组为单链线状三分体RNA:S RNA、M RNA、L RNA。S RNA正链编码非结构蛋白NSs,负链编码核壳体蛋白N;M RNA正链编码移动蛋白NSm,负链编码糖蛋白Gn/Gc;L RNA负链编码病毒的复制酶RdRp。
TZSV为球形病毒,直径约80-120 nm,具有双层脂膜组成的包膜,包膜表面有 Gn/Gc 糖蛋白构成的突起。相关证据表明Gn/Gc糖蛋白是Tospovirus病毒与蓟马识别的关键蛋白[3]。Gn/Gc 蛋白是由前体蛋白G经剪切而形成的,TZSV的前体蛋白G大小为3 369 bp,编码1 122 aa,其中,氨基酸1-25位为信号肽,经剪切后,进一步在高尔基内进行N-糖基化修饰形成成熟的Gn蛋白和Gc蛋白。对同为布尼亚病毒科的汉坦病毒(Hantavirus)研究显示:Gn和 Gc 结构相似,均由胞外区、跨膜区及C末端胞质尾区(GP-CT)3部分组成[4]。由于Gn是膜蛋白,具有跨膜结构域,用全长Gn不能顺利表达出目的蛋白。本研究通过生物信息学的方法,预测Gn蛋白的抗原表位和跨膜结构域,通过选择Gn蛋白的非跨膜区进行Gn的原核表达,并通过免疫家兔得到了专一性的Gn蛋白多抗血清,旨为研究TZSV与蓟马互作奠定基础。
1 材料与方法
1.1 材料
TZSV感染的番茄由本实验室保存。Marker DL2000、pMD Simple18-T 载体、rTaq酶、限制性内切酶Nco I及Xho I为TaKaRa公司产品。M-MLV为Promega公司产品。DNA回收试剂盒购自Sangon公司。IPTG、NBT及BCIP购自Sigma公司。硝酸纤维素酶购自BioRad公司。Ni+亲和纯化柱购自GE公司。1.2 方法
1.2.1 Gn蛋白结构及Protean预测 从NCBI下载TZSV Gn/Gc蛋白的基因序列(序列号:NC_010490),用SignalP-4.0预测Gn/Gc蛋白的剪切位点;TMHMM预测Gn跨膜结构。利用DNAstar软件提供的Protean模块,用Kyte-Doolittle方法预测Gn蛋白亲水性;Eisenberg方法预测Gn蛋白二级结构;Karplas-Schulz方法预测蛋白质骨架区的柔韧性;Jameson-wolf方法预测蛋白潜在的抗原表位;用Emini方法预测蛋白的表面可及性[5]。
1.2.2 Gn基因的RT-PCR扩增及原核表达载体构建 采用LiCI沉淀法提取RNA[1],两步法RT-PCR扩增Gn序列。上游引物Gn-F:CATGCCATGGCATCGAAGACGGAGCATGAA(下划线为限制性内切酶Nco I);下游引物Gn-R:CCGCTCGAGTCATGAGAAGTTCGTGCAGA(下划线为限制性内切酶Xho I)。cDNA合成体系为20 μL,用12 μL总RNA为模板,1 μL 100 nmol/L引物混合,70℃ 预变性10 min后,迅速插入冰上5 min,加入5×RT Buffer 4 μL,10 mmol/L dNTP 2 μL,5 U/μL M-MLV 1 μL,40 U/μL RNase Inhibitor 1 μL,42℃延伸60 min。以cDNA为模板进行PCR扩增,反应体系为:94℃下预变性4 min;然后94℃下变性1 min,55℃退火1 min,72℃下延伸1 min 30 s,循环扩增30次;最后72℃下延伸10 min。PCR产物经切胶纯化后,克隆到pMD simple 18-T载体上。菌落PCR鉴定阳性克隆并送华大基因测序。将Gn基因用Nco I及Xho I双酶切后,与表达载体pET-30a相连接,转化大肠杆菌BL21(DE3),菌落PCR鉴定阳性克隆并进一步测序验证编码框是否正确。
1.2.3 Gn融合蛋白的诱导表达、SDS-PAGE分析和纯化 阳性克隆接菌于含100 mg/L卡那霉素(Kanamycin)的LB培养基中,培养至对数生长期,OD600=0.6时加入终浓度为1 mmol/L的IPTG 37℃诱导表达6 h,以不加IPTG诱导的培养物为阴性对照。离心收集菌体,在8 mol/L尿素变性条件下破碎细胞,用Ni+亲和纯化柱回收带有His标签的融合蛋白。纯化后的蛋白在复性缓冲液(50 mmol/L Tris,0.5 mM EDTA,50 mmol/L NaCl,1%甘氨酸)中透析48 h,作为抗原免疫家兔。同时取样加2×SDS-PAGE上样缓冲液(0.1 mol/L Tris-HCl,1% β-巯基乙醇,4% SDS,0.2%溴酚蓝,20%甘油,pH 6.8)充分悬浮。沸水中处理10 min,12 000 r/min离心2 min后取上清进行SDS-PAGE电泳分析。
1.2.4 多抗血清的制备及效价测定 用紫外分光光度计测定Gn蛋白浓度,稀释至1 mg/mL,参照欧阳元龙等[6]的方法免疫家兔。抗血清按1/512、1/1 024、1/2 048、1/4 096、1/8 192、1/16 384倍比进行梯度稀释,间接ELISA进行抗血清效价测定[7]。
1.2.5 Gn蛋白的Western blotting检测 取0.1 g感病叶片液氮研磨后,加入200 μL 2×SDS蛋白上样缓冲液,进行SDS-PAGE电泳,电泳结束后电转移至硝酸纤维素膜上,TZSV Gn多抗血清孵育1 h(1∶4 000倍比稀释),TBST洗膜后,加入碱性磷酸酯酶标记羊抗兔二抗(1∶8 000倍比稀释),加入NBT/BCIP显色至条带清晰。
2 结果
2.1 TZSV Gn蛋白结构及Protean预测
SignalP-4.0 预测前体蛋白G氨基酸序列第25位为信号肽剪接位点,Gn和Gc的剪切位点位于前体蛋白G氨基酸序列第436位,由此推算Gn蛋白核苷酸序列长1 233 bp,编码411 aa,分子量为47.5 kD。TMHMM预测前体蛋白G氨基酸第295位后有连续3个跨膜结构(图1)。在原核表达中,信号肽及跨膜结构会影响到目的蛋白的定位及表达量;此外综合Kyte-Doolittle、Eisenberg、Karplas-Schulz、Jameson-wolf方法、Emini方法,Gn蛋白氨基酸残基1-260区段内有较好的亲水性、可及性和较高的抗原性指数(图2),因此在本研究中选择前体蛋白G氨基酸序列26-295之间的区域,基因片段长度810 bp用于Gn蛋白的原核表达及多抗血清制备。
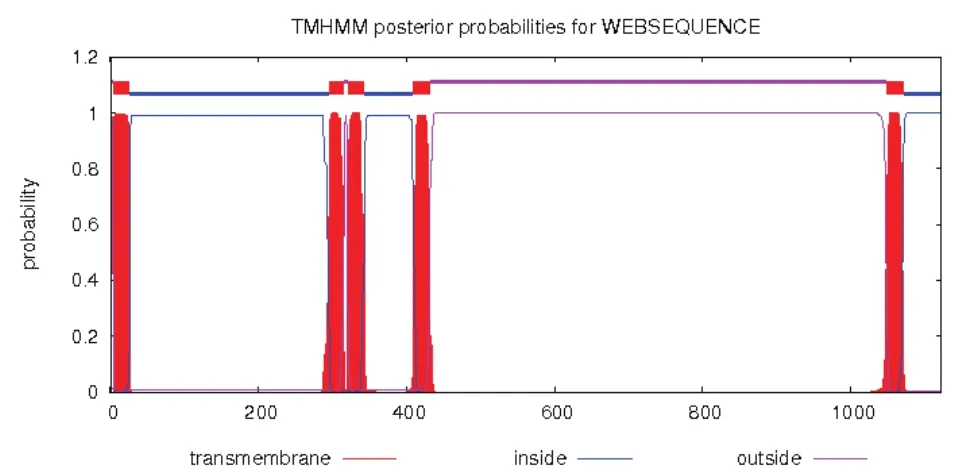
图1 TMHMM预测前体G蛋白跨膜结构域
2.2 TZSV Gn原核表达载体构建、诱导表达及纯化
用RT-PCR从感病番茄中扩增得到长度为810 bp的Gn基因,经1%琼脂糖电泳分析与目的片段大小相吻合,PCR产物纯化后与pMD simple 18-T连接,并测序。证实扩增片段即为Gn基因片段,且ORF正确。用Nco I及Xho I双酶切原核表达载体pET-30a-Gn,结果显示从载体上能特异性切下一段约810 bp左右的片段。同时测序结果也说明该片段即为TZSV的Gn基因,证明所构建的原核表达载体是正确的。转化有原核表达载体pET-30a-Gn阳性克隆的大肠杆菌BL21菌株在LB液体培养基中培养,然后加入1 mmol/L IPTG诱导表达6 h,而不加IPTG诱导的阳性克隆菌株则作为对照。经12.5%的SDSPAGE分析显示,IPTG诱导的菌株特异性表达约40 kD融合蛋白(图3)。该融合蛋白在N端和C端都表达有His-tag标签。用GE公司的Ni+亲和纯化柱纯化回收带有His标签的Gn蛋白。
2.3 TZSV Gn蛋白的抗血清制备及效价测定
以纯化后的Gn蛋白作为抗原免疫家兔,每周注射一次,注射4次后取血收集血清,用间接ELISA测定抗血清的效价。结果显示1 mg/mL的抗原能和多抗血清反应的最高稀释倍数为1/16 384倍。2.4 TZSV Gn蛋白的Western blot检测

图2 Protean预测Gn蛋白抗原表位
用制备好的抗血清对TZSV感病病样及原核表达的Gn蛋白进行Western blot检测(图4),在约40 kD处检测到原核表达的Gn蛋白,在约47 kD附近检测到TZSV感病病样的Gn蛋白条带,与预期相符合。

图3 Gn蛋白的诱导表达及纯化

图4 Gn蛋白的Western blot检测
3 讨论
Gn/Gc糖蛋白为布尼亚病毒科病毒的重要结构蛋白,它镶嵌在成熟病毒颗粒的包膜上,在感染人或动物的布尼亚病毒中,因其具有中和抗原决定簇和各型的特异性抗原位点,能够刺激机体产生特异性抗体,而成为基因工程疫苗研究的靶点[8,9]。本研究在对感染植物Tospovirus病毒TZSV Gn蛋白结构及Protean预测发现,TZSV Gn蛋白氨基酸残基1-260区段内有较好的亲水性、可及性和较高的抗原性指数,制备的抗血清具有良好的特异性。用本实验室制备的Gn抗血清做TZSV Gn蛋白免疫胶体金标记,Gn蛋白抗体能很好的定位到病毒粒子表面,具有非常好的电镜形态学诊断特征[10]。
Gn蛋白一个重要的功能是介导病毒与细胞受体的识别。对感染人的汉坦病毒(Hantavirus)研究显示Gn蛋白可以介导病毒与细胞蛋白β3整合素(β3 integrins)、衰变加速因子(DAF/CD55)、补体分子球形结构域受体(gC1qR/p32)识别而使病毒进入细胞[11-12]。对感染植物的Tospovirus病毒代表种番茄斑萎病毒(Tomato spotted wilt virus,TSWV)的Gn定位研究表明,Gn能特异性定位到西花蓟马(Frankliniella occidentalis)中肠部位,表明与TSWV Gn识别的受体蛋白可能为蓟马中肠受体蛋白[13],也发现了可能为Gn受体的蓟马蛋白[14-15]。在所有已知的5 500多种蓟马中,仅发现蓟马科(Thripidae)的花蓟马属(Frankliniella)、蓟马属(Thrips)、硬蓟马属(Scirtothrips)及缘蓟马属(Ceratothripoides)4个属的12种蓟马能够传播Tospoviruses[16]。推测Tospovirus病毒与蓟马的特异性很可能是Gn/Gc蛋白与蓟马中肠受体蛋白特异性密切相关。本研究所制备的TZSV Gn蛋白多抗血清不仅可以应用于病毒快速检测及病毒发生流行监测,还可进一步利用免疫标记、far-western blotting、免疫共沉淀及酵母双杂等技术应用于Gn蛋白与相关传毒介体蓟马的互作功能研究。
4 结论
本研究通过生物信息学方法对Gn蛋白结构及B细胞的潜在抗原表位预测,选择了Gn的非跨膜区进行原核表达及多抗血清制备,所制备的抗血清具有较高的效价,1 mg/mL的抗原能和多抗血清反应的最高稀释倍数为1/16 384倍。Western blot表明多抗血清特异性好,不仅能检测到原核表达的Gn蛋白(40 kD),也能检测到感染TZSV病样中的Gn蛋白(49 kD)。该多抗血清不仅能应用于TZSV检测,还能进一步应用于Gn的功能研究。
[1]Dong JH, Cheng XF, Yin YY, et al. Characterization of Tomato zonate spot virus, a new Tospovirus species in China[J]. Archives of Virology, 2008, 156(5):755-764.
[2]Whitfield AE, Ullman DE, German TL. Tospovirus-thrips interactions[J]. Annual Review of Phytopathology, 2005, 43(7):459-489.
[3]Medeiros RB, Ullman DE, Sherwood JL, et al. Immunoprecipitation of a 50-kDa protein:a candidate receptor component for Tomato spotted wilt tospovirus(Bunyaviridae)in its main vector, Frankliniella occidentalis[J]. Virus Research, 2000, 67(2):109-18.
[4]盛欣, 兰英华, 李用国. 汉坦病毒糖蛋白及其在细胞融合中作用的研究进展[J]. 医学研究杂志, 2013, 42(12):16-18.
[5]马凡舒, 张蕾, 王洋, 孙彦刚, 闫喜军. B细胞抗原表位预测方法的研究进展[J]. 中国畜牧兽医, 2016, 43(1):63-67.
[6]欧阳元龙, 吴建祥, 熊如意, 等. 水稻黑条矮缩病病毒外壳蛋白基因S10的原核表达、多克隆抗体制备及应用[J]. 中国水稻科学, 2010, 24(1):25~30.
[7]Wang M, Gonsalves D. ELISA detection of various Tomato spotted wilt virus isolates using specific antisera to structural proteins of the virus[J]. Plant Disease, 1990, 74(2):154-158.
[8]Faburay B, Lebedev M, McVey DS, et al. A glycoprotein subunit vaccine elicits a strong Rift Valley fever virus neutralizing antibody response in sheep[J]. Vector Borne Zoonotic Dis, 2014, 14(10):746-756.
[9]于蒙蒙, 于澜. 汉坦病毒糖蛋白的结构及功能研究进展[J].微生物学免疫学进展, 2012, 40(4):63-66.
[10]郑宽瑜, 董家红, 方琦, 等. 胶体金免疫电镜技术检测番茄环纹斑点病毒[J]. 电子显微学报, 2015, 34(1):67-70.
[11]Jin M, Park J, Lee S, et al. Hantaan virus enters cells by clathrindependent receptor-mediated endocytosis[J]. Virology, 2002, 294(1):60-69.
[12]Cifuentes-Muñoz N, Salazar-Quiroz N, Tischler ND. Hantavirus Gn and Gc envelope glycoproteins:key structural units for virus cell entry and virus assembly[J]. Viruses, 2014, 6(4):1801-1822.
[13]Whitfield AE, Ullman DE, German TL. Expression and characterization of a soluble form of Tomato spotted wilt virus glycoprotein Gn[J]. Journal of Virology, 2004, 78(23):13197-13206.
[14]Bandla MD, Campbell LR, Ullman DE, et al. Interaction of Tomato spotted wilt tospovirus(TSWV)glycoproteins with a thrips midgut protein, a potential cellular receptor for TSWV[J]. Virology, 1998, 88(2):98-104.
[15]Jones DR. Plant viruses transmitted by thrips[J]. European Journal of Plant Pathology, 2005, 113(2):119-157.
[16]Sin SH, McNulty BC, Kennedy GG, et al. Viral genetic determinants for thrips transmission of Tomato spotted wilt virus[J]. PNAS, 2005, 102(14):5168-5173.
(责任编辑 朱琳峰)
Prokaryotic Expression and Polyclonal Antiserum Preparation of Gn Protein in Tomato Zonate Spot Virus(TZSV)
ZHENG Kuan-yu1SHANG Wei-na2ZHANG Jie1ZHENG Xue1DONG Jia-hong1
(1. Institute of Biotechnology and Germplasm Resources,Yunnan Academy of Agricultural Sciences,Yunnan Province Key Laboratory of Agricultural Biotechnology,Key Lab of the Southwestern Crop Gene Resources and Germplasm Innovation,Ministry of Agriculture,Kunming 650223;2. Life Sciences Institute,Zhejiang University,Hangzhou 310058)
Bioinformatics prediction showed that Gn protein in tomato zonate spot virus(TZSV)is a transmembrane protein consisting of 3 transmembrane domains. Antigen epitope prediction showed that there was a high antigen index region,thus non-transmembrane region of Gn was selected to conduct the prokaryotic expression and to prepare polyclonal antiserum. The Gn gene fragment of TZSV was amplified from TZSV-infected tomato sample by RT-PCR,and then cloned into prokaryotic expression vector pET-30a. The recombinant plasmids pET-30a-Gn were transformed into Escherichia coli BL21(DE3),and then induced to express with IPTG. The result of SDS-PAGE analysis showed the specific fusion protein with molecular weight of 40 kD was highly expressed. Using the purified fusion protein,rabbit was immunized to obtain polyclonal antiserum against TZSV Gn protein. The titer of antiserum was 1/16384 determined by ID-ELISA. The specific band was detected by Western blot while applying the prepared antiserum to the TZSV-infected samples. These results reveal that the specificity of the antiserum is fine,and it is applicable for immune-reaction test related to TZSV Gn.
tomato zonate spot virus;Gn protein;prokaryotic expression;antiserum
10.13560/j.cnki.biotech.bull.1985.2017-0242
2017-03-28
国家自然科学基金项目(31660508,31371919,31060237)
郑宽瑜,女,副研究员,研究方向:植物病理学;E-mail:zhengkuanyu@126.com尚卫娜,女,实验师,研究方向:植物病理学;E-mail:wnshang@zju.edu.cn。尚卫娜与郑宽瑜为共同第一作者
董家红,男,研究员,研究方向:植物病理学;E-mail:dongjhn@126.com
猜你喜欢
今日农业(2021年11期)2021-11-27
现代园艺(2017年21期)2018-01-03
安徽医科大学学报(2015年9期)2015-12-16
动物医学进展(2015年10期)2015-12-07
动物医学进展(2015年10期)2015-12-07
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年6期)2015-07-01
癌变·畸变·突变(2015年3期)2015-02-27
现代检验医学杂志(2015年5期)2015-02-06
郑州大学学报(理学版)(2014年2期)2014-03-01
