
青海海北3种典型高寒草甸植物群落特征研究
2017-09-14 09:10:34井向前王瑞红任德智白玛玉珍周尧治
草地学报 2017年1期
井向前,王瑞红,任德智,白玛玉珍,潘 刚,周尧治*
(1.高原生态研究所,西藏 林芝860000 2.西藏大学农牧学院,西藏 林芝860000)
高寒草甸是适应高原高寒环境条件而形成的植被类型,其在青藏高原具有很强的典型性[1],具有双重地带性的典型植被,即高原地带性和山地垂直地带性[2],也是畜牧业的主要物质基础。近年来由于畜牧业的发展,高寒草甸超载放牧,导致草地生态系统发生不同程度的恶化,表现为草地退化面积增大,物种多样性减少,草甸初级生产力下降,草地生态环境日益恶化[3]。目前有关高寒草地多样性的研究多集中在草地荒漠化和不同草地植物群落特征[4]及不同放牧强度下高寒草地的退化过程等方面。随着放牧强度的增加,物种多样性和丰富度指数逐渐降低,草地初级生产力也随之降低。植被群落特征是反映草地生产状况的重要指标,尤其是植物多样性和初级生产力是权衡草地生产状况及其趋势的重要因子。本文通过对高寒草地不同植物群落特征及多样性进行比较,旨在了解高寒草地不同植物群落的植被现状,为研究区域内不同草地植物群落保护和进一步利用提供依据。
1 材料与方法
1.1 研究区概况
门源县位于中国青海省东北部,属海北藏族自治州管辖区域,在海北藏族自治州东部,面积为6896km2。青海省门源县门源马场,N 37.5°~38°,E 102°~102.5°。区内以高原亚寒带气候为主,干湿季分明,冬季寒冷干燥,夏季凉爽湿润。全年日照时数2264.8~2739.8h,气温日较差11.6~17.5℃,年平均气温0.8℃,年平均降水520mm。
1.2 试验设计及调查地点
在门源马场干柴滩、风匣口南滩和口门子分别选择1个草地,共3个,于2010年植物生长季后期进行调查,每个草地各设5个调查样地,共15个。根据草地情况,在每个样地设置一条100m样线,沿样线设置10个1m×1m的小样方,共计150个小样方。植被调查以样方为单位,估测整个群落的盖度,同时用钢卷尺测量群落内植物的高度,并统计植物的多度;剪取地上生物量,装入自封袋,测定鲜重。将取回的地上生物量在烘箱内以65℃烘干至恒重,称取干重。地下生物量的测定采用土柱法取样。地下生物量样方设在地上的生物量样方内,共3次重复。每个样方分4层取样,分别为0~10cm,10~20cm,20~30cm,30~40cm。样品按层标注装入土壤袋中,带回实验室冲洗、分离,然后在65℃烘箱内烘干至恒重,及时称其干重。
1.3 多样性指数计算
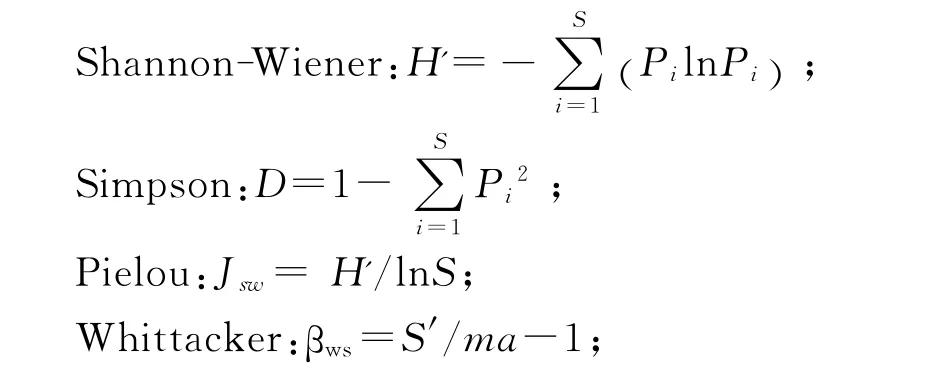
式中Pi为群落内第i种物种的重要值,S为样地中的物种数。ma为各样方或样本的平均物种数。
草本植物重要值计算公式:Pi=(相对盖度+相对高度+相对多度)/3。
1.4 数据分析
数据来源于中国科学院海北高寒草甸生态系统定位研究站2010年调查数据,采用origin9.0软件进行地上、地下生物量分布分析。
2 结果与分析
2.1 不同高寒草甸植物群落的组成与结构
青海海北不同高寒草甸群落组成存在明显差异,矮嵩草草甸植物群落的优势种为垂穗披碱草(Elymusnutans)和矮嵩草(Kobresiahumilis);金露梅草甸植物群落的优势种为金露梅(Potentillafruticosa)和溚草(Koeleriasp.);门源嵩草草甸群落的优势种为门源嵩草(Kobresiamenyuanica)和矮嵩草(Kobresiahumilis)。矮嵩草草甸植物伴生种以禾本科的优良牧草羊茅(Festucaovina)为主,金露梅草甸以莎草科的优良牧草线叶嵩草(Kobresiacapillifolia)居多,门源嵩草草甸以禾本科的异针茅(Stipaaliena)为主,亦为优良牧草(表1)。植物物种数表现为矮嵩草草甸>金露梅草甸>门源嵩草草甸。3种群落间植被高度差异较大,表现为矮嵩草草甸>金露梅草甸>门源嵩草草甸;3种草甸群落盖度均大于90%,群落垂直结构均为复层。
2.2 青海海北高寒草甸植物群落α和β多样性
青海海北不同类型高寒草甸植物群落多样性存在明显差异(表2)。均匀度指数表现为矮嵩草草甸和门源嵩草草甸明显大于金露梅草甸。3种高寒草甸植物群落多样性指数与均匀度指数的相关系数分别为0.6502,0.9371和0.8729;多样性指数与丰富度指数的相关系数分别是-0.7214,-0.7259和-0.7081,3种高寒草甸植物群落的β多样性指数均无显著性差异。

表1 青海海北高寒草甸植物群落特征Table 1 Community characteristics of alpine meadow in Haibei region of Qinghai province

表2 青海海北高寒草甸植物群落α和β多样性Table 2 αandβdiversity of alpine meadow communities in Haibei region of Qinghai province
2.3 不同高寒草甸植物群落地上生物量分布趋势
青海海北不同高寒草甸群落的地上生物量差异较为明显(图1),矮嵩草高寒草甸的地上生物量远大于金露梅高寒草甸和门源嵩草高寒草甸(P<0.05),整体表现为矮嵩草高寒草甸地上生物量分别是金露梅高寒草甸和门源嵩草高寒草甸的1.62和1.69倍。
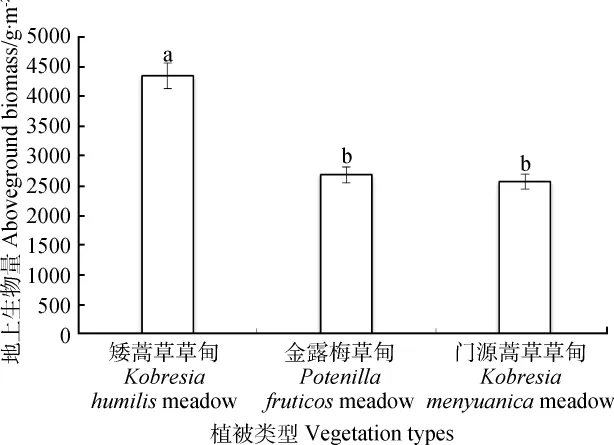
图1 青海海北高寒草甸植物群落地上生物量Fig.1 Aboveground biomass of alpine meadow in Haibei region of Qinghai province
2.4 不同高寒草甸植物群落地下生物量的垂直分布特征
青海海北不同高寒草甸植物群落的地下生物量随土层深度的变化而变化(表3),其地下生物量主要分布在土壤表层(0~10cm)。矮嵩草草甸的地下生物量为2842.62g·m-2,其中0~10cm土层生物量约占地下总生物量的77.44%。金露梅草甸的地下生物量为3241.79g·m-2,0~10cm土层生物量约占地下总生物量的54.02%。门源嵩草草甸地下生物量为4740.77g·m-2,其中0~10cm土层生物量约占地下总生物量的78.73%。他们的垂直空间分布结构表现为总的呈T字形趋势,锯齿状分布(图2)。
2.5 不同高寒草甸植物群落地上、地下生物量之间的关系
由表4可知,矮嵩草草甸、金露梅草甸和门源嵩草草甸的地上和地下生物量呈负相关关系。3种草甸群落地下、地上生物量的比值分别为0.65,1.21和1.85。
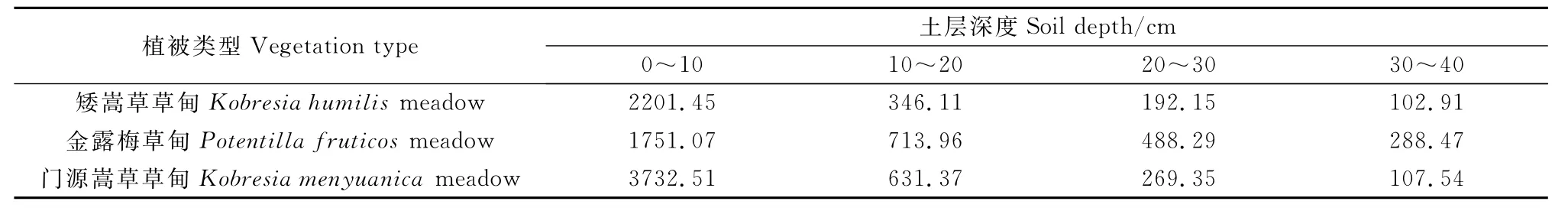
表3 高寒草甸植物群落不同土壤深度地下生物量Table 3 The belowground biomass of different soil depths of alpine meadow communities/g·m-2
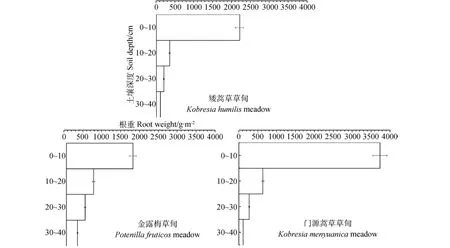
图2 青海海北地区主要植物群落地下生物量垂直分布Fig.2 The vertical distribution of belowground biomass of dominant vegetation communities in Haibei region of Qinghai province

表4 不同草甸植物群落地上、地下生物量相关性Table 4The correlation of aboveground and belowground biomass of communities
3 讨论
高寒草甸群落特征和生产力均随环境的变化而变化[5]。青海海北地区门源嵩草草甸植物群落上层为禾本科牧草,下层为嵩草;而金露梅草甸植物群落上层为金露梅,下层为苔藓,3种高寒草甸植物群落呈明显的双层结构,3种高寒草甸的气候特征具有一定的相似性,但其初级生产力差异较大,主要原因是植物群落的优势种和主要伴生种存在差异。因此,应因地制宜地制订青海海北地区不同草地类型经营策略。
本文采用α多样性指数测定青海海北地区高寒草甸的植物多样性,发现植物多样性指数和均匀度指数均表现为矮嵩草和门源嵩草草甸植物群落大于金露梅草甸植物群落,该地区的物种既包括高原高海拔地区特有的物种,如委陵菜属植物,又有喜湿、热的植物,这是由于纬度,水热条件和海拔等过渡性的生境条件为多物种的生存和发展提供了资源基础。青海海北地区不同高寒草甸植物群落间共有物种较少,物种替代速率较高。
青海海北地区高寒草甸植物群落地下生物量占总生物量的比例非常大,且地下、地上生物量的比值越大,地上生物量越低。矮嵩草草甸植物群落较高的生物多样性维持了较高的初级生产力,这与Isbell等[6]的研究结果具有一定的一致性,即植物物种的多样性是草地生态系统维系高产服务的支持者。
4 结论
青海海北地区3种典型高寒草甸植物群落的垂直结构均为复层;其多样性指数与均匀度指数关系较其与丰富度指数的关系更为密切;不同高寒草甸植物群落内样方间共有物种较少,物种替代速率较高;矮嵩草高寒草甸地上生物量远高于金露梅高寒草甸和门源嵩草高寒草甸;其地下生物量主要分布在土壤表层,垂直空间分布结构呈T字形和锯齿状;其地下生物量所占总生物量的比例非常大,且地下、地上生物量呈负相关关系。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
矿产勘查(2021年3期)2021-07-20 08:01:36
中国水土保持(2020年11期)2020-11-19 01:52:16
党的生活·青海(2019年12期)2019-12-23 08:28:56
活力(2019年21期)2019-04-01 12:17:10
岷峨诗稿(2017年4期)2017-04-20 06:26:36
中国卫生(2016年8期)2016-11-12 13:27:04
黄河之声(2016年24期)2016-02-03 09:01:52
