
镉胁迫对红椿(Toonaciliate Roem.)幼苗生长及碳、氮、磷、钾累积与分配的影响
2017-09-03 08:57:10王岑涅刘柿良李勋王丽萍张健
农业环境科学学报 2017年8期
王岑涅,刘柿良,李勋,王丽萍,张健,4*
(1.四川农业大学生态林业研究所,四川省林业生态工程重点实验室,成都 611130;2.成都大学旅游与经济管理学院,成都 610106;3.四川农业大学风景园林学院,成都 611130;4.长江上游生态安全协同创新中心,成都 611130)
镉胁迫对红椿(Toonaciliate Roem.)幼苗生长及碳、氮、磷、钾累积与分配的影响
王岑涅1,2,刘柿良3,李勋1,王丽萍1,张健1,4*
(1.四川农业大学生态林业研究所,四川省林业生态工程重点实验室,成都 611130;2.成都大学旅游与经济管理学院,成都 610106;3.四川农业大学风景园林学院,成都 611130;4.长江上游生态安全协同创新中心,成都 611130)
为了解镉(Cd)胁迫对红椿(Toona ciliata Roem.)生长及养分[碳(C)、氮(N)、磷(P)和钾(K)]吸收的影响,采用盆栽控制试验研究了不同浓度Cd处理[0(对照)、10、20、40、80、160 mg·kg-1]对其幼苗生长发育、生物量以及养分积累与分配特征的影响。结果显示:与对照相比,红椿幼苗生长特性指数(如叶数、叶长、叶宽、根长及地径和株高)与根、茎、叶生物量随Cd浓度增加而降低,但低Cd处理(<40 mg·kg-1)对各器官指数、株高及总生物量无显著影响(P>0.05)。随着Cd胁迫浓度增加,根、茎、叶中Cd浓度逐渐升高,且根大于茎;根K、茎K、叶K和叶N累积量随胁迫浓度增加呈先升后降,而根C、根N、茎N、根P和叶P累积量则逐渐降低。此外,茎C、叶C和茎P在低Cd处理时无显著差异,但较高Cd胁迫处理(≥40.00 mg·kg-1)则显著抑制并改变其累积与分配格局。红椿幼苗具有一定的抗Cd胁迫能力,但较高浓度Cd胁迫(≥40.00 mg·kg-1)显著影响了红椿幼苗的生长特性及其养分格局。
镉胁迫;生长发育;养分累积;红椿;生物量
土壤重金属污染已经成为危害全球生物圈的重大因素,其中土壤镉(Cd)的污染受到广泛关注[1-3]。土壤较低的Cd浓度即可对植物的生长和发育造成危害[4-5]。尽管有研究表明低浓度Cd有促进植物生长的作用[6-8],但大多数研究显示一定浓度(程度)的Cd胁迫会阻碍植物的生长[7,9]。同时,Cd会影响土壤中养分的有效性,影响植物对土壤养分的吸收利用[10-11],进而形成Cd与养分同时限制植物生长的恶性循环。目前的研究大多关注Cd在植物-土壤系统间的富集、迁移和转化过程及其对植物生理过程的影响[12-15],但由于所用材料、处理方式和浓度设置不尽相同,其各异结果导致Cd胁迫对植物养分作用机理尚不清楚。
乡土木本树种具有生物量高、生长迅速、栖息地适应能力强等优势[16-17],而迄今为止尚无相关研究关注其在Cd污染条件下的生长适应特征。红椿(Toona ciliata Roem.)作为我国国家级(Ⅱ)重点保护植物,也是我国(亚)热带地区的珍贵速生用材树种,目前陷于濒临灭绝的境地,但对其研究仅有零星报道,如关于其天然居群遗传结构与体内化学成分分析等研究[18-19]。红椿作为长江上游重要的速生树种,先前研究报道其对土壤(紫色土与冲击土)重金属铅(Pb)具有较强耐受性和积累能力[17],而长江上游典型紫色土和冲积土受到重金属污染日趋严重[20-21],红椿是否对长江上游典型土壤中大量存在的重金属Cd同样具有较强的耐(抗)性却未见报道。因此,本研究以红椿(T.ciliata)幼苗为试验材料,采用盆栽控制试验,研究不同Cd胁迫条件下红椿幼苗生长以及对养分(C、N、P和K)的积累、分配与利用特征,分析不同Cd胁迫水平对红椿生长适应与养分利用机制的影响,了解红椿对Cd污染的耐受性,以期为深入认识Cd污染条件下植物生长适应过程提供基础数据。
1 材料与方法
1.1 试验地与材料
试验设置在四川农业大学成都校区实验基地温室(30°38′N,103°45′E),海拔510~600 m,属亚热带湿润气候区,四季分明,日照偏少,雨量充沛,湿度较大[3]。供试红椿幼苗为楝科(Meliaceae)香椿属(Toona Roem.)植物,本研究以四川省宜宾市种苗站内生长环境相同且长势一致的一年生红椿种苗(株高约40 cm、地径约8 mm)为供试植物。试供土壤为黄壤,采自雅安市四川农业大学读书公园,其基本理化性质见表1。
1.2 试验设计
2013年3月,将试供土壤风干,研磨,分别经5 mm和2 mm网格过筛,静置,按每盆10 kg(干土)装入塑料花盆(37 cm×25 cm×27 cm),置于大棚中(透光率为80%,温度为27℃±2℃,相对湿度为75%±3%)待用。2013年6月,选取粗细均匀、生长良好的1年生半木质化红椿幼苗移栽至塑料盆,每盆种植1株,待植物恢复生长后,于2014年5月10日进行不同浓度Cd处理试验。Cd浓度梯度设置为:Control(0 mg· kg-1)、T1(10 mg·kg-1)、T2(20 mg·kg-1)、T3(40 mg·kg-1)、T4(80 mg·kg-1)和T5(160 mg·kg-1),共35盆(包括初始测定5盆),每处理5次重复。Cd添加方法:以CdCl2与蒸馏水配制成约500 mL溶液均匀施入相应塑料盆中,渗出液反复回收浇灌,直到Cd离子与土壤均匀混合。试验期间,对所有供试植株进行统一浇水,除草防虫和防病等管理,控制浇水量以避免多余水分流出塑料盆,且不喷施农药与追施化肥。

表1 试供土壤中营养元素、有机质、导电性及Cd的背景值Table 1 The background values of nutrient elements,organic matter,and electric conductivity(EC)in the studied soil
1.3 样品分析
2014年9月10日,测量所有植株的叶长、叶宽、地径、主根长及地上部高。采用破坏性收获法采集所有植株(除用于初始测定植株),并将样品分为根、茎、叶三部分,先后多次用Na2-EDTA(祛除表面附着离子)和去离子水洗净,105℃烘箱内杀青30 min,75℃烘干至恒重,计算其根、茎、叶及单株生物量。将样品(根、茎、叶)粉碎研磨过1 mm筛,待测。
植株全C采用K2Cr2O7氧化-FeSO4滴定法[22]测定,为保证有机C氧化完全,样品测定时滴定所用FeSO4标准液体积小于空白标定所消耗体积1/3时,需要减少称取样品量而重做。样品N和P含量测定先采用H2SO4-H2O2消煮。靛酚蓝比色法[22]测定N含量,平均回收率99.87%:试验中选择690 nm为测定波长,比色在显色1~6 h内进行;标准溶液H2SO4浓度控制在0.7 mol·L-1左右,待测(标准)液氮浓度控制在0~300 μg·mL-1,不允许有H2O2残存。P含量测定则采用钼蓝法[23]:在根系、茎干和叶片的干样中加入4 mL HNO3和1 mL H2O2进行消解,450 nm波长下比色,将溶液中残留的H2O2加热分解,避免影响比色测定。按照Hernández等[24]的方法对干样消解测定K含量,其平均回收率为89.05%。植株Cd含量采用湿样消解法[25]消解,原子吸收分光光度计(AA320N型,上海精密科学仪器有限公司)测定,Cd测定精度(RSD)为1.97%。试验选择灰化温度为560℃,标准加入法测定Cd平均回收率为92.43%。
1.4 数据分析与处理
根据试验初期红椿幼苗各器官生物量和实验结束时相应器官生物量计算不同Cd处理条件下红椿各器官生物量增幅;根据试验期间各器官生物量增幅与C、N、P和K浓度计算红椿幼苗C、N、P和K累积量(即各器官生物量与C、N、P和K浓度之积)与分配特征。单因素方差检验(One-way ANOVA,Tukey检验)用于比较不同Cd处理条件下红椿幼苗的生长特征,生物量增幅和C、N、P和K累积与分配差异,显著性水平设定a=0.05,统计分析均使用SPSS 17.0软件。
2 结果与分析
2.1 Cd处理对红椿幼苗组织Cd浓度的影响
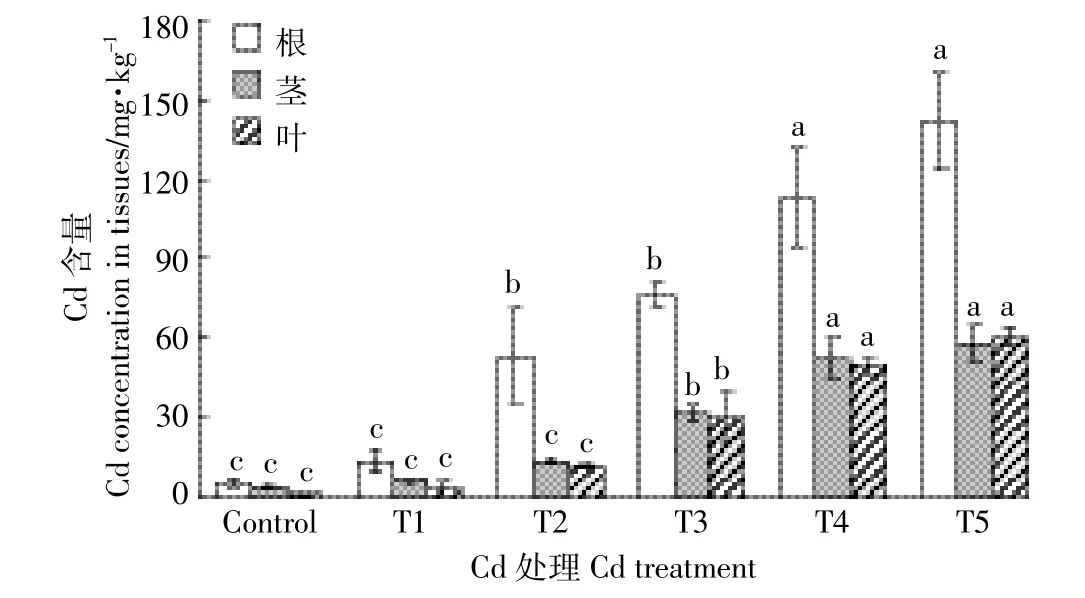
图1 Cd处理对红椿幼苗根、茎、叶中Cd浓度的影响Figure 1 Effect of cadmium concentrations of roots,stem,and leaves of Toona ciliata seedlings under different cadmium concentration treatments
图1显示,红椿各器官中Cd浓度随着Cd处理浓度的增加而显著增加(P<0.05)。与对照相比,T1处理并未显著改变根系(13.66 mg·kg-1)、茎干(6.50 mg· kg-1)和叶片(3.13 mg·kg-1)中Cd浓度(P>0.05);随着Cd处理浓度增大,T5处理下根系(141.47 mg·kg-1)、茎干(57.78 mg·kg-1)和叶片(60.23 mg·kg-1)中Cd浓度较对照分别显著增加27.43、13.49、53.42倍。同时,幼苗各器官中Cd浓度在T4和T5处理间无显著差异,叶片和茎干中Cd浓度在T1、T2与对照处理间亦无明显差异(P>0.05)。相同浓度Cd处理下,根系中Cd浓度显著大于茎干且大于叶片。
2.2 Cd处理对红椿幼苗生长特性及生物量的影响
由表2和表3可见,不同浓度Cd处理对红椿幼苗生长特性与生物量的影响显著(P<0.05)。与对照处理相比,10 mg·kg-1Cd胁迫(T1)对叶数、根长、株高及根、茎、叶生物量无显著影响(P>0.05),而叶长、叶宽和地径较对照分别降低7.18%、11.48%和7.38%(P<0.05)。随着Cd处理浓度增加,叶宽、叶长和叶生物量呈先升后降,在T2处理时达到最高值,较T1处理分别提升了1.43%(P<0.05)、1.06%(P>0.05)和3.27% (P>0.05)。同时,T5处理下叶数、根长、株高及根、茎和总生物量较T1处理分别显著降低40.15%、13.18%、9.66%、20.13%、14.92%和15.60%,R/S比虽未随Cd浓度增加呈现规律性逐渐降低趋势,但其在T5处理下也比T1处理降低5.85%,而地径随Cd处理浓度增加无明显变化(P>0.05)。
2.3 Cd处理对红椿幼苗组织C、N、P、K浓度的影响
图2所示,不同浓度Cd处理在不同程度上影响了幼苗根系、茎干和叶片中C、N、P和K浓度。随着Cd浓度增加,根K、叶K和叶N浓度呈先升后降趋势,在20 mg·kg-1Cd胁迫(T2)下达到最高值,较对照
分别升高96.06%、17.68%和44.28%,而后逐渐降低,到T5处理时呈最低值。相似地,叶C、茎K、根N和茎P浓度在对照、T1和T2处理间无明显差异(P>0.05),之后随Cd浓度增加而逐渐降低。然而,不同浓度Cd处理对根C与茎C浓度无显著影响(P>0.05)。

表2 Cd处理对红椿幼苗生长特性的影响Table 2 Effect of growth character of Toona ciliate seedlings under different cadmium concentration treatments
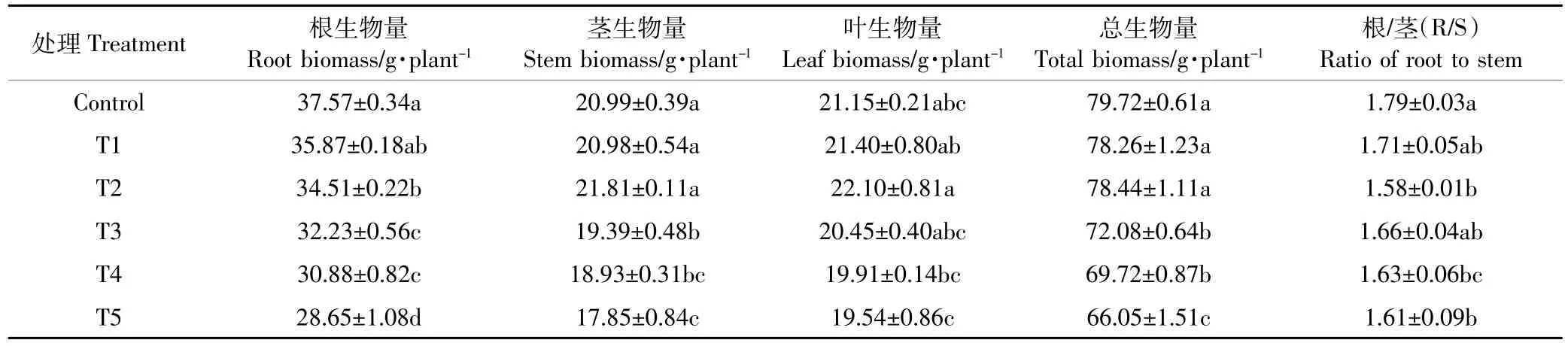
表3 Cd处理对红椿幼苗各器官生物量及根/茎生物量(R/S)的影响Table 3 Effect of biomass of Toonaciliate seedlings under different cadmium concentration treatments

图2 Cd处理对红椿幼苗根、茎、叶中C(A)、K(B)、N(C)和P(D)浓度的影响Figure 2 Effect of C(A),K(B),N(C)and P(D)concentrations of roots,stems,and leaves of Toona ciliata seedlings under different cadmium concentration treatments
2.4 Cd处理对红椿幼苗组织C、N、P、K累积及分配的影响
不同浓度Cd处理显著影响幼苗各器官中C、N、P和K的积累量(图3)。与对照处理相比,10 mg·kg-1Cd胁迫处理(T1)显著(P<0.05)升高根K和叶N累积量,分别为68.89%和35.68%,却显著降低茎N、根P和叶P累积量分别为16.26%、22.46%和25.15%(P<0.05)。随Cd浓度增加,根K、茎K、叶K和叶N累积量均呈先升后降(T2最高),随后逐渐降低并在T5处理达到最低值。根C、根N、茎N、根P和叶P累积量均随Cd浓度增加而降低(T5最低)。茎C、叶C和茎P在对照、T1与T2间无显著差异(P>0.05),之后随Cd浓度增大而降低。同时,不同浓度Cd处理明显改变了根系、茎干和叶片中C、N、P和K分配(图4)。随Cd浓度增加,叶N、茎P和根K积累量比例呈现先升后降,根N、茎N、根P、茎K和叶K积累量比例则相反;而叶C积累量比例逐渐增大,叶P积累量比例逐渐降低。同时,C和P累积在根系中的比例显著大于其地上部(茎干+叶片),而N和K在地上部中累积比例高于根系。

图3 Cd处理对红椿幼苗根、茎、叶中C(A)、K(B)、N(C)和P(D)积累量的影响Figure 3 Effect of C(A),K(B),N(C)and P(D)accumulations of roots,stem,and leaves of Toona ciliata seedlings under different cadmium concentration treatments
3 讨论
3.1 Cd处理与红椿幼苗组织Cd浓度的关系
本研究中,不同浓度Cd处理下红椿幼苗根系中Cd浓度显著高于其地上部浓度(图1),表明根系对Cd富集能力高于地上部,与先前大多数研究结果一致[7,12,16]。这主要是因为植物吸收的Cd作为带正电荷的金属离子极易与组织中带负电荷化合物结合,从而限制部分Cd向地上部(茎干和叶片)运输而累积在根系[13];同时,植株根系对重金属离子具有截留作用,借助细胞质子泵产生的能量“主动地”将根细胞中的生物大分子与Cd离子结合成稳定螯合物[26-28]。这也被认为是植物对逆境胁迫的生存反应。
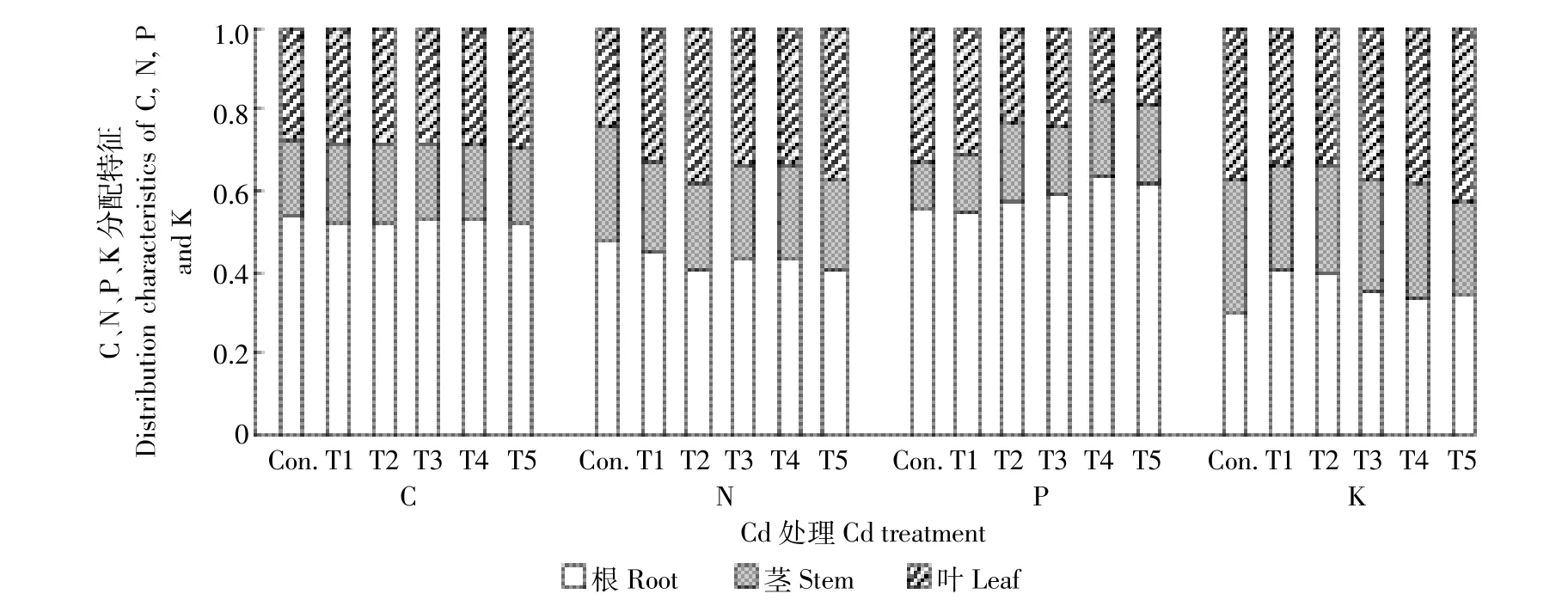
图4 Cd处理下红椿幼苗根、茎、叶中C、N、P和K分配特征Figure 4 Distribution characteristics of C,N,P and K in each organ to total plant of Toona ciliata under different cadmium concentrations
3.2 Cd处理与红椿幼苗生长及其Cd浓度的关系
植物形态上的改变通常也是其适应逆境的基本机制。Cd胁迫常常限制土壤养分的有效性和可利用性,加大植物各器官之间对资源的竞争,进而表现出植物各器官生物量及其分配格局的改变[29]。本研究中,低Cd浓度处理(<40 mg·kg-1)对红椿幼苗的生长特性(叶数、根长和株高)与生物量(茎、叶和总生物量)无显著影响(表2、表3),但高浓度Cd处理(≥40 mg·kg-1)则抑制其叶片的生长特性(如叶长、叶宽和叶数)、降低植株高度以及生物量。这些研究结果与先前研究类似[27],表明植物在Cd胁迫下均表现出生长受阻的现象。
植物通常以改变生物量分配与利用的方式,将有限资源分配到更有效器官而更利于适应逆境[21,30]。本研究中,低浓度Cd处理(<40 mg·kg-1)对红椿幼苗根、茎、叶生物量及其分配格局改变并不显著(P>0.05),而较高浓度Cd处理(≥40 mg·kg-1)则降低其生物量增幅(表3),表明Cd能够有效地限制土壤养分利用效率,加大植物器官对养分资源的竞争,进而改变红椿幼苗各器官的生物量与分配格局[27,31]。另一方面,在高Cd浓度处理(160 mg·kg-1)时,红椿仍然最大限度地增加叶片比例,加大光合产物量以维持生存,这是植物在恶劣条件下的适应机制;而低浓度Cd胁迫并没有显著改变其生物量及其分配格局,则证明了其对Cd污染具有抗性。R/S作为植物获取有限土壤资源维持生长的重要指标,比值越高表明其抗性越强[14]。本研究中,Cd处理降低了红椿幼苗R/S(表2),与对桂花(Osmanthus fragrans)的研究结果相悖[32],而随着Cd浓度增加变化却不显著,其原因还有待进一步研究。
3.3 Cd处理与红椿幼苗养分累积与分配的关系
均衡的营养元素是植物正常生长发育的前提,但重金属能够改变土壤环境,进而影响植物对土壤养分的吸收与利用[33]。C是植物干物质重要组成元素,C平衡程度决定着其受害程度[34]。Elser等[35]对全球492种陆地植物叶片研究显示,其C浓度为(464±32.1)mg·g-1。本研究中,红椿幼苗在正常条件下(Control)叶C浓度为(368.79±6.66)mg·g-1(图2),表明其本身具有较强的C储存能力。随着Cd胁迫浓度的增加,茎C浓度不同程度降低,而根C和叶C浓度却未见显著变化(图2A)。试验中大量累积的Cd破坏了根、茎细胞透性,扰乱对养分元素吸收、利用,可能是由于植物将吸收的N首先利用在叶片生产以维持最大固C率[36]。
本研究中较高浓度Cd胁迫(≥40 mg·kg-1)导致了叶N浓度逐渐降低(图2C),表明Cd胁迫已经限制了植物对N元素的吸收,进而影响其光合生理过程,抑制植物生长发育。同时,高浓度Cd处理显著降低植物各器官的P浓度,也可能与Cd胁迫条件下植物养分吸收受限有关(图2D)。随着Cd浓度增加,根N和茎N浓度均逐渐降低,其原因是Cd影响植物体内N代谢限速酶活性而抑制土壤中N元素的吸收,与对桂花(O.fragrans)的研究结果一致[32]。另一方面,由于根系与茎干为植物中储存N最集中的部分,较低浓度Cd处理(<40 mg·kg-1)使植物吸收、利用与分配N元素达到相对平衡,而对生物量及其对C的分配影响不大(图2);相反,较高浓度Cd处理(≥40 mg·kg-1)显著地抑制植物根生长,降低根生物量比例,增加叶片生物量比例(图3、图4),同时也降低了各器官对N的吸收(图2C)。这种N吸收的减少可能也造成细胞内阳离子有效浓度的降低[17],进而增加了Cd的毒性,导致根生物量比例的降低。因此,Cd胁迫对N代谢与利用等生理过程的影响可能是植物在Cd胁迫条件下生理响应特征的主要原因。
P在ATP反应、糖类代谢、蛋白质代谢以及脂类代谢中起重要作用[4],并对细胞分裂和植物各器官分化发育,特别是开花结实具有重要作用。本研究中高浓度Cd处理显著降低植物各器官P浓度也可能与Cd胁迫条件下植物养分吸收受限有关。因此,证实了一定程度的Cd胁迫将会导致土壤养分(如P)成为限制植物生长的重要因子之一[15]。另一方面,由于P能够与土壤中存在的Cd形成难溶化合物,调节Cd在土壤中的生物有效性[13],从而影响植物对养分(包括P)的吸收与利用。目前,虽然具体的机制仍然不甚清晰,但这为后续研究提供了有益的思路。
K作为一种重要的抗逆元素对植物细胞渗透压和膨压的调节具有重要作用。本研究中低浓度Cd处理(<40 mg·kg-1)增加了根K和叶K浓度与累积量,而在高浓度Cd处理(≥40 mg·kg-1)时其含量却显著降低(图2~图4),表明介质中Cd2+剂量与植物对K+的吸收和运输相关,且Cd2+可能与K+存在某种形式的交互作用。这与Hernández等[24]的研究结果相类似,但其主要机理还有待进一步研究探索。
植物器官养分积累与分配格局能够响应环境因子的变化[32]。一种环境刺激机制在最初使得植物形成一种保护特性进而促进系列生理反应,而随着环境胁迫程度增加,这种保护性会随着植物伤害增加而消失,从而导致植物生长下降[8,15]。本研究中较低浓度Cd处理并未明显改变幼苗叶C积累量(图3A),且可在轻度Cd污染条件下很好地生长;相对较低浓度Cd处理则仅对根C和N积累量无显著影响(P>0.05),表明Cd导致的促进植物生长的机制主要作用于根系活动;而高浓度Cd处理抑制了各器官C、N和P的累积,表明高浓度Cd污染可以限制植物对土壤养分的吸收、同化和积累。
4 结论
较高浓度Cd处理(≥40 mg·kg-1)抑制了红椿幼苗生长与生物量产生与累积,同时也改变了植物体内C、N、P、K的累积与分配,但低Cd对其并无显著影响;在80~160 mg·kg-1Cd处理时,根系中Cd浓度超过了100.00 mg·kg-1,较地上部更具Cd积累能力;在160 mg·kg-1重度Cd处理时,红椿仍能通过改变养分累积与利用特征维持生物量生产。尽管这些结果无法完全揭示Cd胁迫对红椿生长、养分吸收、积累与利用的影响机制,但为进一步探讨Cd胁迫下植物生长与适应机制提供了良好的研究基础。
[1]Zacchini M,Pietrini F,Mugnozza G S,et al.Metal tolerance,accumulation and translocation in poplar and willow clones treated with cadmium in hydroponics[J].Water,Air&Soil Pollution,2009,197(1/2/3/4):23-34.
[2]Liu S L,Yang R J,Ma M D,et al.Effects of exogenous NO on the growth, mineral nutrient content,antioxidant system,and ATPase activities of Trifolium repens L.plants under cadmium stress[J].Acta Physiologiae Plantarum,2015,37(1):1-16.
[3]Liu S L,Yang R J,Pan Y Z,et al.Nitric oxide contributes to minerals absorption,proton pumps and hormone equilibrium under cadmium excess in Trifolium repens L.plants[J].Ecotoxicology and Environmental Safety,2015,119(1):35-46.
[4]Nedjimi B,Daoud Y.Cadmium accumulation in Atriplex halimus subsp. schweinfurthii and its influence on growth,proline,root hydraulic conductivity and nutrient uptake[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2009,204(4):316-324.
[5]Benavides M P,Gallego S M,Tomaro M L.Cadmium toxicity in plants[J]. Brazilian Journal of Plant Physiology,2005,17(1):21-34.
[6]Boussama N,Ouariti O,Ghorbal M H.Changes in growth and nitrogen assimilation in barley seedlings under cadmium stress[J].Journal of Plant Nutrition,1999,22(4/5):731-752.
[7]刘柿良,潘远智,杨容孑,等.外源一氧化氮对镉胁迫下长春花质膜过氧化、ATPase及矿质营养吸收的影响[J].植物营养与肥料学报, 2014,20(2):445-458.
LIU Shi-liang,PAN Yuan-zhi,YANG Rong-jie,et al.Effects of exogenous NO on mineral nutrition absorption,lipid peroxidation and ATPase of plasma membrane in Catharanthus roseus tissues under cadmium stress[J].Journal of Plant Nutrition and Fertilizer,2014,20(2):445-458.
[8]Chien H F,Kao C H.Accumulation of ammonium in rice leaves in response to excess cadmium[J].Plant Science,2000,156(1):111-115.
[9]Liu S L,Yang R J,Pan Y Z,et al.Exogenous NO depletes Cd-induced toxicity by eliminating oxidative damage,re-establishing ATPase activity,and maintaining stress-related hormone equilibrium in white clover plants[J].Environmental Science and Pollution Research,2015,22(21):16843-16856.
[10]Cuypers A,Smeets K,Ruytinx J,et al.The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings[J].Journal of Plant Physiology,2011,168(4):309-316.
[11]Kovácˇik J.Hyperaccumulation of cadmium in Matricaria chamomilla:A never-ending story?[J].Acta Physiologiae Plantarum,2013,35 (5):1721-1725.
[12]杨容孑,刘柿良,宋会兴,等.不同氮形态对龙葵Cd积累、抗氧化系统和氮同化的影响[J].生态环境学报,2016,25(4):715-723.YANG Rong-jie,LIU Shi-liang,SONG Hui-xing,et al.Impacts of different nitrogen forms on cadmium accumulation,antioxidant system and nitrogen assimilation in hyperaccumulator Solanum nigrum L.[J]. Ecology and Environmental Sciences,2016,25(4):715-723.
[13]Naresh K R,Nagendran R.Fractionation behavior of heavy metals in soil during bioleaching with Acidithiobacillus thiooxidans[J].Journal of Hazardous Materials,2009,169(1/2/3):1119-1126.
[14]Wang C Q,Song H.Calcium protects Trifolium repens L.seedlings against cadmium stress[J].Plant Cell Reports,2009,28(9):1341-1349.
[15]Liu S L,Yang R J,Pan Y Z,et al.Beneficial behavior of nitric oxide in copper-treated medicinal plants[J].Journal of Hazardous Materials, 2016,314(1):140-154.
[16]田如男,袁安全,薛建辉.4种常绿阔叶乔木树种幼苗抗铅胁迫能力的比较[J].南京林业大学学报(自然科学版),2005,29(6):81-84.
TIAN Ru-nan,YUAN An-quan,XUE Jian-hui.Comparison on the ability of resistance to Pb stress of four evergreen broadleaved trees seedlings[J].Journal of Nanjing Forestry University(Natural Sciences), 2005,29(6):81-84.
[17]胡方洁,张健,杨万勤,等.Pb胁迫对红椿(Toona ciliata Roem.)生长发育及Pb富集特性的影响[J].农业环境科学学报,2012,31 (2):284-291.
HU Fang-jie,ZHANG Jian,YANG Wan-qin,et al.Effects of Pb stress on the growth,development and Pb enrichment properties of Toona ciliata Roem.saplings[J].Journal of Agro-Environment Science,2012, 31(2):284-291.
[18]刘军,陈益泰,孙宗修,等.基于空间自相关分析研究毛红椿天然居群的空间遗传结构[J].林业科学,2008,44(6):45-52.
LIU Jun,CHEN Yi-tai,SUN Zong-xiu,et al.Spatial genetic structure of Toona ciliata var.pubescens populations in terms of spatial autocorrelation analysis[J].ScientiaSilvae Sinicae,2008,44(6):45-52.
[19]卢海啸,李家洲,莫花浓,等.红楝子枝叶化学成分研究[J].中药材, 2009,32(10):1539-1542.
LU Hai-xiao,LI Jia-zhou,MO Hua-nong,et al.Study on chemical constituents of tress of Toona ciliata[J].Journal of Chinese Medicinal Material,2009,32(10):1539-1542.
[20]刘小诗,李莲芳,曾希柏,等.典型农业土壤重金属的累积特征与源解析[J].核农学报,2014,28(7):1288-1297.
LIU Xiao-shi,LI Lian-fang,ZENG Xi-bai,et al.Characterization of heavy metal accumulation in typical agriculture soils and its source analysis[J].Journal of Nuclear Agricultural Sciences,2014,28(7):1288-1297.
[21]Liu S L,Luo Y M,Yang R J,et al.High resource-capture and-use efficiency,and effective antioxidant protection contribute to the invasiveness of Alnus formosana plants[J].Plant Physiology and Biochemistry, 2015,96(1):436-447.
[22]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社, 2000.
LU Ru-kun.Methods for chemical analysis of soil agriculture[M].Beijing:China Agricultural Science and Technology Press,2000.
[23]Mukherjee S K,Asanuma S.Possible role of cellular phosphate pool and subsequent accumulation of inorganic phosphate on the aluminum tolerance in Bradyrhizobium japonicum[J].Soil Biology and Biochemistry,1998,30(12):1511-1516.
[24]Hernández L E,Gárate A,Carpena-Ruiz R.Effects of cadmium on the uptake,distribution and assimilation of nitrate in Pisum sativum[J]. Plant and Soil,1997,189(1):97-106.
[25]中国标准出版社.食品卫生检验方法:理化部分(一)[M].北京:中国标准出版社,2012. China National Standard Publishing House.Food hygiene inspection methods of physical and chemical components(1)[M].Beijing:China Standard Press,2012.
[26]Dahmani-Muller H,Van Oort F,Gelie B,et al.Strategies of heavy metal uptake by three plant species growing near a metal smelter[J]. Environmental Pollution,2000,109(2):231-238.
[27]Babula P,Klejdus B,Kovácˇik J,et al.Lanthanum rather than cadmium induces oxidative stress and metabolite changes in Hypericum perforatum[J].Journal of Hazardous Materials,2015,286(1):334-342.
[28]Hasan S A,Fariduddin Q,Ali B,et al.Cadmium:Toxicity and tolerance in plants[J].Journal of Environmental Biology,2009,30(2):165-174.
[29]刘柿良,杨容孑,马明东,等.土壤镉胁迫对龙葵(Solanum nigrum L.)幼苗生长及生理特性的影响[J].农业环境科学学报,2015,34(2):240-247.
LIU Shi-liang,YANG Rong-jie,MA Ming-dong,et al.Effects of soil cadmium on growth and physiological characteristics of Solanum nigrum L.plants[J].Journal of Agro-Environment Science,2015,34(2):240-247.
[30]Gonzaga M I S,Santos J A G,Ma L Q.Phytoextraction by arsenic hyperaccumulator Pteris vittata L.from six arsenic-contaminated soils:Repeated harvests and arsenic redistribution[J].Environmental Pollution,2008,154(2):212-218.
[31]Benavides M P,Gallego S M,Tomaro M L.Cadmium toxicity in plants [J].Brazilian Journal of Plant Physiology,2005,17(1):21-34.
[32]吴福忠,杨万勤,张健,等.镉胁迫对桂花生长和养分积累、分配与利用的影响[J].植物生态学报,2010,34(10):1220-1226.
WU Fu-zhong,YANG Wan-qin,ZHANG Jian,et al.Effects of cadmium stress on growth and nutrient accumulation,distribution and utilization in Osmanthus fragrans var.thunbergii[J].Chinese Journal of Plant Ecology,2010,34(10):1220-1226.
[33]Chien H F,Kao C H.Accumulation of ammonium in rice leaves in response to excess cadmium[J].Plant Science,2000,156(1):111-115.
[34]Güsewell S.N∶P ratios in terrestrial plants:Variation and functional significance[J].New Phytologist,2004,164(2):243-266.
[35]Elser J J,Fagan W F,Denno R F,et al.Nutritional constraints in terrestrial and freshwater food webs[J].Nature,2000,408(6812):578-580.
[36]Broadley M R,Escobar-Gutiérrez A J,Burns A.What are the effects of nitrogen deficiency on growth components of lettuce?[J].New Phytologist,2000,147(3):519-526.
Impacts of Cd stress on growth,and accumulation and distribution of C,N,P and K in Toona ciliata Roem. seedlings
WANG Cen-nie1,2,LIU Shi-liang3,LI Xun1,WANG Li-ping1,ZHANG Jian1,4*
(1.Key Laboratory of Ecological Forestry Engineering of Sichuan Province,Institute of Ecology&Forest,Sichuan Agricultural University, Chengdu 611130,China;2.College of Tourism and Economic Management,Chengdu University,Chengdu 610106,China;3.College of Landscape Architecture,Sichuan Agricultural University,Chengdu 611130,China;4.Collaborative Innovation Center of Ecological Security in the Upper Reaches of Yangtze River,Chengdu 611130,China)
In the presented work,the effects of different Cd stress on the growth and nutrient absorption(C,N,P,and K)of Toona ciliata Roem.seedlings were investigated.A controlled pot experiment was arranged with different treatments of six Cd dosages[control,10,20,40, 80,and 160 mg·kg-1]to investigate the growth characteristics,biomass production and nutrient accumulation and distribution in the T.ciliata seedlings.The results indicated that the growth characteristic parameters(leaf growth,leaf length,leaf width,root length,base diameter, and shoot height)and tissue biomasses(i.e.,root,stem,and leaf)declined with increasing Cd dosage,whereas the treatments with lower Cd dosage(<40 mg·kg-1)had no significant(P>0.05)influences on tissue index,plant height,and total biomass.Additionally,the Cd distribution pattern under different Cd treatments showed the same order,with root>stem and/or root>leaf.Moreover,with increasing Cd dosage,theaccumulation of root K,stem K,leaf K,and leaf N showed a trend of increase at first and then decrease,whereas accumulation of root C,root N,stem N,root P,and leaf P gradually decreased.Furthermore,the treatments with higher Cd dosages(≥40.00 mg·kg-1)significantly inhibited the accumulation of stem C,leaf C,and stem P and altered their distribution patterns,whereas treatment with lower Cd dosages(<40 mg·kg-1)did not show significant effects.Results suggest that T.ciliata could adapt efficiently to Cd-contaminated environments by altering its nutrient use strategy and biomass distribution pattern.
cadmium stress;growth and development;nutrient accumulation;Toona ciliata Roem.;biomass
X171.5
A
1672-2043(2017)08-1492-08
10.11654/jaes.2017-0011
2017-01-03
王岑涅(1985—),男,重庆开县人,在读博士,讲师,主要从事土壤污染及其修复研究。E-mail:wangcn100@163.com
*通信作者:张健E-mail:sicauzhangjian@163.com
国家自然科学基金项目(31370628);国家科技支撑计划项目(2011BAC09B05)
Project supported:The National Natural Science Foundation of China(31370628);The National Key Technology Research and Development Program of the Ministry of Science and Technology of China(2011BAC09B05)
王岑涅,刘柿良,李勋,等.镉胁迫对红椿(Toonaciliate Roem.)幼苗生长及碳、氮、磷、钾累积与分配的影响[J].农业环境科学学报,2017,36(8):1492-1499.
WANG Cen-nie,LIU Shi-liang,LI Xun,et al.Impacts of Cd stress on growth,and accumulation and distribution of C,N,P and K in Toona ciliata Roem. seedlings[J].Journal of Agro-Environment Science,2017,36(8):1492-1499.
猜你喜欢
东北林业大学学报(2022年7期)2022-08-04 01:48:02
落叶果树(2021年6期)2021-02-12 01:28:54
今日农业(2020年19期)2020-12-14 14:16:52
文苑(2020年12期)2020-04-13 00:54:10
海峡姐妹(2019年1期)2019-03-23 02:42:46
乡村科技(2018年17期)2018-08-16 02:27:50
湖北林业科技(2016年5期)2016-12-02 10:23:28
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
云南农业科技(2012年6期)2012-11-13 06:29:30
