
精氨酸对仔猪的营养调节作用及最新研究进展
2017-09-03 11:01:14毛俊舟王淑楠姜茂成
猪业科学 2017年8期
毛俊舟,王淑楠,彭 众,姜茂成
(扬州大学动物科学与技术学院,扬州 225009)
精氨酸对仔猪的营养调节作用及最新研究进展
毛俊舟,王淑楠,彭 众,姜茂成
(扬州大学动物科学与技术学院,扬州 225009)
精氨酸是目前已发现的动物体内功能最多的氨基酸。日粮中添加精氨酸能有效地调节仔猪的生长发育和肠道免疫功能。该文就精氨酸在仔猪体内的合成代谢、营养调控及免疫功能等方面进行综述。
精氨酸;生长性能;营养调控;肠道免疫
精氨酸(arginine,Arg)是一种功能性氨基酸,1886年,Schule等首次从羽扇豆幼苗提取物中分离出精氨酸,后来在哺乳动物的蛋白质中也发现了精氨酸。仔猪早期断奶有助于提高母猪繁殖力,增加母猪的产仔数,提高养殖场的经济效益。然而,新生仔猪处于幼龄阶段,早期断奶存在消化道功能紊乱、腹泻、饲料利用率低、生长缓慢等缺陷,严重影响仔猪的生长发育。而精氨酸是哺乳仔猪的必需氨基酸[1]。采食适量的精氨酸有利于恢复仔猪的生长性能,提高仔猪存活率,还能够增加仔猪肝脏热休克HSP70等蛋白的含量[2]。除此之外,精氨酸在体内合成蛋白质、NO以及促进细胞分裂、伤口复原、免疫功能和分泌激素各方面有着重要作用,是机体不可或缺的。哺乳动物机体中几乎所有组织均可利用L-精氨酸合成胞浆蛋白和核蛋白[3]。近年来,发现精氨酸可以通过NO途径参与机体的营养调控与免疫调节,故人们加大了对精氨酸的研究。
1 精氨酸的理化性质
精氨酸是一种双性氨基酸,包含L-精氨酸和D-精氨酸两种形态。生物体内常见的为L-精氨酸,是幼年动物的必需氨基酸之一,成年动物的条件性必需氨基酸。在动物体内起营养、代谢及免疫功能的是L-精氨酸。分子式为C6H14N4O2,比旋光度+26.9 °~27.9 °,相对分子质量为170.20,是白色斜片状(二水物)晶体或白色结晶性粉末。熔点244 ℃,经水重解结晶后,于105 ℃失去结晶水。其水解呈强碱性,可从空气中吸收二氧化碳。溶于水(15%,21 ℃),不溶于乙醚,微溶于乙醇。天然精氨酸由于胍基的存在,呈碱性,易与酸反应形成盐,由瓜氨酸通过精氨酸琥珀酸合成酶(ASS)及精氨酸琥珀酸裂解酶(ASL)合成,这个过程需较大的能量,因为要将每一个分子合成精氨基需要将三磷酸腺苷(ATP)水解成一磷酸腺苷(AMP),即两个三磷酸腺苷需求量。精氨酸不仅是蛋白质的基础构件,还是许多低分子量生物活性分子综合体合成的前体,包括NO、脯氨酸、谷氨酸、多胺、肌酸等。多胺对细胞的生长、分化有关键的调节作用,其他物质在生物体内都有不同的生理和免疫功能。
2 精氨酸的合成代谢
2.1 精氨酸的合成
在过去30年的研究中,已经确定哺乳动物小肠中释放的瓜氨酸几乎是内源性氨基酸的唯一来源。肾脏是成年动物肠源性瓜氨酸通过精氨琥珀酸合成酶和裂解酶转化为精氨酸的主要场所,且该合成途径几乎发生在所有类型细胞的生命周期中[4]。新生仔猪自身合成的精氨酸无法满足机体需要,通常需要从外界获取。动物机体内的精氨酸来源主要有3个:一是食物蛋白;二是内源合成;三是机体蛋白质代谢,其中最主要的来源是食物[5]。Blachier等[6]研究发现,幼年动物体内精氨酸合成大部分是发生在小肠上皮细胞。小肠上皮细胞会从谷氨酰胺和谷氨酸产生瓜氨酸,再经由肾脏的肾小管细胞协助抽取出来并转化为精氨酸。WU等[4]同样发现,猪小肠中的谷氨酰胺(谷氨酸)和脯氨酸(猪乳中的丰富氨基酸)是瓜氨酸和精氨酸合成的前体。P5C是谷氨酰胺(谷氨酸)和脯氨酸在精氨酸合成途径的中间体(图1)。因此,仔猪只有从外界获取一定量的精氨酸才能维持机体的正常发育和促进发育。
2.2 精氨酸的代谢
研究表明,畜禽体内的精氨酸主要是由小肠中段吸收,即日粮中的氨基酸主要由小肠吸收,其中精氨酸有约40%是在小肠内被消化,其他部分进入体内循环,转化成其他功能物质。
精氨酸的代谢途径多种多样,主要有以下几种方式,如图2[7]所示,第1种是在精氨酸酶Ⅰ的作用下分解为尿素和鸟氨酸,鸟氨酸经过一系列反应转化为多胺、谷氨酰胺等,谷氨酰胺再进入Krebs循环,生成CO2;第2种是氧化途径,一氧化氮合成酶在还原型辅助酶Ⅱ、Ca2+、黄素腺嘌呤二核苷酸等作用下将精氨酸分解为瓜氨酸和NO。NO可以杀死体内的寄生虫、细菌,抑制癌细胞增殖以及促进血管舒张,而且NO对糖尿病、肥胖症中葡萄糖、脂肪有一定的影响作用,故我们可以加强对NO通路的研究,了解其作用机制。第3种是在精氨酸酶的作用下分解为尿素和鸟甘酸。
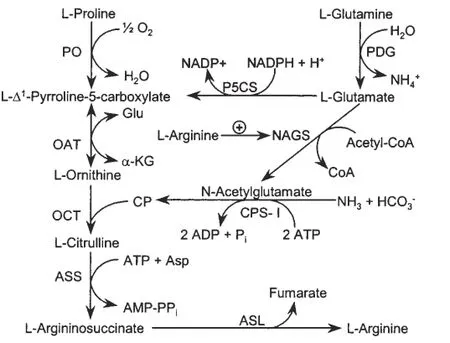
图1 谷氨酸和脯氨酸合成精氨酸途径[4]

图2 精氨酸的主要分解代谢途径[7]
3 精氨酸对仔猪的营养调控作用
3.1 仔猪的生理特点
有多数研究表明,仔猪的死亡率与初生重关系紧密[8-9]。Smith等[10]研究猪早期断奶应激表明,早期断奶(20日龄)导致长期的有害变化,包括影响胃肠道屏障性能,炎症出现率增加等。新生仔猪离开母体后,消化道发育不全,不耐低温,免疫功能低下,而早期断奶仔猪由于应激会引起心理和营养系统的急剧变化,应激包括和母猪的分离,环境温度的降低,与新的陌生仔猪的竞争以及新的位序设置等[11]。正常情况,仔猪的消化系统14日龄左右发育成熟,约40日龄发育完善。断奶1周后胰酶等多种消化酶活性下降,直至断奶2周后恢复[12-13]。消化系统的不健全,导致仔猪对外界环境应激和饲料消化难度增大,影响仔猪的生长发育。新生仔猪自身的免疫系统也尚未构建完全,主要是依靠从母乳中获取的免疫球蛋白抵抗免疫应激,而母乳中的免疫球蛋白含量会越来越低,故对疾病的抵抗能力有限[14]。仔猪在断奶后的20 d左右开始建立自己的免疫系统,但速度缓慢,此阶段仔猪抵抗力弱,患病几率高。同时,在断奶后的两周时间,仔猪会大量死亡,高的甚至达到35%以上。此外,断奶后,仔猪的小肠形态结构发生变化,Cera[15]研究发现,断奶仔猪小肠占空体重的比例高于同龄仔猪,说明仔猪断奶应激时部分营养物质用于发育小肠组织。刘玉兰等[16]研究了在日粮中添加脂多糖(LPS),再饲喂精氨酸能在一定程度上缓解LPS刺激所导致的生长抑制和应激反应,即对仔猪补充精氨酸可以降低其断奶应激。仔猪的小肠黏膜是精氨酸合成的重要场所,而幼畜体内的精氨酸酶活性较高,能快速分解精氨酸,降低了精氨酸的净合成量。早期仔猪断奶的目的是尽可能减少外界刺激和感染,提高经济效益,所以,应当做好断奶仔猪的饲养管理和营养调控,提高仔猪存活率等各项指标。
3.2 精氨酸对仔猪生长性能的影响
精氨酸是动物体内含氮最多的氨基酸。精氨酸分解后产生的氮可维持体内的氮平衡,是组织中蛋白质最丰富的氮载体,其有多种使用途径,在细胞中呈现出显著的代谢和调节功能。日粮中添加适量的精氨酸能有效提高仔猪的平均日采食量和日增重并提高多种蛋白和激素的表达水平,提高饲料利用率,增加经济效益。H.SHENZHOU[17]的试验结果显示,在断奶仔猪日粮中添加精氨酸,仔猪的日增重显著提高(P<0.05),料重比显著降低(P<0.05)。He等[18]在仔猪日粮中分别添加精氨酸和丙氨酸发现,精氨酸增重效果更明显,比丙氨酸组高出5.6%。精氨酸组的料重比也低于丙氨酸组和对照组。Yang等[19]研究表明,仔猪断奶前补充精氨酸在其21日龄时胴体重显著增加(P<0.05),且在11~21日龄阶段平均日增重显著提高(P<0.05),均以8 g/kg的精氨酸添加量增加最明显。Hu等[20]发现,在猪基础日粮中添加1%、2%精氨酸显著提高猪的体重(P<0.05)和生长性能,添加2%的精氨酸效果显著(P<0.05)高于1%组。因而,在日粮中添加适量的精氨酸能有效提高猪的各项指标,增加效益。
4 精氨酸对断奶仔猪肠道的影响及机制
4.1 对黏膜生长和免疫功能的影响
肠道健康对断奶后哺乳动物的机体健康和生长发育至关重要[21]。免疫系统是由免疫细胞和免疫器官共同组成。早期断奶仔猪由于机体抗逆性较弱,以及来自环境等的应激,易发生断奶应激综合征[22]。而肠道是机体内最大的免疫器官,也是食物消化吸收的主要场所。肠道黏膜结构和功能的正常决定仔猪肠道的健康。仔猪的断奶应激会刺激肠道黏膜和功能,影响正常功能。断奶仔猪小肠可见明显的绒毛萎缩和隐窝增生、伴随肠道生化改变的组织学变化、肠道功能下降、腹泻。小肠绒毛的高度和隐窝深度决定着肠道的健康,在一定范围内,绒毛高度与小肠吸收能力呈正相关,隐窝深度与肠道健康呈负相关。研究表明,断奶仔猪基础日粮中添加适量精氨酸可增加小肠绒毛高度并降低隐窝深度。Huang等[23]声称,对于4~21日龄的仔猪,用牛奶替代日粮可促进肠道的发育,尤其是小肠的相对重量,以及增加小肠绒毛的高度、面积和功能的强化。
Yang等[19]研究表明,仔猪断奶前饲喂精氨酸增加了十二指肠、空肠、回肠绒毛高度、表面积,以及降低它们的隐窝深度。在一定范围内,它们与精氨酸含量呈正相关,在精氨酸含量为8 g/kg时效果最显著(P<0.05)。Yao等[24]在断奶仔猪基础日粮中添加精氨酸,提高了黏膜和黏膜下层血管内皮生长因子的表达;Yao等还发现在21日龄的阉割断奶仔猪的日粮中添加1%的精氨酸,小肠的相对重量高于对照组33%,隐窝深度显著低于对照组(P<0.05),小肠绒毛高度也显著提高(P<0.05)。Puiman等[25]报道,精氨酸可以促进仔猪肠道黏膜的生长。寇涛等[26]研究表明日粮中添加精氨酸能显著提高仔猪小肠黏膜上皮细胞和杯状细胞数量,提高仔猪免疫功能。Tan等[27]饲喂断奶仔猪不同含量的精氨酸,结果显示0.6%和0.8%组的体重以及平均日增重显著高于其他组(P<0.05)。此外,还发现不同的精氨酸添加量分别提高了小肠IgM、IgG、IL-2等基因表达水平。WU等[28]研究发现,在21日龄的断奶仔猪日粮中添加6 g/kg的精氨酸1周,提高了小肠绒毛高度和隐窝深度,同时改善了肠道黏膜萎缩。
4.2 对肠道免疫功能影响机制
猪的肠道与人的肠道结构具有高度同源性,故对其肠上皮细胞的研究可为人体肠道黏膜的研究提供一定的参考。精氨酸通过对蛋白质翻译水平的影响来调节仔猪肠道上皮的修复,还能调节肠道内细菌新陈代谢和上皮细胞多种受体的表达。最近几年有研究表明,用精氨酸处理肠上皮细胞能通过雷帕霉素/核糖体蛋白S6(p70S6K)通路可提高细胞内蛋白质的合成量[29]。Ren等[30]在小鼠体内试验得出添加精氨酸显著降低(P<0.05)了空肠中TLR6的mRNA水平,增加了TLR8的mRNA水平。同样,回肠中的TLR4、TLR5、 TLR8的mRNA水平均显著增加。Gao等[31]在肉鸡的胚胎中注射精氨酸能提高肠黏膜中IL-2、IL-4含量以及TLR2、TLR4的mRNA表达,还发现能增加iNOS蛋白的丰度。iNOS对L-Arg有明显的催化作用,使其生成NO。NO能够通过抑制DNA和蛋白质合成以及使酶失活等方式起消毒、灭菌作用。Xia等[32]研究发现,在肠上皮细胞培养基中添加0.02 mmol/L精氨酸,CAT1、EAAT3和ASCT1的mRNA的相对表达水平显著高于0 mmol/L组与0.07 mmol/L组。CAT1是y+系统中重要的转运蛋白,能够转运L型lys、Arg和Orn三种氨基酸,参与蛋白质合成,EAAT3和ASCT1在细胞中都分别发挥重要作用。H.SHENZHOU[17]观察到断奶仔猪日粮中添加精氨酸促进了胰岛素生长因子(IGF)的表达。IGF信号通路是调节胎儿和胎盘的生长、增殖和抑制哺乳动物细胞凋亡的重要调节因子。朱惠玲等[33]声称添加精氨酸能减少肥大细胞数量,增加sIgA、CD4+、CD8+细胞数,说明日粮中添加精氨酸能有效改善肠道黏膜屏障功能。Corl等[34]观察到添加精氨酸增加了肠黏膜蛋白的合成,改善了肠黏膜上皮的渗透性。
5 小结与展望
近些年,随着对精氨酸生理作用研究的加深,发现其功能已经超出一般氨基酸的功能范围。精氨酸作为动物机体的条件性必需氨基酸,在蛋白质合成以及动物肠道免疫等多个过程都有关键作用。有关研究中还发现,精氨酸提高免疫的作用可能替代抗生素等,改变当下抗生素禁用后的境况。对于不同动物的不同生理状态来说,精氨酸的最佳摄取量有待考察。精氨酸对畜禽和人体的营养及保护作用需进一步地研究。
略,若需要请与作者联系)
2017-05-24)
苏北科技专项
毛俊舟(1993-),男,安徽六安人,硕士研究生,从事单胃动物营养研究,E-mail:jzm781446817@163. com
猜你喜欢
小学阅读指南·教研版(2023年7期)2023-07-17 09:26:41
今日农业(2022年2期)2022-11-16 12:29:47
基层中医药(2021年6期)2021-11-02 05:46:16
现代畜牧科技(2021年2期)2021-03-19 07:48:26
河南畜牧兽医(2020年11期)2020-01-11 05:31:49
故事会(2019年10期)2019-05-27 06:06:58
广东饲料(2016年4期)2016-12-01 03:43:18
中外医疗(2015年11期)2016-01-04 03:58:50
中国当代医药(2015年30期)2015-03-01 02:08:19
中国当代医药(2015年30期)2015-03-01 02:08:13
