
洱海蓝藻爆发的时空特征及影响因子
2017-08-28 15:14:37王教元陈光杰黄林培刘晓东陈倩倩刘晓海张虎才
生态学报 2017年14期
王教元,陈光杰,*,黄林培,刘晓东,陈倩倩,刘晓海,张虎才
1 云南师范大学旅游与地理科学学院高原地理过程与环境变化云南省重点实验室,昆明 6505002 中国科学技术大学地球与空间学院极地环境研究室,合肥 2300263 云南环境科学研究院,昆明 650034
洱海蓝藻爆发的时空特征及影响因子
王教元1,陈光杰1,*,黄林培1,刘晓东2,陈倩倩2,刘晓海3,张虎才1
1 云南师范大学旅游与地理科学学院高原地理过程与环境变化云南省重点实验室,昆明 6505002 中国科学技术大学地球与空间学院极地环境研究室,合肥 2300263 云南环境科学研究院,昆明 650034
湖泊生态环境对区域气候变化和流域人类活动十分敏感。随着流域的持续开发,洱海作为云贵高原第二大淡水湖泊面临着严重的生态与环境问题,主要包括水质恶化和生态功能的衰退,其中蓝藻水华问题尤为突出。针对湖泊现代监测数据存在时间序列较短、连续监测记录缺乏、监测位点不完全一致等问题,应用沉积物记录开展色素等多指标分析和环境变化重建研究,并对洱海湖区南、中、北3个湖盆的沉积物记录进行对比分析,从而探讨洱海富营养化与蓝藻爆发的历史与变化特征,并识别藻类响应模式的空间异同。沉积物色素记录结果表明,洱海蓝藻生物量变化具有明显的时空差异性,呈现由南至北,先后增加、最后呈现蓝藻水华全湖性持续爆发的模式。进一步的简约模型方差分解结果表明气候变暖和营养盐富集是洱海蓝藻生物量变化的主要驱动因子,此外相对较浅的南部湖盆还受到水位波动、水动力减弱、水生植物演化的综合影响。因此,在气候变暖的背景下,控制水体营养盐输入、合理调控湖泊水位、提高水体透明度并恢复水生植物是控制洱海蓝藻水华爆发和进行生态恢复的重要措施。
沉积物色素;蓝藻爆发;时空差异;洱海
湖泊生态系统的结构与健康是湖泊水质与流域环境特征的敏感指示器,能够综合反映气候变化和人类活动的长期影响[1]。蓝藻水华的发生反映了浮游植物群落结构的转变,同时也指示了湖泊生态系统健康的恶化,如部分水华蓝藻能产生毒素,通过饮用水和渔业资源危害人类健康[2],且持续的、高生物量的蓝藻爆发会对整个湖泊环境与生态系统产生长期的影响。蓝藻水华爆发的主要驱动因素包括适宜的光照、温度、营养盐等条件[3]。入湖氮、磷等营养盐的富集可促进初级生产者(如浮游植物)的大规模繁殖[4],反映了可获得性资源的上行效应对藻类群落结构与生产力的控制作用[5]。同时水文水动学过程也被认为是影响浮游植物的重要因子[6],如水动力的减弱可导致蓝藻在水体表面聚集[7]。水动力学特征可通过改变水体的混合强度和热力分层直接影响浮游植物在水柱中的垂直分布,也可以调节影响藻类生长过程所经历的水温和光强。水动力过程的变化也影响水体中颗粒物与营养盐的再悬浮与传输过程,从而间接驱动浮游植物群落生产力与结构的变化[8]。
随着洱海流域开发规模的增强和水体水质的日益恶化,1996年首次报道洱海出现全湖性藻类水华后,近年来全湖性水华事件不断,已经对洱海生态系统造成了严重影响。现代调查结果显示, 黎尚豪等发现1957年洱海南部湖区出现水华束丝藻[9],1980s—1990s初期以水华束丝藻为主的水华主要出现在洱海南部[10],1996年秋季爆发的水华优势藻属则转变为螺旋鱼腥藻,1998年为卷曲鱼腥藻水华,1999年以后铜绿微囊藻成为洱海水华的优势属种。从藻类组成来看,洱海蓝藻优势种类出现长期演替的特征,此外还夹杂有硅藻水华和绿藻水华,但总体出现蓝藻生物量不断增加,且占藻类总物量的比例不断提高,最终处于绝对优势[11-12]。
目前,对于洱海藻类水华的研究主要是报道水华发生年份的优势种群、生物量和分布状况,以及水质营养水平、浮游植物多样性的季节变化,但对不同湖区藻类爆发的长期历史缺少系统研究。同时,由于洱海环境压力类型较多[13-14]、湖盆结构复杂[15]等特征,对洱海藻类水华的爆发历史与驱动因子识别仍需考虑湖盆间水文、水质等空间差异。沉积物色素是指示湖泊藻类生产量与浮游植物群落变化的可靠指标[16-17],在定量重建湖泊初级生产力变化研究中被广泛应用[18- 20]。玉米黄素(Zeaxanthin)和海胆酮(Echinenone)这两个组分作为指示蓝藻的特征色素,由于其在沉积物记录中化学性质相对稳定[21],是表征历史时期水体中蓝藻生物量的主要指标,在海洋和湖泊研究中被用于评估蓝藻生物量的长期变化及识别富营养化和蓝藻爆发的历史[22- 24]。因此,本文针对洱海水质与藻类调查缺乏长期与连续的监测数据,利用洱海不同湖区的沉积物柱芯样品,通过沉积物测年、沉积物蓝藻特征色素、营养元素多指标分析方法,重建了近百年来洱海3个湖区水体富营养化与蓝藻爆发的历史,识别了蓝藻爆发的时空差异特征与主要环境因子的驱动强度,可为洱海的生态修复与综合治理提供重要的基础数据与科学依据。
1 研究区域与研究方法
1.1 研究区概况
洱海(25°36′—25°58′N,100°06′—100°28′E)位于云南省西北部,大理白族自治州中部,(图1),属澜沧江流域黑惠江支流的构造断陷湖泊,流域面积为2565 km2。湖面海拔高度为1965.8 m(八五高程),湖泊面积251.35 km2,最大水深19.5 m,是云贵高原第二大淡水湖。水体矿化度为167—175 mg/L,平均为172.33 mg/L,pH值在8.3—9.0之间变化(平均值=8.5)。湖水主要由地表径流和降水补给,主要入湖河流有22余条:如东侧的凤尾箐、玉龙河,西侧的苍山十八溪水,南侧的波罗江,北侧的罗时江、弥苴河、永安江等,西南角的西洱河是洱海唯一的天然出湖河流,最终汇入澜沧江。湖区属中亚热带高原季风气候,年均温15℃,年均降水量1056.6 mm(图2),径流的年内变化主要受气候因素的影响,雨季5—10月径流量占年径流量的65%以上[25]。
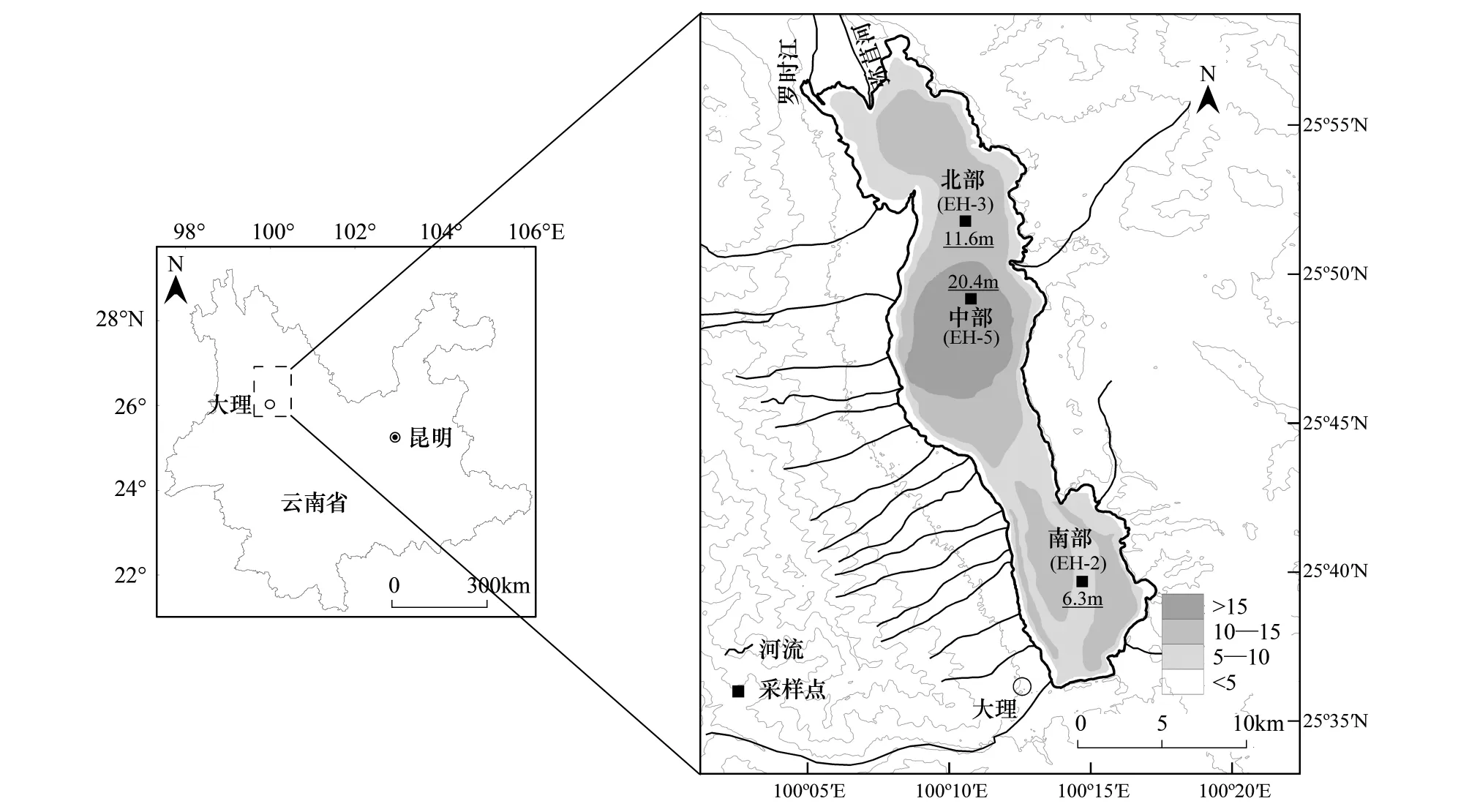
图1 洱海及采样点位置及水深Fig.1 Site map showing the location of Erhai Lake, the coring sites and related water depthsEH-3、EH-5、EH-2分别为洱海北部、中部、南部湖盆采集的沉积物柱芯编号
湖泊现代监测结果表明,1970s以来洱海水位变化剧烈,且水体富营养化进程加快,水质呈现不断下降的趋势[26-27](图2)。1974年以前洱海水位年际变化较小,年均水位维持在1974 m左右(八五高程,下同),1974—1982年洱海年均水位急剧下降,低至1971m左右,比多年年均水位低3 m左右,是水位最低时期。之后,洱海水位缓慢、波动回升,并保持在相对低水位(年均水位1972 m左右)运行。洱海水生植物的覆盖度自1970s末期以来持续下降,由1977年40%左右迅速下降到2011年5%左右,说明洱海水生植物面积近40年来骤减,生态功能不断退化。洱海水体主要营养盐浓度自1970s呈现快速增加的趋势,水体总氮(TN)自1985年的200 μg/L增加至2006年的700 μg/L;水体总磷(TP)自1992年的10 μg/L增加至2006年35 μg/L,表明洱海水体营养水平已经由贫-中营养状态转变成中-富营养状态。藻类密度由1998年之前的低密度状态(2×106个/L)转为2000年代的高密度状态(15×106个/L)。水体透明度(SD)由2003年之前的>3 m下降至近年的<1.5 m,表明洱海水体已经从高透明度、清水状态转变为低透明度、浑浊状态(图2)。
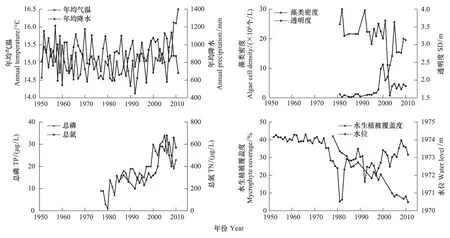
图2 洱海流域气象、水文、水质数据(数据来源:文献[26-33]) Fig.2 Documentary data showing the time series of meteorological, hydrological and water quality changes[26-33]
1.2 样品采集与分析
2012年3月对洱海不同湖区进行了系统的湖泊调查和样品采集,分别在洱海南部(水深6.3 m)、北部(水深11.6 m)和中部湖区(水深20.4 m,图1)用重力采样器采得保持完好的沉积岩芯6根(其中每个采样点提取平行钻2根),沉积物-水体界面的悬浮层均未受扰动,界面水清澈透明。沉积物岩芯按0.5 cm间隔在野外进行避光分样,样品均封装于聚乙烯自封口塑料袋并避光、冷藏运回实验室后,立即置于-20℃冷冻保存用于室内实验分析。本文利用沉积物柱芯EH- 2(南部湖盆)、EH- 3(北部湖盆)、EH- 5(中部湖盆)进行年代学、色素和其他理化指标测定。
本研究通过对样品放射性核素210Pb与137Cs的测试分析用于建立年代学序列。实验步骤包括:称取3—5 g干重样品放入离心管中用玻璃棒压实至4 cm左右高度,向其中注入约1 cm的环氧树脂AB结构胶至少静置2周,并量取样品高度。利用美国Canberra公司生产的高纯锗探测器、数字化谱仪及多通道分析系统,并应用恒定补给速率CRS模型建立年代学序列。
沉积物碳氮同位素样品的处理步骤包括:研磨冻干沉积物样品,定量称取10 mg左右并用锡箔纸包裹,将处理好的样品送入反应炉中,产生的气体通过FLASH 2000装置与Thermo MAT- 253质谱仪与元素分析仪进行测定。碳、氮同位素组成和元素含量分别对应国际标准PDB与大气中的N2,同位素和元素含量的分析误差均分别为±0.1‰和±1.0‰。
沉积物色素的分析参考Leavitt和Hodgson的国际通用方法[4]。主要步骤包括:沉积物冻干样品在-4℃下通过丙酮-乙醇-水混合溶液萃取后,经0.22 μm GF/F膜过滤后的溶液在暗光条件下用氮气吹干富集,所得样品在Agilent 1200系列HPLC系统测定,结果经丹麦DHI公司提供的标样校正。上述色素分析工作由加拿大麦吉尔大学生物系和中国科技大学极地研究室完成。
1.3 数据分析
沉积物玉米黄素和海胆酮浓度是反映湖泊蓝藻生物量变化的主要指标,首先应用主成分分析(Principal Component Analysis, PCA)提取蓝藻色素变化的主要方向。之后,我们对环境数据进行标准化处理,应用冗余分析(Redundancy Analysis, RDA)探究环境因子对蓝藻色素浓度变化的解释能力,保留独立冗余分析中P<0.05的环境变量进行方差分解分析,检查方差膨胀系数并剔除膨胀系数>20的环境变量,对环境变量进一步进行简约模型选择得到驱动蓝藻色素变化的显著环境变量。由于气象、水文等现代监测数据主要开始于1951年,且3个沉积物钻孔色素变化主要也发生在1950年代以后,因此RDA分析主要覆盖1951—2011年的时间段。本文所有统计分析均在R软件(版本2.14.0)中完成。
2 结果与分析
2.1 沉积物年代序列
基于沉积物210Pbex、137Cs活度和CRS模型建立的钻孔年代-深度模型显示沉积物钻的210Pbex比活度随深度的增加呈指数递减(图3),137Cs曲线自底部出现的第一峰值,其CRS模型年代(±1SE)分别为(1958±3)年(EH- 2)、(1963±1)年(EH- 3)、(1960±2)年(EH- 5),可较好地与1963年的137Cs蓄积峰相对应。3个沉积物钻孔出现的第二个峰值大约在1986年,可以与北半球记录的1986年切尔诺贝利核电站核泄漏事件产生的峰值相对应。湖盆北部、中部沉积物岩芯自底部,沉积速率呈波动上升的趋势,其中分别从8 cm(~1993 AD)、4 cm(~2000 AD)至表层出现明显的持续升高。湖盆南部沉积物岩芯的沉积速率波动较大,自底部至10 cm(~1900—1980 AD)呈现升高趋势,在5—10cm(~1988—1992AD)深度出现明显的峰值,随后自5 cm深度附近至表层沉积速率有所降低(图3)。
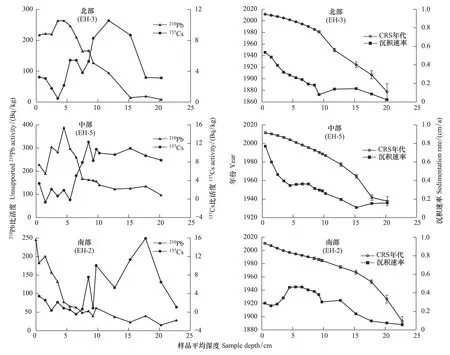
图3 沉积物3个钻孔210Pbex、137Cs活度变化和测年结果Fig.3 Depth profiles showing 210Pbex and 137Cs radioactivities, CRS models and sedimentation rates for three cores, respectively
2.2 洱海蓝藻爆发历史的色素记录
洱海北部钻孔(EH- 3)的两种蓝藻色素记录呈现了同步的变化趋势,1990年之前含量均偏低,其中玉米黄素含量均值(±1SD)为(0.66±0.26)μg/g、海胆酮含量为(0.73±0.24) μg/g。从1990年开始出现显著的上升,大约2003年蓝藻色素浓度出现一个峰值,玉米黄素达到(6.27±1.27)μg/g、海胆酮达到(4.62±0.86)μg/g,而2008—2011年出现连续的、大幅的上升,玉米黄素含量为(8.68±3.44)μg/g,海胆酮含量为(5.63±1.75)μg/g。总体而言,湖盆北部沉积物蓝藻色素自1990年出现持续增加,并在~2000年以后总体维持在较高的含量水平(图4)。
中部钻孔(EH- 5)沉积物蓝藻指示色素浓度变化趋势可以分成两个主要阶段:2000年之前变化波动较小且处于较低水平,如玉米黄素、海胆酮含量均值(±1SD)分别为(0.98±0.39)μg/g、(1±0.34)μg/g。大约在1996年,出现一个弱峰值;2000年之后,两种色素均出现显著的增加且处于较高水平,且在2009年左右达到峰值,如玉米黄素达到(9.82±6.87)μg/g、海胆酮达到(5.23±3.17)μg/g(图4)。
南部钻孔(EH- 2)沉积物蓝藻色素含量1900—1985年间年以来呈现持续增加的趋势但波动幅度不大,玉米黄素含量均值(±1SD)为(3.56±1.10)μg/g、海胆酮含量为(3.14±0.65)μg/g。~1985年之后增加趋势明显且波动较大,玉米黄素含量为(6.96±2.25)μg/g、海胆酮含量为(4.17±1.24)μg/g,且在1983、1986年左右分别出现两个弱峰值。大约1995—2006年间,南部湖盆沉积物蓝藻色素出现一个小幅的下降,2006年来又出现明显的上升(图4)。
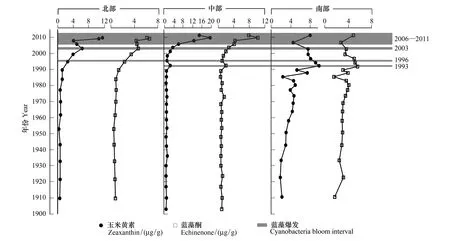
图4 沉积物钻孔蓝藻色素记录地层图Fig.4 Sedimentary profiles of cyanobacterial pigments for the three cores of Erhai Lake
2.3 洱海沉积物营养元素的变化特征
洱海北部和中部沉积物钻孔总氮(TN)含量变化与沉积物叶绿素含量变化趋势基本一致(图5)。北部钻孔TN含量自1960年持续增加,由2.0 mg/g左右增加至7.0 mg/g左右,与蓝藻色素变化主轴1得分显著相关(r=0.931,P<0.001)。中部钻孔大约1995年之前TN含量偏低且比较稳定,1995—2005年间出现明显的升高,达到10.0 mg/g左右的峰值,2005年之后又有一个明显的下降趋势,2008—2011年维持在5.0 mg/g左右含量水平,与蓝藻色素变化主轴1得分显著相关(r=0.507,P<0.01)。洱海南部钻孔TN含量1960年之前含量较低且稳定,1960—1985年缓慢增加到5mg/g左右,1995—2011年波动上升至7.0 mg/g左右,与蓝藻色素变化主轴1得分显著相关(r=0.656,P<0.001)。
洱海北部钻孔的总碳(TC)含量在1960年之前无明显变化且含量较低(平均值为19.6 mg/g),1960年之后显著持续地增加,TC含量为前一个时期的3倍左右(平均值为64.3 mg/g)。中部钻孔TC含量的显著变化出现在1990s后期,1990年之前含量维持在25.0 mg/g左右,之后的大约10年时间内迅速升高至90.0 mg/g左右,自2003年又出现明显的下降趋势,但TC含量仍较高。南部钻孔TC含量从大约1960年开始出现持续的增加,在1980s后维持在较高的含量(平均值为108.8 mg/g),大约1990年之后出现小幅的、缓慢的下降(图5)。
洱海北部 、中部钻孔沉积物的全岩C/N比值范围总体为8—15且变化较小,南部钻孔的C/N比值在10—35之间变化,同时波动较大。北部钻孔C/N比值1910—1970年间由14左右缓慢持续地下降到11左右,1970年左右出现明显的升高,并在1990年之后保持在14左右。中部钻孔C/N比值在2000年之前基本保持在9左右,大约1998年左右升高至14左右,2000年后在12左右。不同于北部和中部钻孔,南部钻孔的C/N比值在1900—1970年间基本为10左右,在1970—1980年、1985—1995年两个时段出现明显的升高,达到30左右的峰值,1995—2011年则又呈现持续的降低,并维持在18左右(图5)。
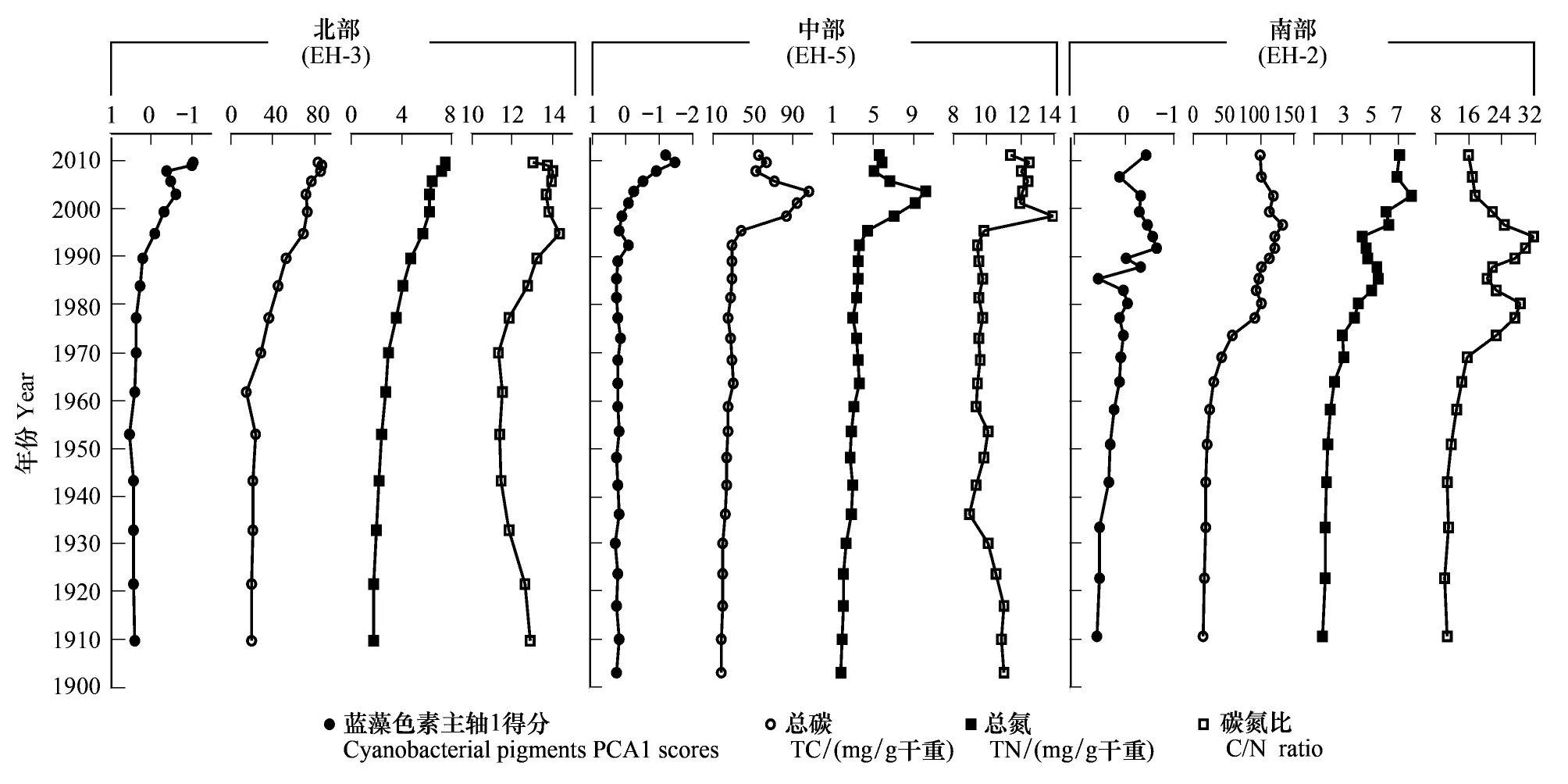
图5 洱海沉积物蓝藻色素PCA主轴1得分与碳氮元素结果Fig.5 Comparison of sedimentary cyanobacterial pigments PCA1 scores and C N elements
2.4 洱海蓝藻色素含量变化的驱动因子
为了识别蓝藻色素变化的主要影响因素及其驱动强度,我们结合现代气象、水质、水文、营养盐等监测数据与沉积物理化指标记录,对沉积物钻孔蓝藻色素数据进行独立的RDA分析。进一步的简约模型方差分析结果表明(图6),驱动湖盆北部、中部蓝藻色素含量变化的显著环境因子均为气温和TN。其中,气温、TN分别独立解释了北部湖盆蓝藻色素变化的55.7%、13.7%,两者共同解释了8.4%。与之相似的,气温、TN分别独立解释了中部湖盆蓝藻色素变化的38.3%、15.3%,两者共同解释了12.2%。与湖盆北部、中部钻孔记录明显不同的是,沉积物C/N比值、气温和TN则是驱动南部湖盆蓝藻色素含量变化的3个主要环境因子,其中C/N比值独立解释了南部湖盆蓝藻色素变化的15.2%,气温和TN分别解释了10.7%、9.2%,其中气温和TN共同解释了13.0%,但是未解释的比例(50.0%)高于中部和北部湖区(分别为34.2%和22.2%),指示了驱动湖盆南部蓝藻生物量变化的过程较湖泊中、北部地区更为复杂。
由于南部湖盆较浅且受水位大幅变化影响较大[34],为评价水位变化对沉积物色素降解和保存的影响,将 1950年之后的水位监测数据(图2)和湖盆南部钻孔的沉积物蓝藻色素进行了相关性分析,结果显示两者之间无显著的相关关系(P> 0.5),表明了水体水深与水位变化对沉积物蓝藻色素的影响是十分有限的。

图6 驱动蓝藻色素变化的简约模型选择与方差分解结果Fig.6 Summary of variance partitioning results for key environmental variables included in the minimum model at each site
3 讨论
本研究初步揭示了洱海的蓝藻爆发历史与现代监测记录基本一致,而蓝藻生物量呈现由南至北先后增加的空间特征,最后表现为蓝藻水华的全湖持续爆发。方差分解结果进一步表明,气温上升和营养盐富集是导致洱海不同湖区出现蓝藻增加与爆发的重要驱动因子,而与C/N比值变化相关的水生植物演替、水动力变化等环境过程与南部浅水湖盆的蓝藻变化模式最为密切相关。研究结果表明,洱海湖盆地形与流域开发类型的空间异质性可能导致了该大型湖泊蓝藻生物量变化呈现明显的时空差异模式。
3.1 湖泊富营养化与蓝藻生物量变化
已有研究表明营养盐的增加(即水体富营养化)是蓝藻水华爆发的主要驱动因子之一[35],而N、P是控制浮游植物(特别是蓝藻)生长的关键生源要素[36]。洱海湖泊水质调查结果表明,水体N、P输入自1980年以来总体持续增加且与浮游植物数量(如藻类密度)的增加显著相关[30],都表明了营养盐的富集对浮游植物数量的促进作用[37-38]。沉积物TN记录反映了湖泊营养盐持续增加:自1960s开始,洱海湖盆不同地区的沉积物TN含量开始增加并在1990s达到较高水平,与此同时,沉积物蓝藻色素含量呈现同步增加的变化趋势(图5)。沉积物蓝藻色素记录与1993年、1996年、2003年以及2000s中后期洱海全湖性蓝藻水华事大致是吻合的[24- 28],表明洱海蓝藻水华主要发生在1990s之后。南部钻孔1980s即出现蓝藻色素的增加,表明洱海南部蓝藻生物量增加最早,可能最先出现蓝藻水华现象,这与相关的湖泊调查的结果是一致的[33]。
洱海湖盆不同位置的沉积物蓝藻色素记录表明蓝藻爆发存在明显的时空差异特征。如南部湖盆沉积物蓝藻色素最先出现增加的趋势,而北部、中部湖盆蓝藻爆发主要发生在1990s以来。蓝藻水华发生的空间差异不仅与湖盆自然条件有关,也受流域开发类型与空间分布的影响。洱海北部的洱源县是云南省重要的畜牧业和商品粮基地之一,农业开发历史较长,如该地区自1960s开始施用工业化肥,年均使用量超过4000t[39]。北部钻孔沉积物TN浓度自1960年开始持续增加,这与农业施肥和牲畜畜养量的增加趋势是一致的。相关调查结果显示,洱海流域北部洱源县的氮、磷产生量最高,洱海流域西部和南部乡镇农业氮磷产生量次于北部,洱海东部乡镇农业氮磷产生量相对最低[40]。而洱海南部、西南部是洱海流域人口主要聚居地区,也是洱海流域范围内城镇、工业活动较为集中的地区。随着城镇化和工业化进程的增强,工业生产污水和城镇生活污水排放量自1970s开始迅速增加[41],大量污水注入导致洱海水质自1990s以来迅速恶化。
洱海沉积物营养元素含量呈现出与色素记录类似的空间变化特征。湖盆北部钻孔位置可能受到主要入湖河流弥苴河和罗莳江河口的影响,同时反映了洱海北部流域以农业开发为主的人类活动影响。自1960s始,农业活动的加强(包括耕地面积增加、农田水利工程的建设、森林植被破坏等)导致了外源有机质随入湖径流输入量的不断增加,从而出现了北部钻孔沉积物TC含量自1960年来持续升高的趋势(图5)。中部钻孔沉积物TC含量大约1990年之前含量较低且变化较小,大约1995年之后出现明显的上升,并在2000年左右达到峰值,随后有所下降。由于中部钻孔水深 >15m抑制了水生植被生长[42-43],因此沉积物TC可能主要反映了浮游藻类生产量的变化。湖盆南部钻孔沉积物记录的TC含量在1960年之前总体稳定但较北部、中部湖盆高,1960—1990年代中期以来含量持续升高,并在~1981年、1992年左右出现两个峰值,可能是由于水利工程修建导致了湖泊水位显著下降,水生植被得以持续生长,湖泊沉积物有机质有所升高。2000年以来,由于水生植被的消亡,浮游藻类(特别是蓝藻)占据主导优势,沉积物TC含量有所下降。
沉积物C/N比值常用于指示湖泊沉积物中有机质的主要来源[44-45],一般而言,湖泊内源藻类有机质 C/N比值较小(4—12),而陆源有机质C/N较大(>22)[46]。湖盆北部、中部的C/N比值均<15,说明中、湖盆北部的有机质主要来自内源性藻类;南部湖盆C/N比值在10—30间变化,说明湖盆南部的有机质主要受来自流域外源输入和内源水生植物的影响。此外,大型水草具有较高的C/N比值(>15),而低等植物(如藻类)的输入增加则可以导致沉积物C/N比值总体呈现降低的特征[47]。湖盆南部沉积物C/N比值在1900—1960年间一直处于持续缓慢上升的过程,但基本稳定在10左右,指示了该阶段湖盆南部有机质主要来自藻类等的大量增加。在1970—1978年、1988—1992年湖盆南部沉积物出现两个C/N值升高的阶段,这可能是水草大规模生长与外源有机质输入持续增加的结果,而1990年代中期之后浮游植物(特别是蓝藻)生物量的进一步增加则导致了C/N值出现持续的下降。
3.2 气候变化与藻类响应
气候变暖已成为了区域与全球气候变化的重要特征[48],而湖泊系统对气候变化的响应十分敏感[49-50]。气象监测资料表明,洱海流域1980年以来呈现明显的气温升高、降水总体较低的趋势(图2),而太阳辐射量自1990年以来呈现波动上升的趋势[51]。洱海大规模蓝藻水华爆发的年份(1996年、2003年、2006—2012年)与云南地区出现的几次明显干旱事件在时间上相对应[52]。秦红杰的研究指出蓝藻具有抵御UV-B辐射的较强能力,因此小幅度的太阳辐射量增加对蓝藻水华的爆发有促进作用[53]。同时,气候变暖会增强深水湖泊热力分层的形成与维持,并降低水体垂直湍流的混合作用,都有利于蓝藻水华的发展、扩散和维持[54]。此外,高温可能会促进湖泊内源性营养的分解、释放,进而促进蓝藻生物量的不断升高与蓝藻水华的爆发[55-56]。
气候变暖不仅促进了蓝藻的生长,初级生产力的提高,同时也对整个浮游植物群落结构产生影响[57]。例如,淡水蓝藻DNA和蛋白质合成系统的热稳定性使得蓝藻可以适应高温条件[58],因此蓝藻属种具有明显的竞争优势。洱海浮游藻类优势种属经历了从1950年代的绿藻、甲藻、硅藻为主,到1980年代的绿藻、蓝藻、硅藻为主且甲藻门几乎消失,自1990年代以来蓝藻为绝对优势种类,绿藻、硅藻次之[9,59- 61]。洱海现代调查数据与沉积物蓝藻色素记录都表明了气候变暖对对蓝藻生长的促进作用,这与我国其他地区以及全球的湖泊研究结果是一致的[62- 65]。这种光合生物的优势类群转变在洱海形成了由蓝藻占优势的群落,可以明显改变湖泊营养盐的循环途径、食物网结构和生态系统功能[66]。
3.3 湖泊水文的影响评价
在营养水平较高的水体中,水动力条件可以成为影响蓝藻水华爆发的主要限制因子[67]。云贵高原地区的湖泊总体滞水时间较长,水体更新周期能力弱,入湖营养盐的持续输入会加速富营养化过程和藻类爆发[68]。1963年西洱海水闸建成后,洱海湖泊水位由自然调控变为人工调控;1973—1989年间,河床的加深和梯级电站的建设等导致了洱海水位显著降低。湖泊水动力的变化可以直接影响湖泊热力分层的稳定性、水体营养盐的浓度与滞留时间以及沉水植物的生长[6]。Schindler 和 Donahue指出湖泊的低水位可能会与高蒸发、高营养盐同时发生[38],从而导致水体营养水平的升高、浮游藻类的生长。在洱海全湖性的蓝藻水华发生时段内,洱海水位总体偏低,而水体TP、TN较高且藻类生物量迅速升高[31]。已有研究表明,湖泊水动力变弱会加剧水体营养化程度[69],从而促进藻类生长[70]。因此,由于修闸及梯级电站的建设等人类活动的增强,洱海水动力自1980s年代开始趋于变弱,从而有利于水体较浅且靠近西洱海水闸的湖泊南部地区蓝藻的生长与爆发。
同时湖泊的持续富营养化与水位波动可以抑制沉水植物的生长甚至导致其完全消亡[71],从而导致湖泊从清水状态向浊水稳态的转变[72]。洱海自1950年代至今经历了多次水位波动,水生植物覆盖面积比由1977年的~40%下降至2011年的<10%(图2),分布范围经历了从湖滨带连续分布、存在洱海湖盆南部到湖滨带不连续分布最后在洱海湖盆南部分布区的基本消失[73]。同时,沉水植物群落演替由海菜花、大茨藻、黑藻、苦草等为主且指示清水状态的、喜中贫营养的多优势种群落结构[74]发展到现今的以微齿眼子菜为主的单优势种群落结构[75]。洱海湖盆南部水深较浅(<10 m),是洱海水生植物分布的主要区域[76]。该湖区的沉积物TC和C/N比值呈现波动较大的特征,如在1980s初期、1990s初期出现两个峰值区,与西洱河梯级电站的建设、运行导致的湖泊低水位的时段一致,同期水生植物覆盖面积总体较高(如>20%),导致TC和C/N比值均较高。而自1996年以来,洱海总体处于相对高水位运行阶段且水体营养水平持续上升,湖盆南部水生植物出现覆盖面积的迅速缩减且同期藻类生物量明显升高(图2),从而导致了沉积物C/N比值的降低(图5)。
4 结论
本文通过沉积物色素和理化等多指标的综合分析表明,1960年左右开始洱海水体营养盐浓度升高并出现明显的富营养化过程,同时气候变暖趋势的加剧,叠加导致了湖泊初级生产力的升高和蓝藻水华的发生。同时洱海蓝藻爆发模式显著的时空差异性,湖泊水文调控加剧了相对较浅的南部湖盆蓝藻爆发的频次和规模。因此在气候暖的长期背景下,控制水体营养盐的输入、合理的水位调控并适当恢复水生植物是控制洱海蓝藻水华爆发并进行生态恢复的重要前提。同时,在湖盆形态与流域开发类型存在较大空间差异的大型湖泊中,需要对湖盆的典型区域开展生态环境调查并进行空间差异的对比分析,从而为污染湖泊的治理与生态修复提供针对性的保护与恢复措施。
致谢:云南师范大学高原地理过程与环境变化重点实验室师生对野外采样、样品处理及数据分析提供帮助,特此致谢。
[1] Lampert W, Sommer U. Limnoecology. 2nd ed. New York: Oxford University Press, 2007: 14- 15.
[2] Walker H W. Harmful Algae Blooms in Drinking Water: Removal of Cyanobacterial Cells and Toxins. Boca Raton: CRC Press, 2014: 2- 3.
[3] 马健荣, 邓建明, 秦伯强, 龙胜兴. 湖泊蓝藻水华发生机理研究进展. 生态学报, 2013, 33(10): 3020- 3030.
[4] Leavitt P R, Hodgson D A. Sedimentary pigments // Smol J P, Birks H J B, Last W M, Bradley R S, Alverson K, eds. Tracking Environmental Change Using Lake Sediments. Netherlands: Springer, 2001: 295- 325.
[5] 刘建康. 高级水生生物学. 北京: 科学出版社, 1999: 172- 174.
[6] 卡尔夫. 湖沼学: 内陆水生态系统. 古滨河, 刘正文, 李宽意, 译. 北京: 高等教育出版社, 2011: 48- 49.
[7] 秦伯强, 许海, 董百丽. 富营养化湖泊治理的理论与实践. 北京: 高等教育出版社, 2011: 32- 33.
[8] Vogt R J, Sharma S, Leavitt P R. Decadal regulation of phytoplankton abundance and water clarity in a large continental reservoir by climatic, hydrologic and trophic processes. Journal of Great Lakes Research, 2014, 41(S2): 81- 90.
[9] 黎尚豪, 俞敏娟, 李光正, 曾继绵, 陈嘉佑, 高宝云, 黄宏金. 云南高原湖泊调查. 海洋与湖沼, 1963, 5(2): 87- 114.
[10] 董云仙. 洱海蓝藻水华研究. 云南环境科学, 1999, 18(4): 28- 31.
[11] 王芸. 洱海夏秋季蓝藻种群动态变化及水华成因分析. 大理学院学报, 2008, 7(12): 39- 42.
[12] 吕兴菊, 朱江, 孟良. 洱海水华蓝藻多样性初步研究. 环境科学导刊, 2010, 29(3): 32- 35.
[13] 王云飞, 潘红玺, 吴庆龙, 黄群. 人类活动对洱海的影响及对策分析. 湖泊科学, 1999, 11(2): 123- 128.
[14] 颜昌宙, 金相灿, 赵景柱, 沈兵, 李宁波, 黄昌筑, 熊仲华. 云南洱海的生态保护及可持续利用对策. 环境科学, 2005, 26(5): 38- 42.
[15] 李原, 李任伟, 尚榆民, 李宁波. 云南洱海的环境沉积学研究——表层沉积物的粒度分布、水流方向和能量. 沉积学报, 1999, 17(S1): 769- 774.
[16] 申慧彦, 李世杰, 舒卫先. 湖泊沉积物中色素的研究及其环境指示意义. 海洋地质与第四纪地质, 2007, 27(3): 37- 42.
[17] 张振克, 吴瑞金, 朱育新, 潘红玺, 吴艳宏, 夏威岗, 李徐生. 云南洱海流域人类活动的湖泊沉积记录分析. 地理学报, 2000, 55(1): 66- 74.
[18] Leavitt P R, Vinebrooke R D, Donald D B, Smol J P, Schindler D W. Past ultraviolet radiation environments in lakes derived from fossil pigments. Nature, 1997, 388(6641): 457- 459.
[19] Chen X, McGowan S, Xu L, Zeng L H, Yang X D. Effects of hydrological regulation and anthropogenic pollutants on Dongting Lake in the Yangtze floodplain. Ecohydrology, 2016, 9(2):315- 325.
[20] 梁洁, 侯居峙, 李栋, 王明达. 沉积色素定量重建湖泊系统变化研究进展. 第四纪研究, 2016, 36(3): 630- 645.
[21] Larkum A W D, Douglas S N E, Raven J A. Photosynthesis in Algae. Dordrecht: Springer Science+ Business Media, 2003.
[22] Poutanen E L, Nikkilä K. Carotenoid pigments as tracers of cyanobacterial blooms in recent and post-glacial sediments of the Baltic Sea. Ambio, 2001, 30(4/5): 179- 183.
[23] Guilizzoni P, Massaferro J, Lami A, Piovano E L, Guevara S R, Formica S M, Daga R, Rizzo A, Gerli S. Palaeolimnology of Lake Hess (Patagonia, Argentina): multi-proxy analyses of short sediment cores. Hydrobiologia, 2009, 631(1): 289- 302.
[24] Mcgowan S, Barker P, Haworth E Y, Leavitt P R, Maberly S C, Pates J. Humans and climate as drivers of algal community change in Windermere since 1850. Freshwater Biology, 2012, 57(2): 260- 277.
[25] 王苏民, 窦鸿身. 中国湖泊志. 北京: 科学出版社, 1998: 371- 374.
[26] 郑国强, 于兴修, 江南, 何隆华. 洱海水质的演变过程及趋势. 东北林业大学学报, 2004, 32(1): 99- 102.
[27] 彭文启, 王世岩, 刘晓波. 洱海水质评价. 中国水利水电科学研究院学报, 2005, 3(3): 192- 198.
[28] 于洋, 张民, 钱善勤, 李大命, 孔繁翔. 云贵高原湖泊水质现状及演变. 湖泊科学, 2010, 22(6): 820- 828.
[29] 杜宝汉. 洱海富营养化研究. 湖泊科学, 1992, 4(2): 86- 92.
[30] 吴功果, 倪乐意, 曹特, 谢平, 徐军. 洱海水生植物与浮游植物的历史变化及影响因素. 水生生物学报, 2013, 37(5): 912- 917.
[31] Rong W, Dearing J A, Langdon P G, Zhang E L, Yang X D, Dakos V, Scheffer M. Flickering gives early warning signals of a critical transition to a eutrophic lake state. Nature, 2012, 492(7429): 419- 422.
[32] 王宇. 云南气候变化概论. 北京: 气象出版社, 1996: 45- 56.
[33] 金相灿, 刘鸿亮, 屠清瑛, 章宗涉, 朱萱. 中国湖泊富营养化. 北京: 中国环境科学出版社, 1990: 379- 383.
[34] Cuddington K, Leavitt P R. An individual-based model of pigment flux in lakes: Implications for organic biogeochemistry and paleoecology. Canadian Journal of Fisheries and Aquatic Sciences, 1999, 56(10): 1964- 1977.
[35] Qin B, Liu Z, Havens K. Eutrophication of Shallow Lakes with Special Reference to Lake Taihu, China. Dordrecht: Springer Science+Business Media, 2007: 176- 178.
[36] 谢平. 论蓝藻水华的发生机制——从生物进化、 生物地球化学和生态学视点. 北京: 科学出版社, 2007: 78- 79.
[37] Xu H, Paerl H W, Qin B Q, Zhu G W, Gaoa G. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnology and Oceanography, 2010, 55(1): 420- 432.
[38] Schindler D W, Hecky R E, Findlay D L, Stainton M P, Parker B R, Paterson M J, Beaty K G, Lyng M, Kasian E M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: results of a 37-year whole-ecosystem experiment. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(32): 11254- 11258.
[39] 杜芳芳. 湖泊流域土地利用变化与湖泊水质关系研究[D]. 昆明: 昆明理工大学, 2011.
[40] 姚焕玫. 基于GIS技术的湖泊水质污染综合评价的研究[D]. 武汉: 武汉大学, 2005.
[41] 柯高峰, 丁烈云. 洱海流域城乡经济发展与洱海湖泊水环境保护的实证分析. 经济地理, 2009, 29(9): 1546- 1551.
[42] 储昭升, 叶碧碧, 田桂平, 庞燕, 胡小贞. 洱海沉水植物空间分布及生物量估算. 环境科学研究, 2014, 27(1): 1- 5.
[43] 李威, 何亮, 朱天顺,曹特, 张霄林, 倪乐意. 洱海苦草(Vallisneria natans)水深分布和叶片C、N、P化学计量学对不同水深的响应. 湖泊科学, 2014, 26(4): 585- 592.
[44] Meyers P A, Lallier-vergés E. Lacustrine sedimentary organic matter records of late quaternary paleoclimates. Journal of Paleolimnology, 1999, 21(3): 345- 372.
[45] Cohen A S. Paleolimnology: the History and Evolution of Lake Systems. New York: Oxford University Press, 2003.
[46] 周志华, 李军, 朱兆洲. 龙感湖沉积物碳、氮同位素记录的环境演化. 生态学杂志, 2007, 26(5): 693- 699.
[47] Kenney W F, Waters M N, Schelske C L, Brenner M. Sediment records of phosphorus-driven shifts to phytoplanktondominance in shallow Florida lakes. Journal of Paleolimnology, 2002, 27(3): 367- 377.
[48] 张强, 韩永翔, 宋连春. 全球气候变化及其影响因素研究进展综述. 地球科学进展, 2005, 20(9): 990- 998.
[49] Adrian R, O′Reilly C M, Zagarese H, Baines S B, Hessen D O, Keller W, Livingstone D M, Sommaruga R, Straile D, Van Donk E, Weyhenmeyer G A, Winder M.. Lakes as sentinels of climate change. Limnology and Oceanography, 2009, 54(6part2): 2283- 2297.
[50] 傅国斌, 李克让. 全球变暖与湿地生态系统的研究进展. 地理研究, 2001, 20(1): 120- 128.
[51] 杨小梅, 安文玲, 张薇. 中国西南地区日照时数变化及影响因素. 兰州大学学报: 自然科学版, 2012, 48(5): 52- 60.
[52] 段旭, 陶云, 杜军, 李永华, 杨淑群, 段长春, 郑建萌, 段玮, 陈艳, 任菊章. 西南地区气候变化基本事实及极端气候事件. 北京: 气象出版社, 2011: 156- 178.
[53] 秦红杰. 水华蓝藻对环境胁迫的适应性机制研究[D]. 北京: 中国科学院大学, 2013.
[54] Whitton B A. Ecology of Cyanobacteria II: Their Diversity in Space and Time. New York: Springer Science+Business Media, 2012: 216- 222.
[55] Huber V, Wagner C, Gerten D, Adrian R. To bloom or not to bloom: contrasting responses of cyanobacteria to recent heat waves explained by critical thresholds of abiotic drivers. Oecologia, 2012, 169(1): 245- 256.
[56] Paerl H W, Huisman J. Blooms like it hot. Science, 2008, 320(5872): 57- 58.
[57] [邓建明, 秦伯强. 全球变暖对淡水湖泊浮游植物影响研究进展. 湖泊科学, 2015, 27(1): 1- 10.
[58] 郑维发, 曾昭琪. 淡水蓝藻的高温适应. 湖泊科学, 1994, 6(4): 356- 363.
[59] 吴庆龙, 王云飞. 洱海生物群落的历史演变分析. 湖泊科学, 1999, 11(3): 267- 273.
[60] 陈建良, 胡明明, 周怀东, 王雨春, 王英才, 李艳辉, 刘永定, 高继军, 洪宇宁. 洱海蓝藻水华暴发期浮游植物群落变化及影响因素. 水生生物学报, 2015, 39(1): 24- 28.
[61] 朱荣, 王欢, 余得昭, 曾诚, 沈宏, 陈隽. 2013年洱海水华期间微囊藻毒素和浮游植物动态变化. 湖泊科学, 2015, 27(3): 378- 384.
[62] 商兆堂, 任健, 秦铭荣, 夏瑛, 何浪, 陈钰文. 气候变化与太湖蓝藻暴发的关系. 生态学杂志, 2010, 29(1): 55- 61.
[63] Kosten S, Huszar V L M, Bécares E, Costa L S, van Donk E, Hansson L A, Jeppesen E, Kruk C, Lacerot G, Mazzeo N, De Meester L, Moss B, Lürling M, Nõges T, Romo S, Scheffer M. Warmer climates boost cyanobacterial dominance in shallow lakes. Global Change Biology, 2012, 18(1): 118- 126.
[64] Tadonléke R D. Evidence of warming effects on phytoplankton productivity rates and their dependence on eutrophication status. Limnology and Oceanography, 2010, 55(3): 973- 982.
[65] Taranu Z E, Zurawell R W, Pick F, Gregory-Eaves I. Predicting cyanobacterial dynamics in the face of global change: the importance of scale and environmental context. Global Change Biology, 2012, 18(12): 3477- 3490.
[66] 黄邦钦, 胡俊, 柳欣, 张彩云, 周立斌. 全球气候变化背景下浮游植物群落结构的变动及其对生物泵效率的影响. 厦门大学学报: 自然科学版, 2011, 50(2): 402- 410.
[67] 刘洪波, 朱梦羚, 王精志, 顾海欣. 水库水动力对水体富营养化影响. 水资源与水工程学报, 2013, 24(2): 19- 21.
[68] 杨柳燕, 肖琳. 湖泊蓝藻水华暴发、危害与控制. 北京: 科学出版社, 2011: 130- 147.
[69] 王丽平, 郑丙辉, 张佳磊, 刘晓霭. 三峡水库蓄水后对支流大宁河富营养化特征及水动力的影响. 湖泊科学, 2012, 24(2): 232- 237.
[70] Zhao Y, Xia X, Yang Z. Growth and nutrient accumulation of Phragmites australis in relation to water level variation and nutrient loadings in a shallow lake. Journal of Environmental Sciences: China, 2013, 25(1): 16- 25.
[71] Scheffer M. Ecology of Shallow Lakes. New York: Springer Science+Business Media, 2004.
[72] Loverde-Oliveira S M, Huszar V L M, Mazzeo N. Hydrology-driven regime shifts in a shallow tropical lake. Ecosystems, 2009, 12: 807- 807.
[73] 符辉, 袁桂香, 曹特, 钟家有, 张霄林, 过龙根, 张萌, 倪乐意, 王圣瑞. 洱海近50a来沉水植被演替及其主要驱动要素. 湖泊科学, 2013, 25(6): 854- 861.
[74] 李恒, 尚榆民. 云南洱海水生植被. 山地研究, 1989, 7(3): 166- 174.
[75] 厉恩华, 王学雷, 蔡晓斌, 王晓艳, 赵素婷. 洱海湖滨带植被特征及其影响因素分析. 湖泊科学, 2011, 23(5): 738- 746.
[76] 戴全裕. 云南抚仙湖、洱海、滇池水生植被的生态特征. 生态学报, 1985, 5(4): 324- 335.
Spatio-temporal patterns and drivers of cyanobacteria blooms in Erhai Lake, Southwest China
WANG Jiaoyuan1, CHEN Guangjie1,*, HUANG Linpei1, LIU Xiaodong2, CHEN Qianqian2, LIU Xiaohai3, ZHANG Hucai1
1LaboratoryofPlateauGeographicalProcessesandEnvironmentalChanges,YunnanNormalUniversity,Kunming650500,China2InstituteofPolarEnvironment,UniversityofScienceandTechnologyofChina,Hefei230026,China3YunnanInstituteofEnvironmentalScience,Kunming650034,China
Lake environments and ecosystems are known to be sensitive to climate change and human activities. Erhai, a large plateau lake (~250 km2), is characterized by complicated lake morphometry and has experienced multiple environmental stressors (such as eutrophication, water regulation, etc.) over the past few decades. These have led to the deterioration of its water quality and loss of ecosystem resilience, where cyanobacteria blooms have frequently occurred since the 1990s. To uncover the long-term history and spatio-temporal pattern of cyanobacterial blooms in Erhai Lake, multi-proxy analyses of lake sediments from three different parts of the lake basin were conducted and compared. Based on the high-resolution sedimentary pigment data from each core, we found that there existed a strong asynchrony in the long-term changes of algal production among the cores. The significant increase of cyanobacterial pigments in the shallower southern part of the basin started from the late 1980s, earlier than those of the central and northern parts of the lake basin (~1990 and 2000, respectively), with the whole-basin showing cyanobacteria blooms starting from ca. AD 2000. Furthermore, minimum adequate models built to explain the changes in the pigment concentration in the sediments, suggests that climate warming and eutrophication were the main factors that led to the cyanobacteria blooms across the lake basin.In addition, cyanobacteria bloom for the southern part of the lake basin was also related to the water-level fluctuation, hydrodynamics, and macrophyte changes over the last few decades. Overall, our sedimentary records showed that there existed strong heterogeneity in the spatio-temporal patterns of cyanobacterial blooms and the driving factors also varied across space and over the last century in Erhai Lake.. Therefore, taking the current global warming scenario into account, control of both nutrient input and water regulation should be highlighted for ecological restoration and sustainable management of Erhai Lake
sediment pigments; cyanobacteria blooms; spatio-temporal heterogeneity; Erhai Lake
国家自然科学基金项目(U1133601,41171048,41302151);教育部科学技术研究项目(213034A);云南省中青年学术技术带头人培养计划(2012HB012);云南省创新团队项目(2011C1);云南省高端人才项目(2010CI111)
2016- 04- 20; 网络出版日期:2017- 03- 02
10.5846/stxb201604200740
*通讯作者Corresponding author.E-mail: guangjiechen@gmail.com
王教元,陈光杰,黄林培,刘晓东,陈倩倩,刘晓海,张虎才.洱海蓝藻爆发的时空特征及影响因子.生态学报,2017,37(14):4831- 4842.
Wang J Y, Chen G J, Huang L P, Liu X D, Chen Q Q, Liu X H, Zhang H C.Spatio-temporal patterns and drivers of cyanobacteria blooms in Erhai Lake, Southwest China.Acta Ecologica Sinica,2017,37(14):4831- 4842.
猜你喜欢
当代水产(2021年8期)2021-11-04 08:49:00
青海大学学报(2021年3期)2021-06-19 07:12:56
黄河之声(2020年11期)2020-07-16 13:27:12
作文成功之路·小学版(2020年4期)2020-01-02 10:49:54
西南石油大学学报(自然科学版)(2019年4期)2019-11-04 00:34:10
当代水产(2019年8期)2019-10-12 08:58:14
当代水产(2019年9期)2019-10-08 08:02:42
湿地科学与管理(2019年1期)2019-04-03 07:53:08
经济技术协作信息(2018年18期)2019-01-23 07:16:56
海峡姐妹(2017年5期)2017-06-05 08:53:17
