
北黄海獐子岛海域浮游动物群落年际变化
2017-08-10 12:17:13尹洁慧张光涛李超伦王世伟万艾勇
海洋学报 2017年8期
尹洁慧,张光涛,李超伦,王世伟,万艾勇
(1. 烟台大学 海洋学院,山东 烟台264005;2. 山东胶州湾海洋生态系统国家野外科学观测研究站,山东 青岛 266071;3. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071)
北黄海獐子岛海域浮游动物群落年际变化
尹洁慧1,张光涛2,李超伦3,王世伟2,万艾勇2
(1. 烟台大学 海洋学院,山东 烟台264005;2. 山东胶州湾海洋生态系统国家野外科学观测研究站,山东 青岛 266071;3. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071)
本文比较了海洋普查期间(1959年1-12月)和2009-2010年獐子岛海域附近站位的浮游动物的种类组成、优势种、丰度、生物多样性,结果显示,北黄海区域浮游动物群落物种组成未发生较大的变化,在獐子岛海域以及整个北黄海,中华哲水蚤和强壮箭虫的优势种地位没有发生变化,细足法虫戎也一直出现且为冬春季的优势种;太平洋磷虾在2009-2010年獐子岛海域虽然全年均有出现,但是全年均不是优势种,而在1959年的獐子岛海域,作为优势种出现在4月和11月;1959年与2009年相较,腹针胸刺水蚤在春夏季优势种的地位被沃氏纺锤水蚤所取代。2009年浮游动物丰度(131.26 ind/m3)比1959年(78.90 ind/m3)高;浮游动物多样性指数(H′)均在夏秋季高于冬春季节,且年平均多样性指数也有所提高。
浮游动物;优势种;丰度;生物多样性;獐子岛
1 引言
北黄海主要指位于山东半岛、辽东半岛和朝鲜半岛之间的半封闭区域,獐子岛海域位于辽东半岛东侧,黄海北部,是北黄海重要的组成部分,该地区的物理环境较为复杂,冬季受到黄海暖流的影响,夏季则受黄海冷水团影响[1]。该地区的河流都是季节性河流,冬季呈现枯水,夏季出现洪水。相对于南黄海而言,北黄海区域的浮游动物研究资料相对较少,针对獐子岛海域的生态研究更是很少有报道,背景资料缺乏。
北黄海拥有我国最大的虾夷扇贝养殖基地和以大连獐子岛集团为代表的海水养殖企业,是我国重要的水产品生产基地[2]。近些年北黄海出现了一系列的生态问题,如水母和被囊类等胶质生物的大量发生等[3],使得该地区的生态研究受到了关注。
浮游动物是海洋食物链中承上启下的关键环节,是具有调控作用的关键功能群,浮游动物群落的组成及其变化直接影响着海洋生态系统能流、物流的方向和效率[4]。以往关于獐子岛海域浮游动物的资料几乎没有,北黄海浮游动物的研究主要集中于个别月份,如20世纪50-60年代北黄海烟威鲐鱼渔场及邻近水域的浮游动物的初步研究[5], 80-90年代北黄海浮游动物的垂直移动规律和小长山群岛浮游动物以及大连湾的桡足类和枝角类研究[6-8]。近些年,对北黄海浮游动物的研究[9-12]主要是季度月的调查,北黄海的周年调查起源于1958-1959年全国海洋综合普查,至今没有再进行过周年调查。
本文以獐子岛海域为代表,比较1958-1959年北黄海大面站位的浮游动物周年数据以及所对应獐子岛附近站位浮游动物的周年数据与2009年所进行的獐子岛海域浮游动物周年调查的数据,以浮游动物种类、优势种、丰度及物种多样性为指标,研究北黄海獐子岛海域浮游动物群落的变化,为解决生态问题提供一些背景资料。
2 材料与方法
2.1 调查时间及站位布设
1959年1-12月海洋普查在北黄海布设的大面站位(图1a)所示,共计67个站位,獐子岛海域对应的站位为图中所圈站位,共计4个,2009年獐子岛海域调查站位13个(图1b)。
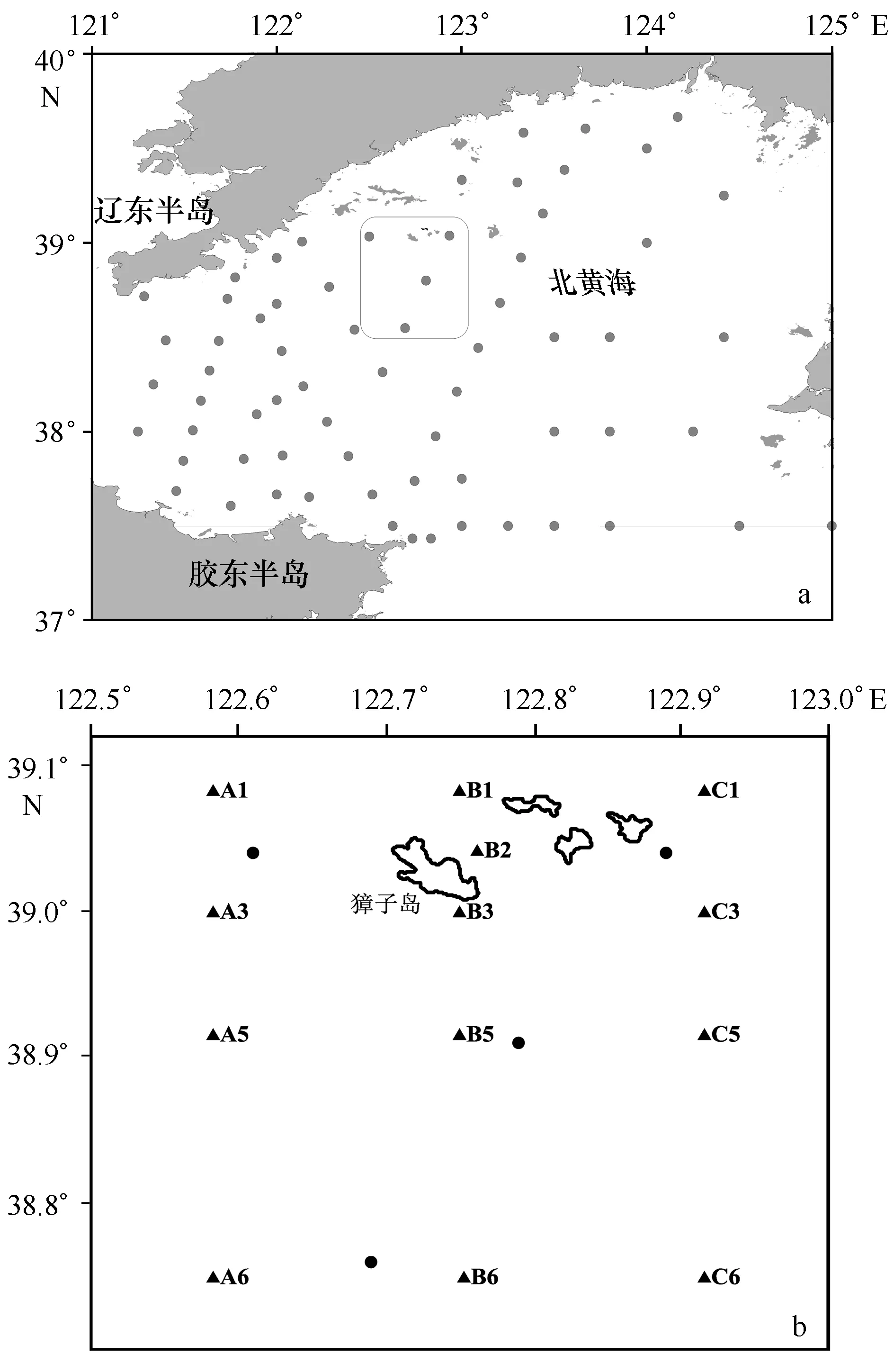
图1 采样站位图Fig.1 Sample sites of cruisesa.1959年北黄海大面站位, 方框为2009年调查区域;b.2009-2010年獐子岛调查区域,·代表1959年位于獐子岛海域附近的站位a.Sites in 1959 in North Yellow Sea. The circled area located around Zhangzi Island waters. b.Sites around Zhangzi Island from 2009-2010.·means sites around Zhangzi Island in 1959
2.2 样品采集及测定方法
样品采用标准大型浮游生物网(网口面积为0.5 m2,孔径为500 μm),进行从底到表的垂直拖网,获取的浮游动物用5%的甲醛溶液进行固定。
2.3 数据分析
将各站位大型浮游生物网采集样品作定量分析,根据采样时的滤水体积,各种类的丰度(abundance)以每立方米水体中的个体数(ind/m3)表示。
使用多元统计软件PRIMER V5.2 (Plymouth Routines In Multivariate Ecological Research)进行生物多样性分析,其中种类多样性指数采用香农-威弗指数(Shannon-Weaver Index)[13],公式为:
(1)
式中,H′为群落的多样性指数,Pi为样品中属于第i种的个体比例,如样品总个体为N,第i种的个体数为ni则Pi=ni/N。
丰富度指数(Margalef′s index,D),它综合了样品中种类数目和丰度的信息,表示一定动物丰度中的物种数,计算公式为:
D=(S-1)/log2N,
(2)
式中,N为丰度。
均匀度指数(Pielou′s evenness index,J)计算公式为:
J′=H′/log2S.
(3)
浮游动物的优势种根据每个种的优势度值(Y)来确定,优势度值根据各物种出现的频数及个体数量来计算[13],其中Y=(ni/N)×fi,式中,ni为第i种的个体数,N为所有种类总个体数,fi为出现频率,Y≥0.02的种类为优势种[14]。
3 结果
3.1 浮游动物种类组成
1959年海洋普查期间(表1A)共出现浮游动物51种,浮游幼虫10种,由于多毛类、糠虾、十足目、涟虫目等都没有记录到具体的种,所以与实际组成相比,种类数偏低。全年均有出现的浮游动物有中华哲水蚤(Calanussinicus)、强壮箭虫(Sagittacrassa)、真刺唇角水蚤(Labidoceraeuchaeta)、太平洋磷虾(Euphausiapacifica)、细足法虫戎(Themistogracilipes)、 涟虫、糠虾、钩虾,出现频率较多的物种有百陶箭虫(Sagittabedoti)、腹针胸刺水蚤(Centropagesabdominalis)、双刺唇角水蚤(Labidocerabipinnata)、汤氏长足水蚤(Calanopiathompsoni)、瘦尾胸刺水蚤(Centropagestenuiremis)、宽额假磷虾(Pseudeuphausialatifrons)、细鳌虾(Leptochelagracilis)、四辐枝管水母(Proboscidactylaflavicirrat)、脆弱似杯水母(Phialellafragilis)、半球美螅水母(Clytiahemisphaerica)、五角水母(Muggiaeaatlantica)、长尾类幼体、磁蟹蚤状幼体、鱼卵。
海洋普查期间对应的獐子岛海域的4个站位共出现浮游动物31种(表1B),浮游幼虫6种,糠虾目、十足目、涟虫目没有记录到具体的种。其中桡足类种类最多12种,占种类组成的38.7%,毛颚类3种,占种类组成的9.7%,磷虾类2种,占种类组成的6.5%,刺胞动物6种,占种类组成的19.4%,栉水母1种,占种类组成的3.2%。其中中华哲水蚤、强壮箭虫在全年均有出现,出现频率较多的物种包括细足法虫戎、双刺唇角水蚤、太平洋磷虾、腹针胸刺水蚤、钩虾。水母类主要集中在夏秋季。由于此区域内覆盖的站位较少,所以种类组成相对较为单一。
2009年的调查中,獐子岛海域共出现浮游动物共29种(表1C),其中桡足类11种,占种类组成的30.6%;毛颚类3种,占种类组成的8.3%;水螅水母类、栉水母类、枝角类、被囊类、莹虾类各2种,占种类组成的5.5%;糠虾类、磷虾类、端足类、原生动物各1种,占种类组成的2.7%。由于布设站位海域面积较小,浮游动物种类组成较为简单,中华哲水蚤、强壮箭虫、双刺唇角水蚤、细足法虫戎、太平洋磷虾在全年均有出现。一些广温广盐的枝角类如鸟喙尖头溞、肥胖三角溞,主要在夏秋季的7-10月出现,一些暖温带种如肥胖箭虫、百陶箭虫、中型莹虾主要在10月、11月在调查海域出现。水母在7月、9月,10月及11月出现,其中10月、11月出现了瓜水母的爆发。在整个调查过程中,没有发现真正意义上的暖水种。

表1 浮游动物种类组成

续表1

B.1959年獐子岛海域

C.2009-2010年獐子岛海域
3.2 浮游动物优势种
中华哲水蚤和强壮箭虫是海洋普查北黄海大面站位中全年的优势种(2A)。中华哲水蚤的优势度在7月份最高(0.67),在其余月份较为平均(0.26~0.47)。强壮箭虫的优势度最高出现在1月(0.66),2月份(0.64)和11月份(0.60)优势度值也较高,在其他月份优势度值在0.15~0.55。细足法虫戎除2月、9-11月外为优势种,最高值出现在5月和7月值为0.06,整体优势度较低,其余月份优势度值范围为0.02~0.04。腹针胸刺水蚤作为优势种,主要出现在春夏季(3-7月),优势度最高值(0.12)出现在5月,其他月份优势度范围为0.02~0.11;双刺唇角水蚤(Labidocerabipinnata)作为优势种只出现在9月,优势度为0.05。
如表2B,中华哲水蚤和强壮箭虫在海洋普查过程对应的獐子岛站位中也是全年的优势种,中华哲水蚤的优势度(0.71)在7月份最高,仅4月和9月优势度在0.3以下,强壮箭虫优势度在1月、2月较高,分别达到了0.70和0.64,细足法虫戎除1月、2月、9月外,其余月份均作为优势种出现,腹针胸刺水蚤作为优势种,主要出现在春夏季(3-7月),优势度最高(0.40)出现在4月,太平洋磷虾为4月、12月的优势种,双刺唇角水蚤、沃氏纺锤水蚤(Acartiaomorii)、拟长腹剑水蚤(Oithonasimilis)、长尾类幼体只在个别月份作为优势种出现,五角水母为9月份优势种。
在2009年獐子岛海域附近站位的调查中,中华哲水蚤和强壮箭虫是海域全年的优势种,如表2C。中华哲水蚤的优势度在6月、7月、8月的时候分别达到了0.93、0.86、0.70。 强壮箭虫的优势度次之,在10月和11月的时候优势度分别为0.52和0.42。细足法虫戎是冬春季的优势种,优势度最高达到0.26,双刺唇角水蚤、沃氏纺锤水蚤、夜光虫、长腕类幼体是春季的优势种,它们的优势度最高分别达到了0.06、0.07、0.03 和 0.23。鸟喙尖头溞较偏好低盐的环境,作为优势种主要出现在受冲淡水影响后的9月,优势度为0.02。
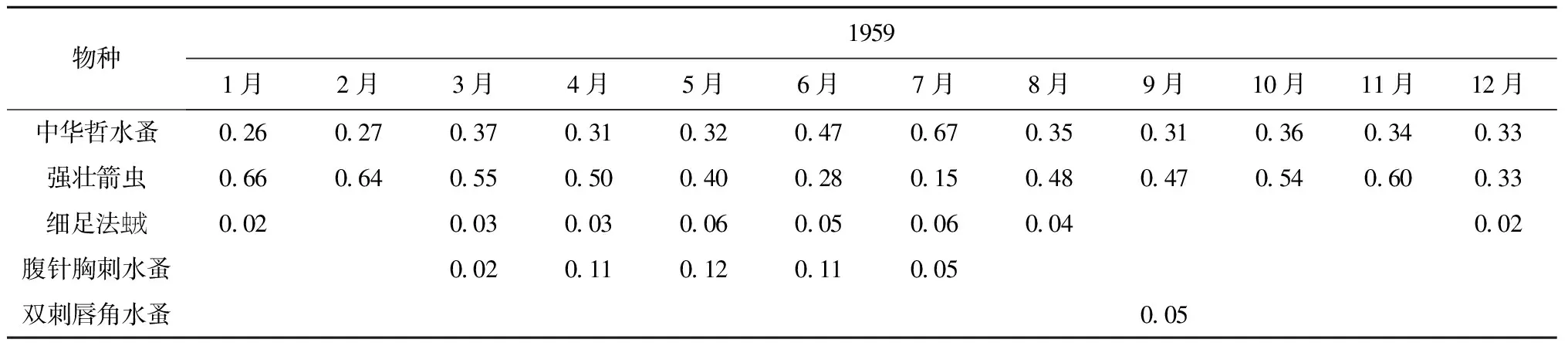
表2 浮游动物优势种及其优势度
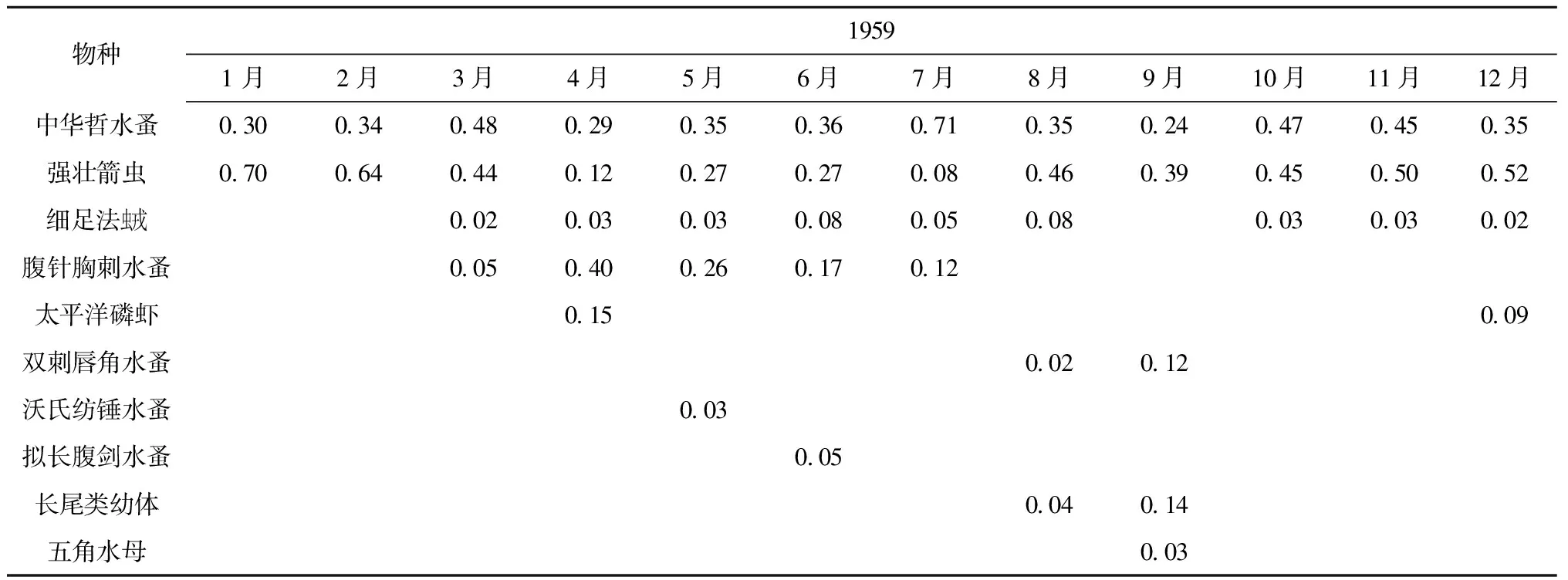
B.1959年獐子岛海域
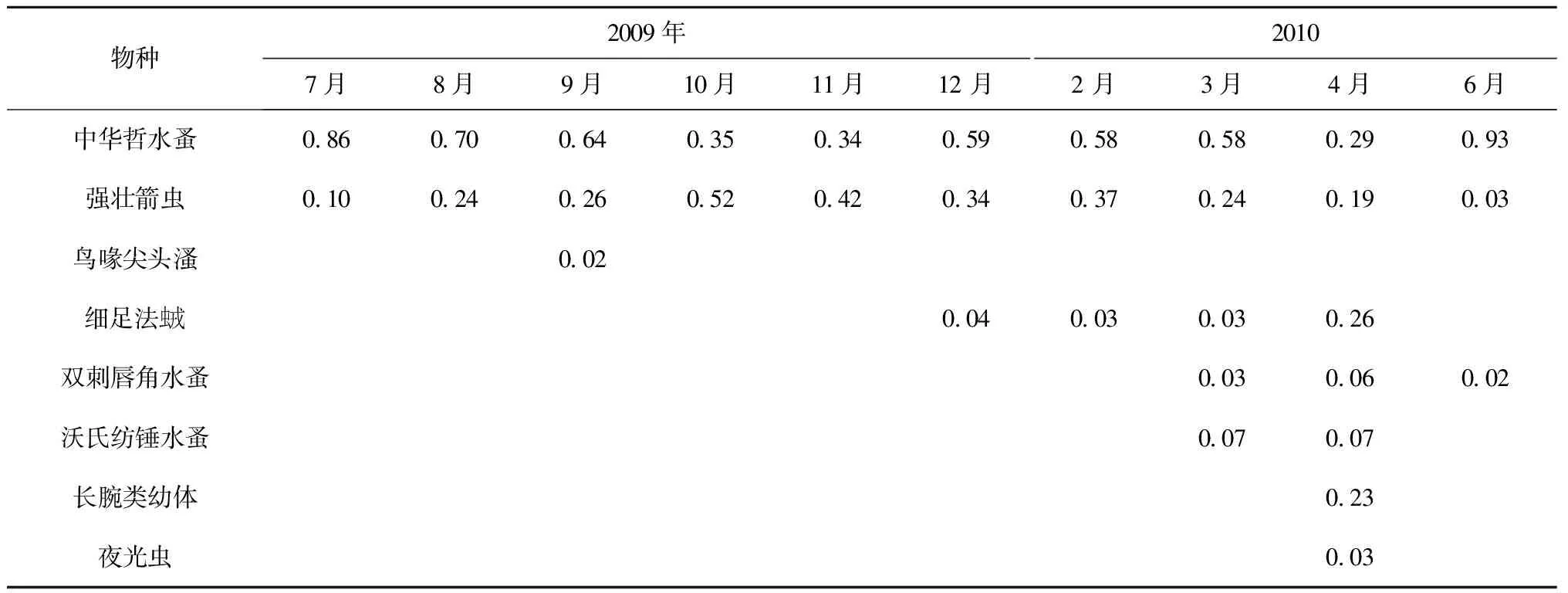
C.2009—2010年獐子岛海域
3.3 浮游动物丰度周年变化
海洋普查期间大面站位的浮游动物丰度在12月份丰度最高(134.95 ind/m3)(图2),5月份丰度(60.18 ind/m3)最低。浮游动物丰度在8-12月期间,值在100 ind/m3以上,从1-7月,浮游动物丰度低于100 ind/m3,范围在60.18~96.10 ind/m3之间,浮游动物全年平均丰度为94.58 ind/m3。
海洋普查期间獐子岛海域浮游动物丰度呈双周期波动,主高峰值出现在9月,丰度为167.2 ind/m3,次高峰值出现在1月,丰度为140.4 ind/m3,浮游动物丰度全年平均值为78.9 ind/m3,全年最低值出现在5月,其值为56.0 ind/m3。
2009年獐子岛海域浮游动物丰度呈现双周期波动, 浮游动物丰度主峰值出现在6月,丰度值为531.6 ind/m3,次高峰出现在7月与12月,其值分别为142.1 ind/m3和133.1 ind/m3,这与优势种中华哲水蚤和强壮箭虫在全年的丰度高峰值的分布一致。浮游动物丰度的最低值(38.19 ind/m3)均出现8月,浮游动物丰度年平均值为131.3 ind/m3。
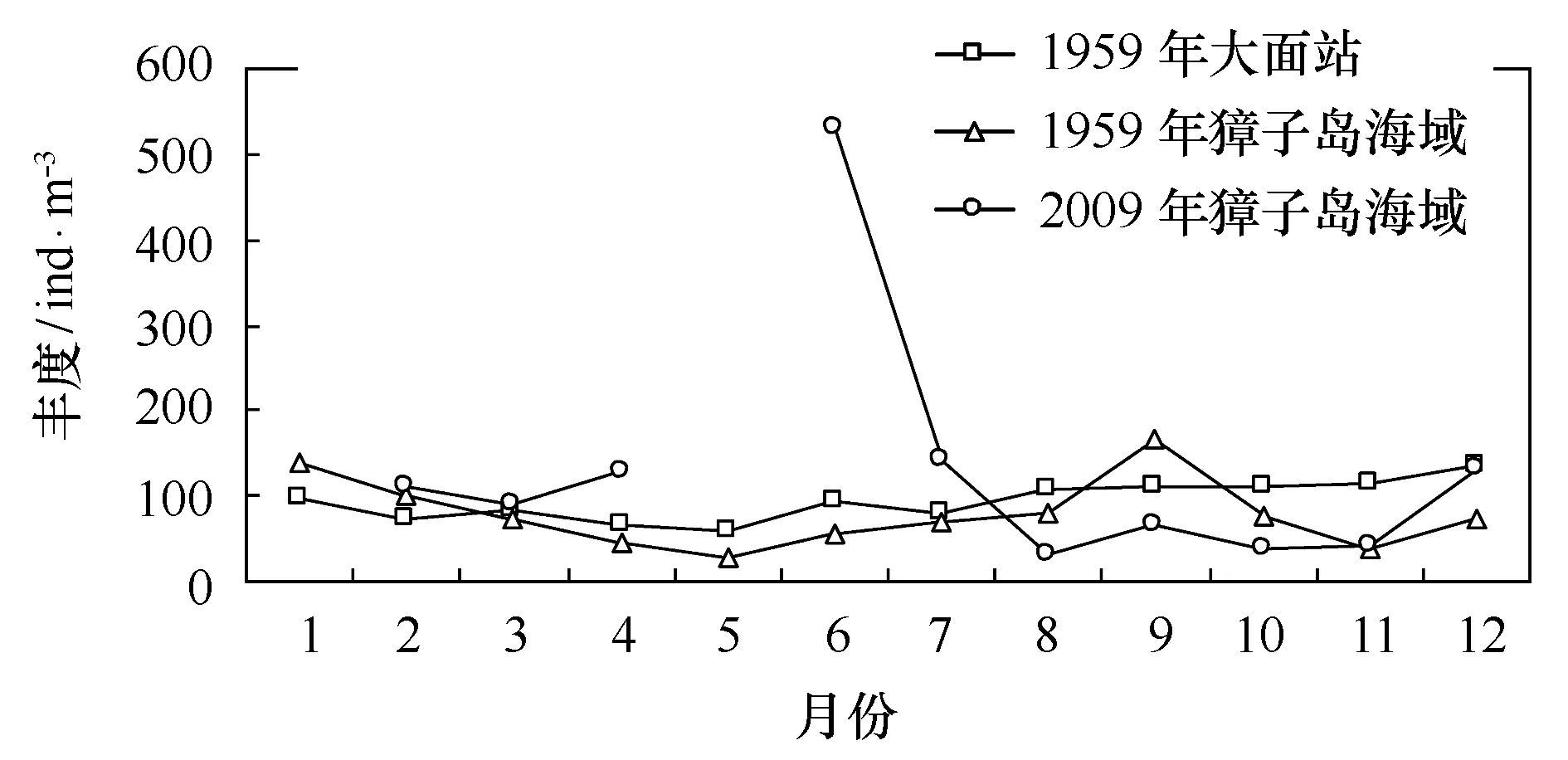
图2 北黄海及獐子岛海域浮游动物丰度年际变化Fig.2 Interannual variations of zooplankton abundance in North Yellow Sea and Zhangzi Island
3.4 浮游动物多样性指数
海洋普查期间,无论在北黄海大面站位还是与獐子岛海域相对应的区域,均呈现多样性指数(H′)和均匀度指数(J′)趋势一致,种类数量(S)和丰富度指数(D)一致的趋势。在大面站位(图3a),浮游动物种类数在6月、9月、12月为36种,种类数最多,在冬季2月种类数最少为17种,多样性指数和均匀度指数在5月份值最高,分别为1.19和0.60,多样性指数在2月份最低,值为0.78,均匀度指数在1月份最低,值为0.42,丰富度指数在9月值最高,为1.18,冬季2月份值最低为0.66。
由于獐子岛附近站位较少,所以浮游动物种类数不高,生物多样性也相应的不高,浮游动物在9月份种类最多(图3b),为21种,冬季1、2月份种类最少,仅有6种,这与站位较少有很大关系。浮游动物生物多样性指数,6月最高,为1.55,丰富度指数10月最高为1.49,均匀度指数较为平均,全年在0.42~0.82波动,这主要与北黄海区域浮游动物种类较单一有关。
在獐子岛海域2009-2010年的调查中(图3c),年平均多样性指数为1.39,丰富度指数为0.95,均匀度指数为0.49,呈现多样性指数、均匀度分布趋势一致,种类数量和丰富度一致的趋势,全年10月份种类数最多,丰富度指数最高为1.56,多样性指数和均匀性指数最高值出现在4月,分别为1.81和0.67。
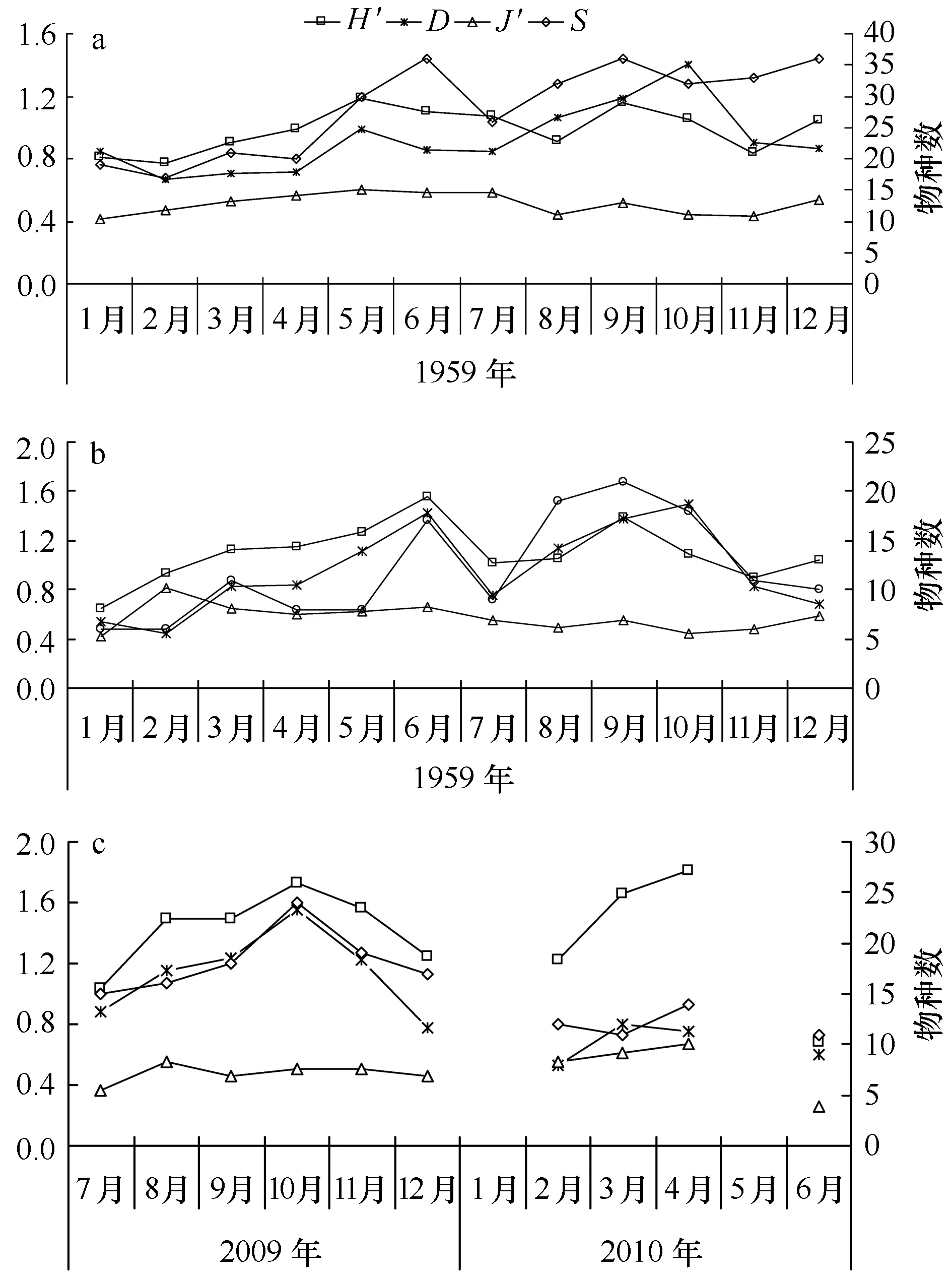
图3 浮游动物多样性周年变化Fig.3 Annual variations of zooplankton diversitya.1959年北黄海海域,b.1959年獐子岛海域,c.2009-2010年獐子岛海域a.North Yellow Sea in 1959,b. Zhangzi Island area in 1959, c. Zhangzi Island area from 2009 to 2010
4 讨论
4.1 浮游动物组成的年际变化
由于1959年海洋普查调查中有很多浮游动物种类并没有具体到种,且为周年调查,不方便与季度调查进行细致的比较,并且这种季度调查的年间变化已有学者总结过[15],我们主要比较北黄海区域在从1959年到2009年在獐子岛海域浮游动物的时间方面的变化,以及獐子岛海域与整个北黄海区域的浮游动物优势种种类和丰度在空间上的区别。
在以往的关于北黄海季度月调查的研究中,2006-2007年调查的站位较多与1959年大体相似,调查中北黄海的浮游动物被划成4个类群,主要包括(Ⅰ)半咸水河口群落,(Ⅱ)辽宁近岸低盐群落,(Ⅲ) 黄海冷水团群落,(Ⅳ)山东半岛近岸群落[8-9]。而在北黄海海洋普查的浮游动物种类也大致分属于这4个群落。2009年,在獐子岛海域设置了13个站位,站位数和覆盖面积较小,浮游动物种类数也相对较少,但浮游动物种类分别分属于这4个群落(表1C),例如中华哲水蚤、细足法虫戎等在7-8月黄海冷水团丰度较高的种类属于种群Ⅲ,鸟喙尖头溞(Penetiaavirostris)、肥胖三角溞(Pseudevadnetergestina)等低盐类群属于种群Ⅰ,沃氏纺锤水蚤、拟长腹剑水蚤主要在春季出现,他们属于群落Ⅱ,所以在浮游动物组成方面,浮游动物种群和种类总数大体上没有变化。
海洋普查时期,对应獐子岛海域浮游动物共31种,2009年浮游动物共29种,而且在种类组成方面,均是桡足类占绝对优势,分别占了种类总数的38.7% 和30.6%。 在1959年普查期间,枝角类在獐子岛海域内的站位没有出现,水母出现的种类较为多样化,包括半球美螅水母、脆弱似杯水母、五角水母、耳状囊水母(Euphysaaurata),而在2009年调查中,在受冲淡水影响的月份内,出现了两种枝角类(鸟喙尖头溞、肥胖三角溞),水母爆发月份,水母种类单一化,仅包括瓜水母(Beroecucumis)和球型侧腕水母(Pleurobrachiaglobosa),可见獐子岛海域浮游动物的主要类群并未发生较大变化。
4.2 浮游动物优势种的年际变化
在优势种组成方面,中华哲水蚤和强壮箭虫一直都是獐子岛海域以及整个北黄海全年的优势种[9-12],中华哲水蚤在这两年均是在夏季6、7月的时候优势度值较高(图4a),全年优势度值平均值2009-2010年相对较高;强壮箭虫在这两年中冬季(12月、1月)丰度值较高,全年优势度平均值1959年相对较高。细足法虫戎一直作为该海域冬春季优势种出现;在1959年海洋普查期间,獐子岛海域太平洋磷虾全年大部分月份均有出现,并为4月和12月的优势种,而2009年在獐子岛海域,太平洋磷虾虽然全年均有出现,但是从未作为优势种出现;1959年, 腹针胸刺水蚤在海洋普查期间是春夏季的优势种,而在2009年这种优势种的地位已被沃氏纺锤水蚤取代。
4.3 浮游动物丰度的年际变化
1959年海洋普查北黄海大面站位浮游动物丰度,1-7月较低,8月-12月较高,而具体到獐子岛海域,在1959年与2009年浮游动物丰度均呈现双周期波动,高峰均出现在夏季和冬季(图2)。北黄海大面站位,獐子岛海域1959年及2009年浮游动物的平均丰度分别为94.58 ind/m3、78.90 ind/m3和131.26 ind/m3。虽然獐子岛海域浮游动物丰度在1959年和2009年均呈现双周期波动,但是在时间上獐子岛海域浮游动物的丰度是增加的,1959年獐子岛海域浮游动物丰度最高值为167.2 ind/m3,而2009年獐子岛海域浮游动物丰度最高值为531.63 ind/m3,丰度增幅较大。在空间上,相对于整个北黄海,2009年獐子岛海域的丰度也是偏高的,1959年北黄海大面站位的浮游动物丰度最高值仅134.95 ind/m3。
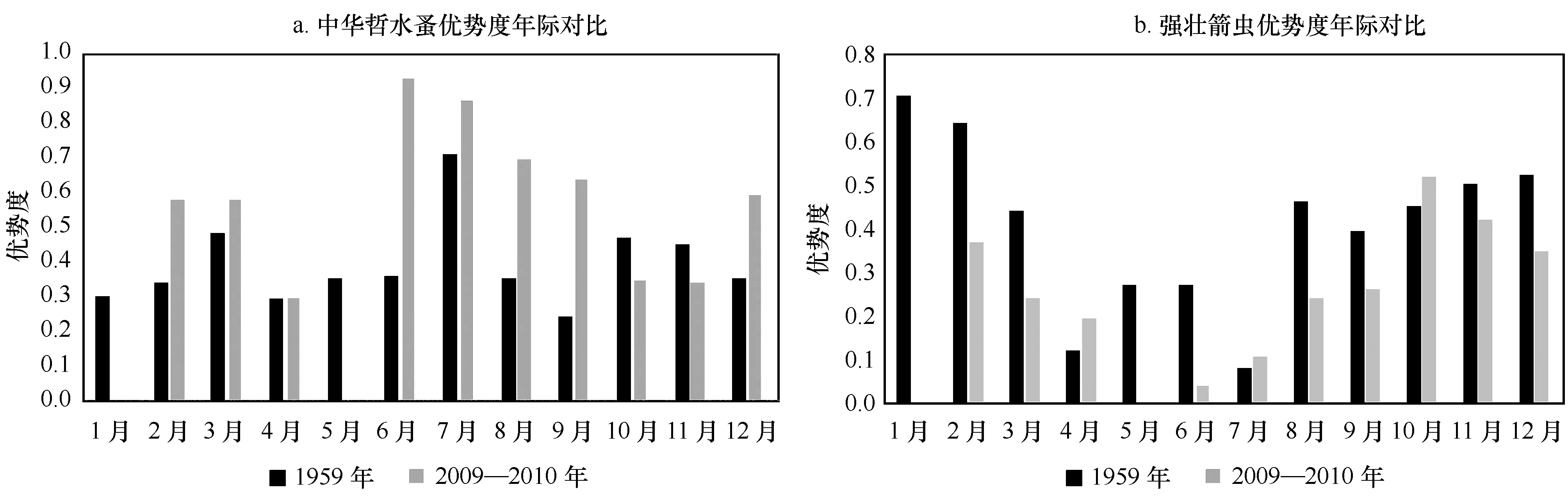
图4 优势种优势度年际变化Fig.4 Interannual varitions of dominant species dominance
4.4 浮游动物多样性的年际变化
在浮游动物多样性方面,无论是海洋普查北黄海大面站位还是獐子岛海域在1959年与2009年的调查,浮游动物种类数均是在夏秋季高于冬春季(图3),黄海冷水团的存在以及夏季径流的增加,使得环境因子(温度、盐度)变化更加多样化,增加了浮游动物的种类数,而随着秋季黄海冷水团的逐渐减弱,一些暖水种进入海域,使得生物多样性得以提高。1959年獐子岛海域浮游动物的生物多样性指数周年平均值为1.09,最高出现在6月,2009年平均值为1.35,最高值出现在4月,整体上獐子岛海域的生物多样性指数有所升高。
[1] 鲍献文, 李娜, 姚志刚, 等. 北黄海温盐分布季节变化特征分析[J]. 中国海洋大学学报, 2009, 39(4): 553-562.
Bao Xianwen, Li Na, Yao Zhigang, et al. Seasonal variation characteristics of temperature and salinity of the North Yellow Sea[J]. Periodical of Ocean University of China, 2009, 39(4): 553-562.
[2] 张继红, 方建光, 王诗欢. 大连獐子岛海域虾夷扇贝养殖容量[J]. 水产学报, 2008, 32(2): 236-241.
Zhang Jihong, Fang Jianguang, Wang Shihuan. Carrying capacity forPatinopectenyessoensisin Zhang Zidao Island, China[J]. Journal of Fisheries of China, 2008, 32(2): 236-241.
[3] 程家骅, 李圣法, 丁峰元, 等. 东、黄海大型水母暴发现象及其可能成因浅析[J]. 现代渔业信息, 2004, 19(5): 10-12.
Cheng Jiahua, Li Shengfa, Ding Fengyuan, et al. Primary analysis on the jellyfish blooms and its cause in the East China Sea and the Yellow Sea[J]. Modern Fisheries Information, 2004, 19(5): 10-12.
[4] 郑重, 李少菁, 许振祖. 海洋浮游生物学[M]. 北京: 海洋出版社, 1984.
Zheng Zhong, Li Shaojing, Xu Zhenzu. Mairine Zooplankton Biology[M]. Beijing: China Ocean Press, 1984.
[5] 郑重, 郑执中, 王荣, 等. 烟、威鲐鱼渔埸及邻近水域浮游动物生态的初步研究[J]. 海洋与湖沼, 1965, 7(4): 329-354.
Zheng Zhong, Zheng Zhizhong, Wang Rong, et al. Ecological investigations on the zooplankton of the mackerel fishing ground off Yantai-Weihai and adjacent waters[J]. Oceanologia et Limnologia Sinica, 1965, 7(4): 329-354.
[6] 王真良, 刘晓丹. 北黄海浮游动物昼夜垂直移动的初步研究[J]. 黄渤海海洋, 1989, 7(4): 50-54.
Wang Zhenliang, Liu Xiaodan. A preliminary research on diel vertical migration of zooplankton in North Huanghai Sea (Yellow Sea)[J]. Journal of Oceanography of Huanghai & Bohai Seas, 1989, 7(4): 50-54.
[7] 王真良. 大连湾水域桡足类的分布[J]. 黄渤海海洋, 1995, 13(1): 47-54.
Wang Zhenliang. Distribution of pelagic copepods in Dalian Bay[J]. Journal of Oceanography of Huanghai & Bohai Seas, 1995, 13(1): 47-54.
[8] 王真良. 小长山岛周围海域浮游动物群落结构的初步研究[J]. 大连水产学院学报, 2003, 18(4): 296-300.
Wang Zhenliang. Preliminary studies on zooplankton community in waters around Xiao Changshan Island[J]. Journal of Dalian Fisheries University, 2003, 18(4): 296-300.
[9] 朱延忠. 夏、冬季北黄海大中型浮游动物群落生态学研究[D]. 青岛: 中国海洋大学, 2008.
Zhu Yanzhong. Study on the macro-zooplankton and meso-zooplankton community ecology in the North Yellow Sea in summer and winter[D]. Qingdao: Ocean University of China, 2008.
[10] 姜强. 春、秋季北黄海大中型浮游动物群落生态学研究[D]. 青岛: 中国海洋大学, 2010.
Jiang Qiang. Study on the macro-zooplankton and meso-zooplankton community ecology in the North Yellow Sea in spring and autumn[D]. Qingdao: Ocean University of China, 2010.
[11] 杨青, 王真良, 樊景凤, 等. 北黄海秋、冬季浮游动物多样性及年间变化[J]. 生态学报, 2012, 32(21): 6747-6754.
Yang Qing, Wang Zhenliang, Fan Jingfeng, et al. Zooplankton diversity and its variation in the Northern Yellow Sea in the autumn and winter of 1959, 1982 and 2009[J]. Acta Ecologica Sinica, 2012, 32(21): 6747-6754.
[12] 邹艺伟, 杨青, 李全宝, 等. 北黄海浮游动物群落结构及年间比较[J]. 海洋环境科学, 2013, 32(5): 683-687.
Zou Yiwei, Yang Qing, Li Quanbao, et al. Community structure and variation of zooplankton in the Northern Yellow Sea[J]. Marine Environmental Science, 2013, 32(5): 683-687.
[13] Shannon C E, Weaver W. The Mathematical Theory of Communication[M]. Urbana, IL: The University of Illinois Press, 1949: 125.
[14] 孙儒泳. 动物生态学原理[M]. 2版. 北京: 北京师范大学出版社, 1992: 356-357.
Sun Ruyong. The Principle of Animals Ecology[M]. 2nd ed. Beijing: Beijing Normal University Press, 1992: 356-357.
[15] Yin Jiehui, Zhang Guangtao, Li Chaolun, et al. Community composition, abundance and biomass of zooplankton in Zhangzi Island waters, Northern Yellow Sea[J]. Chinese Journal of Oceanology and Limnology, 2016, doi: 10.1007/s00343-017-6135-0. (in Press)
Interannual variations of zooplankton in Zhangzi Island, North Yellow Sea
Yin Jiehui1, Zhang Guangtao2, Li Chaolun3, Wang Shiwei2, Wan Aiyong2
(1.OceanCollege,YantaiUniversity,Yantai264005,China; 2.JiaozhouBayMarineEcosystemResearchStation,Qingdao266071,China; 3.KeyLaboratoryofMarineEcologyandEnvironmentalSciences,InstituteofOceanology,ChineseAcademyofSciences,Qingdao266071,China)
The annual zooplankton data for large areas in the Northern Yellow Sea during the marine census period (1959.1-1959.12) was analyzed in comparison to the annual zooplankton data around Zhangzi Island of the year 2009 in the aspects of species composition, dominant species, biological abundance and biodiversity. The results showed that zooplankton community around Zhangzi Island of North Yellow Sea did not exhibit significant changes.CalanussinicusandSaggitacrassawere observed as firmly dominant species.Themistogracilipeswas collected constantly each month, but it was dominant only in the winter and spring. In 2009,Euphausiapacificawas sampled in each month, but did not appear as dominant species; however, in 1959, it was observed as the dominant species in April and November. The dominant position ofCentropagesabdominalisin 1959 was replaced byAcartiaomoriiin 2009 in the spring and summer.The average abundance of zooplankton increased from 78.90 ind/m3to 131.26 ind/m3from 1959 to 2009; the biodiversity index (H′) was higher both in the summer and autumn than in the spring and winter in these two years, and the average biodiversity index value also increased.
zooplankton; dominant species; abundance; biodiversity; Zhangzi Island
2016-10-09;
2017-04-19。
国家自然科学青年基金(41506153);中科院海洋所海洋生态与环境科学实验室开放基金项目;中国科学院战略性先导科技专项(A类)(XDA11020701)。
尹洁慧(1986—),女,内蒙古呼伦贝尔市人,博士,主要从事海洋浮游动物研究。E-mail:yinjiehui@sohu.com
Q958.8
A
0253-4193(2017)08-0078-11
尹洁慧,张光涛,李超伦,等. 北黄海獐子岛海域浮游动物群落年际变化[J].海洋学报,2017,39(8):78—88, doi∶10.3969/j.issn.0253-4193.2017.08.008
Yin Jiehui, Zhang Guangtao, Li Chaolun,et al. Interannual variations of zooplankton in Zhangzi Island, North Yellow Sea[J]. Haiyang Xuebao,2017,39(8):78—88, doi∶10.3969/j.issn.0253-4193.2017.08.008
猜你喜欢
农村青少年科学探究(2023年4期)2023-07-30 10:20:06
法人(2022年5期)2022-05-23 02:30:11
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
科学与财富(2021年35期)2021-05-10 14:54:31
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
儿童故事画报(2020年8期)2020-10-30 01:58:28
党员生活·中(2019年12期)2019-08-13 07:45:54
经理人(2018年1期)2018-02-03 17:56:04
天天爱科学(2017年12期)2018-01-31 02:11:35
