
香港巨牡蛎对3种浮游植物摄食率和滤清率的研究
2017-08-10 12:17:13付家想蓝文陆李天深李琼珍范航清黄凌风
海洋学报 2017年8期
付家想,蓝文陆,李天深,李琼珍,范航清,黄凌风*
(1. 厦门大学 环境与生态学院,福建 厦门361101;2. 广西壮族自治区海洋环境监测中心站,广西 北海 536000;3.广西科学院 广西红树林研究中心 广西红树林保护与利用重点实验室,广西 北海 536000; 4. 广西水产研究院 贝类科学实验站,广西 南宁530004)
香港巨牡蛎对3种浮游植物摄食率和滤清率的研究
付家想1,蓝文陆2,李天深2,李琼珍4,范航清3,黄凌风1*
(1. 厦门大学 环境与生态学院,福建 厦门361101;2. 广西壮族自治区海洋环境监测中心站,广西 北海 536000;3.广西科学院 广西红树林研究中心 广西红树林保护与利用重点实验室,广西 北海 536000; 4. 广西水产研究院 贝类科学实验站,广西 南宁530004)
本文在室内条件下研究了大、中、小3种规格的香港巨牡蛎(Crassostreahongkongensis)对牟氏角毛藻(Chaetocerosmuelleri)、亚心形扁藻(Platymonassubcordiformis)、球等鞭藻(Isochrysisgalbana)3种浮游植物在同等密度、同等生物量条件下的摄食率和滤清率,探讨了香港巨牡蛎对3种浮游植物的摄食差异及其影响因素。结果表明,在28℃水温条件下,香港巨牡蛎的单位个体滤清率和单位体质量滤清率分别为1.40~8.94 L/(ind·h)和0.86~3.17 L/(g·h),等密度和等生物量浮游植物条件下香港巨牡蛎均表现出单位个体滤清率由高到低依次为:大规格、中规格、小规格;不同规格香港巨牡蛎的单位体质量滤清率相近。相同藻类密度条件下,香港巨牡蛎的单位体质量滤清率由高到低依次为:亚心形扁藻、球等鞭金藻、牟氏角毛藻;相同生物量条件下,香港巨牡蛎对3种浮游植物单位体质量滤清率由高到低依次为:球等鞭金藻、亚心形扁藻、牟氏角毛藻。香港巨牡蛎对不同浮游植物种类和密度的滤清率、摄食率差异主要是受饵料的大小、营养质量和密度等因素影响。
香港巨牡蛎;浮游植物;滤清率;摄食率;摄食选择性
1 引言
香港巨牡蛎(Crassostreahongkongensis)属于暖水性贝类,原称近江牡蛎,俗称“白蚝”,是我国粤西、广西沿海特有的优质食用贝,也是广东和广西的一个重要养殖品种。自2003年香港学者Lam和Morton[1]将珠江三角洲的近江牡蛎定为一个新种并命名为香港巨牡蛎以来,各界学者对其便多有研究。近年来,利用生物控制调节富营养化水平已经成为研究热点[2-6],双壳贝类能够滤食浮游生物幼体、大部分浮游植物以及有机碎屑,可以有效控制赤潮的发生[7],但大面积的牡蛎养殖因其较高的滤清率和摄食率可能显著改变周围水体浮游植物群落结构进而对海区生态系统产生较大影响。方建光等[8]依据桑沟湾和庙岛湾双壳贝类养殖前后现场调查结果,发现在大规模贝类养殖的影响下,浮游植物优势种发生改变。卢静等利用海湾扇贝在陆基虾池围隔实验中也发现放养滤食性双壳贝类能显著性改变浮游生物群落结构[9]。国内对对香港巨牡蛎的研究多集中在遗传、育种方面,少有其关于摄食、代谢生理的研究[10-13],同时从生态学角度对不同浮游植物滤清率、摄食率以及下行控藻的研究也尚不足。滤清率、摄食率是贝类的两个重要生理参数,也是研究贝类生物调控、养殖容量及其生态影响等不可或缺的重要指标。本文研究不同规格的香港巨牡蛎对不同浮游植物摄食率、滤清率,以期为香港巨牡蛎滤食能力、控藻水平以及科学评估香港巨牡蛎养殖容量、生态影响的研究提供数据依据。
2 材料方法
2.1 实验用香港巨牡蛎
香港巨牡蛎取自广西钦州市茅尾海龙门海域牡蛎筏式养殖区,分别在养殖1年龄、2年龄、3年龄的养殖筏中选取壳高6 cm、10 cm、14 cm左右代表小、中、大3种不同规格的牡蛎个体。香港巨牡蛎运回到国家贝类产业体系广西贝类综合实验站(北海)开展实验。清洗干净并去除贝壳上的附着物,放入装有80 L砂滤海水的塑料水箱中暂养1周。暂养期间持续充气,每2天换1次海水,保证牡蛎的活性;早晚定时投喂10 L浓度约3×105cells/L的小球藻(Chlorellasp.)各1次。暂养期间观察牡蛎个体的活性,实验前1天停止投饵,随机挑选活性较好的个体进行实验。暂养香港巨牡蛎以及培养藻类的海水为北海市铁山港竹林海域砂滤海水,盐度22~24,海水温度26~28℃。
2.2 实验藻类及其初始条件
实验用藻为北部湾近岸海域常见的牟氏角毛藻(Chaetocerosmuelleri)、球等鞭金藻(Isochrysisgalbana)、亚心形扁藻(Platymonassubcordiformis),分别代表硅藻、金藻和绿藻,均取自国家贝类产业体系广西贝类综合实验站(北海)藻类二级扩种培养室。实验前3种藻经过400目筛绢去除杂质,并在40倍显微镜下测定其细胞大小:每种藻选择3个视野,每个视野随机选定10个细胞测量;每种藻的大小规格及其近似球体时的直径参数如表1。
注:直径为近似球体时的参数,牟氏角毛藻参数未包括角毛。
等密度实验中3种浮游藻类的设定初始密度为5.4×104cells/mL。
3种实验浮游植物单个细胞的体积、生物量按照孙军[14]细胞体积表面积模型及转换生物量的方法计算,其中牟氏角毛藻和亚心形扁藻采用椭圆柱体模型、球等鞭金藻采用圆锥加半球复合体模型计算。等生物量实验中,3种浮游植物的设定生物量为1 μg/mL(以碳计),牟氏角毛藻、亚心形扁藻、球等鞭金藻3种浮游植物对应的密度分别为3×105cells/mL、 2.5×104cells/mL和1×105cells/mL。
2.3 摄食实验方法
摄食实验采用静水系统,在容量为5 L的塑料桶中进行,等密度和等生物量的摄食实验分开进行,采用与牡蛎暂养同样的砂滤海水(盐度24,水温28℃)在室温下32℃条件下进行。
在清洗干净的塑料桶中加入4 L 3种浮游植物藻液,轻移准备好的香港巨牡蛎放入塑料桶中,每个塑料桶放入1只香港巨牡蛎,观察记录每个塑料桶中香港巨牡蛎开口摄食时间,开口时计时并在1 h后取出牡蛎同时取10 mL藻液用鲁戈试剂固定。香港巨牡蛎按个体分为大、中、小3种不同规格,每种规格设置3个重复,另设一个无贝空白作为对照以消除饵料繁殖和自然沉降的影响。实验中用气石充气使饵料混合均匀并保证有充足的溶氧,气量以不搅动桶底排泄物为准。
将实验后固定好的藻类样品,摇匀后用移液枪取100 μL放入浮游生物计数框在显微镜下计数。测定香港巨牡蛎的壳高,随即开壳取出软体部,在烘箱(60℃)中烘干48 h,称量其干质量(精确到0.001 g)。
分别计算香港巨牡蛎的摄食百分比(FR%),单位个体滤清率(CRind)、摄食率(FRind)以及单位质量的滤清率(CRmass)、摄食率(FRmass);摄食百分比、滤清率、摄食率按以下公式进行计算:
摄食百分比FR%=(Q0-Q1)/Q0×100%,
(1)
滤清率CRmass=(Q0-Q1)/Q0×VS×(1/W)b/T,
(2)
CRind=V×(InQ0-InQ1)/N×T,
(3)
摄食率FRmass=(Q0-Q1)×VS×(1/W)b/T,
(4)
FRind=(Q0-Q1)×VS×/N×T,
(5)
式中,Q0、Q1分别为香港巨牡蛎摄食前后海水中浮游植物细胞密度(cells/L);Vs为实验用海水体积(L);W为香港巨牡蛎软体部干质量(g);N为实验牡蛎个数;T为摄食时间(h);b取0.62[15]。
2.4 数据处理
实验数据为3个平行组的平均值,以平均值±标准差(mean±S.D.)表示,用Excel 2010作图,数据差异显著性检验用SPSS19.0分析(以P<0.05为差异显著性标准)。
3 结果与分析
3.1 实验香港巨牡蛎生物学指标
实验用香港巨牡蛎的生物学特征壳高(最长部分,mm)和软体部干质量(g)参数如表2;其中小规格牡蛎平均壳高范围62.46~71.14 mm,平均软体干质量为0.940 1~1.185 9 g;中规格牡蛎平均壳高范围93.67~98.90 mm,平均软体干质量1.515 5~1.763 8 g;大规格牡蛎平均壳高范围135.90~140.55 mm,平均软体干质量2.346 4~2.643 4 g。

表2 香港巨牡蛎生物学测定
3.2 香港巨牡蛎对相同密度浮游植物的摄食
在5.4×104cells/mL藻类密度下,香港巨牡蛎摄食1 h后3种藻类的密度均明显减小,其中亚心形扁藻被摄食比例最高,摄食比例为44%~92%,球等鞭金藻的摄食比例为62%~87%,牟氏角毛藻被摄食的比例最低,只有29%~48%(表3)。整个实验期间,香港巨牡蛎对3种藻类的摄食强度均表现大规格最大,并且随着个体规格减小中等规格、小规格的香港巨牡蛎摄食强度也逐渐降低。

表3 相同密度条件下不同规格香港巨牡蛎对3种浮游植物的摄食百分比
图1和图2分别列出了等密度下香港巨牡蛎对3种藻类的清除率和摄食率。在5.4×104cells/mL藻类密度下,香港巨牡蛎对球等鞭金藻、牟氏角毛藻和亚心形扁藻的个体滤清率变化范围分别为3.87~5.17 L/(ind·h)、1.40~4.39 L/(ind·h)、3.08~8.94 L/(ind·h)。香港巨牡蛎对亚心形扁藻、球等鞭金藻的个体摄食率相近(P>0.05),对牟氏角毛藻的个体摄食率明显低于其他两种藻类(P<0.01)。香港巨牡蛎对每种藻类的个体滤清率都是大规格最高,小规格最小,随着个体大小增加而增加。对球等鞭金藻和亚心形扁藻的单位体重滤清率明显高于牟氏角毛藻(P<0.01)。
在相同藻密度下,香港巨牡蛎对亚心形扁藻(1.65±0.36)×108cells/ (ind·h)、球等鞭金藻(1.61±0.22)×108cells/ (ind·h)的单位个体摄食率相近(P>0.05),对牟氏角毛藻(8.6±4.16)×107cells/ (ind·h)的摄食率明显低于其他两种藻类(P<0.01)。对3种藻类的单位体质量摄食率分别为球等鞭金藻(1.23±0.14)×108cells/(g·h)、牟氏角毛藻(6.28±1.44)×107cells/(g·h)、亚心形扁藻(1.27±0.18)×108cells/(g·h)。香港巨牡蛎对3种浮游植物以及不同规格香港巨牡蛎的单位个体和单位体质量摄食率变化与滤清率相似。
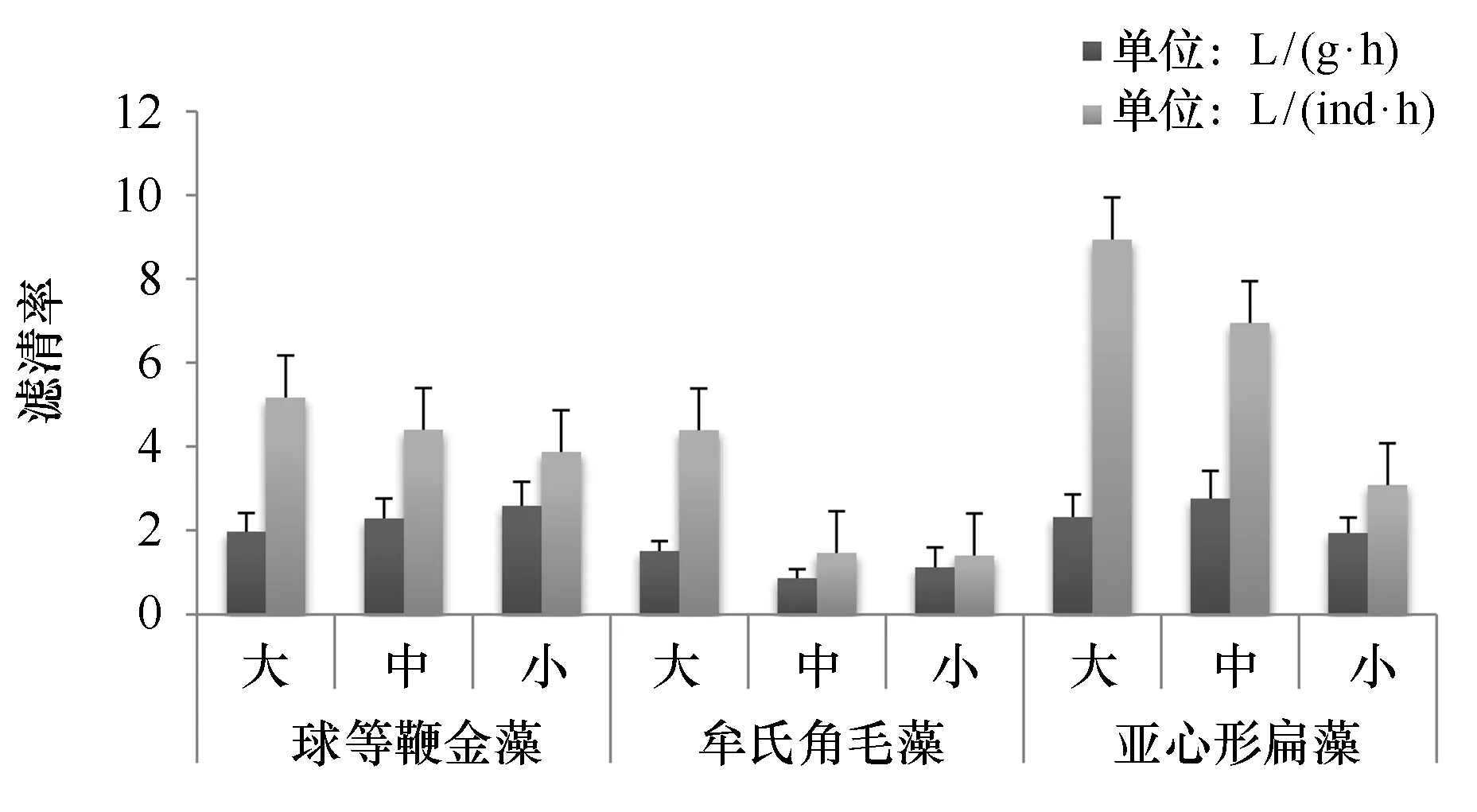
图1 不同规格香港巨牡蛎在相同藻密度条件下对3种浮游植物的滤清率Fig.1 Clearance rate of three different phytoplankton by different size of C. hongkongensis in same algae density
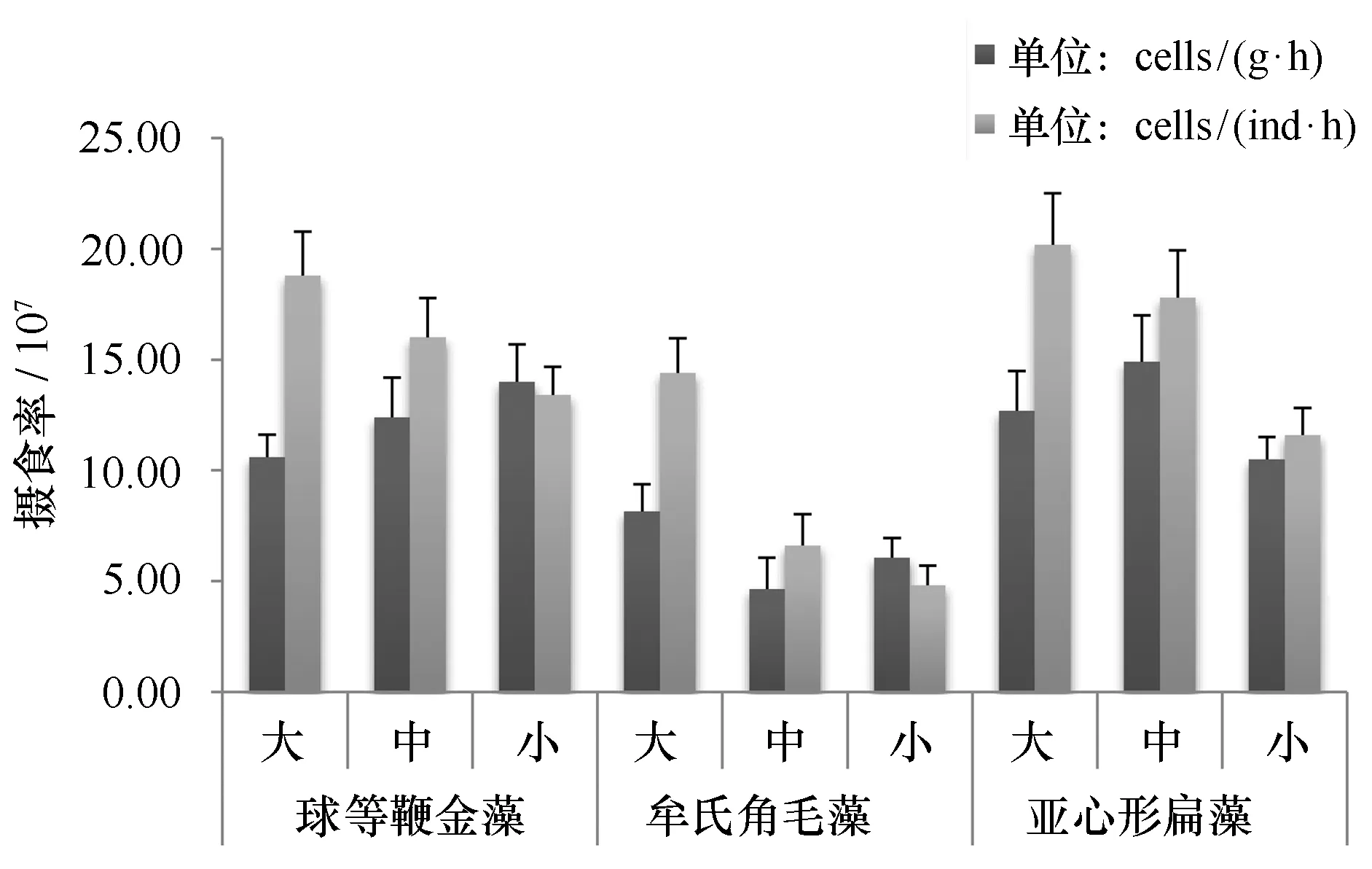
图2 不同规格香港巨牡蛎在相同藻密度条件下对3种浮游植物的摄食率Fig.2 Filtration rate of three different phytoplankton by different size of C. hongkongensis in same algae density
3.3 对相同生物量浮游植物的摄食
在1 μg/mL(以碳计)等生物量条件下,香港巨牡蛎对3种藻类摄食明显,对球等鞭金藻、牟氏角毛藻、亚心形扁藻的摄食比例范围分别为81%~95%、60%~93%、46%~84%;球等鞭金藻被摄食的比例最高,亚心形扁藻最低(表4)。与等密度条件下的表现相似,香港巨牡蛎规格越大摄食强度越大。

表4 相同生物量条件下不同规格香港巨牡蛎对3种浮游植物的摄食百分比
如图3,等生物量条件下,香港巨牡蛎对3种藻的个体滤清率变化范围分别为球等鞭金藻5.71~8.4 L/(ind·h)、牟氏角毛藻2.11~5.49 L/(ind·h)、亚心形扁藻2.43~7.3 L/(ind·h)。与等密度条件下的滤清率相似,3种规格香港巨牡蛎对每种藻类的单位个体滤清率都表现出规格越大滤清率越高(P<0.01),而且同种规格的香港巨牡蛎对3种浮游植物的个体滤清率由高到低依次为:球等鞭金藻、亚心形扁藻、牟氏角毛藻(P<0.05)。对3种浮游植物单位体质量滤清率变现为:球等鞭金藻(2.73±0.42)L/(g·h)、牟氏角毛藻(2.02±0.29)L/(g·h)、亚心形扁藻(1.68±0.12)L/(g·h)。香港巨牡蛎对3种浮游植物单位体质量滤清率差异不明显(P>0.05)。3种不同规格的香港巨牡蛎对同种藻的单位体质量滤清率差异性也不显著(P>0.05),但同种规格的香港巨牡蛎对3种浮游植物的单位体质量滤清率由高到低依次为:球等鞭金藻、亚心形扁藻、牟氏角毛藻。
香港巨牡蛎对3种浮游植物的单位个体摄食率分别为亚心形扁藻(6.92±1.7)107cells/(ind·h)、等鞭金藻(3.51±0.34)×108cells/(ind·h)、牟氏角毛藻(7.24±1.7)×108cells/(ind·h)。香港巨牡蛎对3种浮游植物的单位个体摄食率有显著性差异(P<0.01),由高到低依次为:牟氏角毛藻、球等鞭金藻、亚心形扁藻。香港巨牡蛎单位体质量摄食率变化与滤清率相似(图4)。
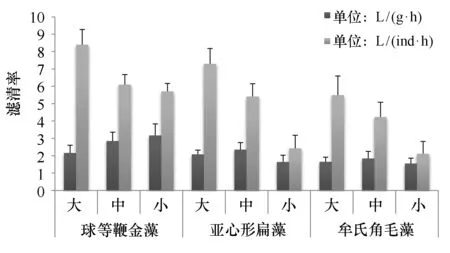
图3 不同规格香港巨牡蛎在相同藻生物量条件下对3种浮游植物的滤清率Fig.3 Clearance rate of three different phytoplankton by different size of C. hongkongensis in same algae biomass
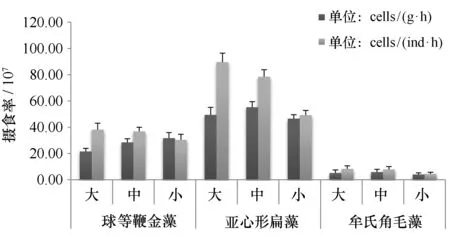
图4 不同规格香港巨牡蛎在相同藻生物量条件下对3种浮游植物的摄食率Fig.4 Filtration rate of three different phytoplankton by different size of C. hongkongensis in same algae biomass
4 讨论
4.1 香港巨牡蛎滤清率与其他巨牡蛎的比较
香港巨牡蛎是我国亚热带沿海地区的一个重要贝类养殖品种,在广东、广西等近岸滩涂和沿岸海域大面积养殖。鉴于目前对香港巨牡蛎不同规格以及对不同浮游植物摄食的系统性研究尚缺乏,本实验在室内通过在3种不同藻类饵料、不同饵料密度和不同饵料生物量的条件下对3种不同规格的香港巨牡蛎的滤清率和摄食率进行了研究。在本实验条件下得出香港巨牡蛎的单位体质量滤水滤为0.86~3.17 L/(g·h),香港巨牡蛎滤清率的结果与国内报道的其他巨牡蛎相比(表5),处在一般正常水平;香港巨牡蛎滤清率与太平洋牡蛎相近,略高于高露姣等[16]报道的巨牡蛎滤清率结果,但低于林丽华等[17]、廖文崇等[18]对香港巨牡蛎研究报道的滤水率,这种差异可能与不同种类之间的种类大小、生理习性不同有着重要关系,也可能与同种类不同个体之间的大小差异有关;另外贝类的摄食还受到其他环境因子的直接影响,如温度、盐度、溶解氧、pH等。此实验是在水温为28℃条件下进行的,有研究[17]表明香港巨牡蛎滤清率在水温为22℃时有最大滤水率,因此较高水温很可能是本实验中香港巨牡蛎滤水率表现偏低的原因。

表5 几种大型巨牡蛎滤清率对比
4.2 不同规格香港巨牡蛎的摄食率和滤清率变化
本实验结果表明香港巨牡蛎个体的大小对单位个体滤清率和摄食率有着显著的影响,随着香港巨牡蛎个体大小的增加个体滤清率和摄食率明显增加(表3,表4,图1至图4)。滤食性双壳贝类规格大小是影响其滤清率、摄食率吸收效率的重要影响因素[18]。由香港巨牡蛎滤清率、摄食率等研究中获取的香港巨牡蛎的壳高与个体滤清率的数据表明二者之间有着较好的相关性。这种相关性为评估现场局部海湾或整个海域香港巨牡蛎对浮游植物的摄食压力以及香港巨牡蛎养殖的容量提供了非常便利的条件。香港巨牡蛎的壳高、干质量等生物参数的测定及获取方便快捷,因此在养殖容量模型和其他相关生态模型中可以加重考虑牡蛎壳高这个简单易测量获取的参数,通过牡蛎规格、产量统计和海区浮游植物丰度研究评估海区香港巨牡蛎的整体滤清率、摄食率及养殖容量。
同种浮游植物饵料条件下大、中、小不同规格香港巨牡蛎单位体质量滤清率和摄食率变化不大,并没有呈现出统一的变化规律(图1至图4);需要在今后的工作中相应地加强有关研究,以获取较为准确的不同规格单位体质量摄食率变化规律为进一步养殖容量和生态影响研究提供科学数据。
4.3 香港巨牡蛎对不同浮游植物滤清率和摄食率的差异
摄食不同的浮游植物时,香港巨牡蛎单位体质量的滤清率与个体大小之间的相互关系存在较大差异;如在摄食等鞭金藻的实验中,香港巨牡蛎的单位体质量滤水率和摄食率与香港巨牡蛎壳高/干质量大小呈负相关关系(图1至图4),这与扇贝等其他滤食性贝类的研究得到的规律一致[22-24],该现象与较小贝类个体具有较高的新陈代谢等生理活性有关。另外亚心形扁藻组小规格牡蛎的单位体质量滤清率、摄食率均小于相应中、大规格的滤水率、摄食率,牟氏角毛藻组小规格香港巨牡蛎的单位体质量滤水率、摄食率略低于大规格(图1至图4),这种现象与王芳等[21]对海湾扇贝和太平洋牡蛎滤水率的研究相似。浮游植物是双壳贝类的主要饵料,不同浮游植物细胞有机质一般也不相同,主要和浮游植物藻体大小规格有关[14]。此实验中的3种浮游植物,牟氏角毛藻细胞最小,但加上角毛后明显大于球等鞭金藻和亚心形扁藻,考虑角毛球等鞭金藻应是3种藻类中细胞最小的种类,个体小的饵料容易被小规格的香港巨牡蛎摄食而容易会被大个体贝类漏掉[25-26],加上小规格较高的生理效率导致其单位体质量滤清率和摄食率均呈现出随着个体大小减少而增加的趋势,而牟氏角毛藻和亚心形扁藻因个体较大不容易被较小的香港巨牡蛎摄食导致不一致的变化特征。因此香港巨牡蛎的滤清率和摄食率变化受浮游植物的细胞大小的控制。
双壳贝类对不同藻类的滤食具有一定的选择性,不同的贝类对不同藻类的摄食也存在差异[27]。本研究的实验结果也发现同等条件下香港巨牡蛎对不同种类的浮游植物清除率和摄食率存在较大的差异。同饵料密度条件下香港巨牡蛎对亚心形扁藻的滤清率最高,其次为球等鞭金藻,牟氏角毛藻最低(图1);等饵料生物量条件下,香港巨牡蛎对亚心形扁藻滤清率最高,其次球等鞭金藻,牟氏角毛藻最低(图3),表明香港巨牡蛎对牟氏角毛藻的喜好程度低于其他两种藻,但3种浮游植物都没有被香港巨牡蛎严重避食。出现这种摄食选择性现象主要是因为不同浮游植物饵料质量的差异。等密度条件下亚心形扁藻细胞的生物量分别是牟氏角毛藻(不含角毛)和球等鞭金藻的12.4倍和4.7倍,饵料质量由高到低依次为:亚心形扁藻、球等鞭金藻、牟氏角毛藻,因此在食物浓度一致的情况下,香港巨牡蛎更倾向摄食质量高的饵料。另外牟氏角毛藻具较长的角毛(通常为藻体的5~6倍),贝类在摄食时这些角毛上的倒刺可能会损害其腮丝和纤毛造成其对贝类适口性较低。再次牟氏角毛藻细胞壁高度硅质化,壳体坚硬以及角毛使得营养价值相对其他两种浮游植物更低。因此浮游植物的生物量及营养价值导致了在相同密度条件下香港巨牡蛎对不同浮游植物的滤清率和摄食率存在着明显差异。相类似,在相同生物量条件下,香港巨牡蛎对3种不同浮游植物的滤清率差异明显降低,相同条件下无细胞壁、裸露的球等鞭金藻具有更高营养价值,且相对于较大活动能力的亚心形扁藻更容易摄食,因此对球等鞭金藻的滤清率高于亚心形扁藻。相同生物量条件下香港巨牡蛎对不同浮游植物种类的摄食率结果差异较大主要是因为饵料密度差异的原因,同时也表明了香港巨牡蛎的这种摄食选择性具有相对性,对高密度的藻类摄食率高,通过自身调节有效避免因有机物含量过低而造成能量摄入不足[28]。
[1] Lam K, Morton B. Mitochondrial DNA and morphological identification of a new species ofCrassostrea(Bivalvia: Ostreidae) cultured for centuries in the Pearl River Delta, Hong Kong, China[J]. Aquaculture, 2003, 228(1/4): 1-13.
[2] 张绍浩. 富营养化湖泊藻类控制技术比较及新方法的研究[D]. 武汉: 华中科技大学, 2006.
Zhang Shaohao. A comparative study and new methods on control algae in eutrophic lakes[D]. Wuhan: Huazhong University of Science & Technology, 2006.
[3] 董文龙, 闵水发, 唐利军, 等. 洪湖、梁子湖水体富营养化研究[J]. 环境科学, 2016, 42(4): 66-70.
Dong Wenlong, Min Shuifa, Tang Lijun, et al. Research of eutrophication of Honghu Lake and Liangzi Lake[J]. Environmental Protection Science, 2016, 42(4): 66-70.
[4] 陈进斌, 苗英霞, 邱金泉, 等. 海水养殖废水处理技术研究进展[J]. 盐业与化工, 2016, 45(5): 1-5.
Chen Jinbin, Miao Yingxia, Qiu Jinquan, et al. Development of marine aquaculture wastewater treatment technology[J]. Journal of Salt and Chemical Industry, 2016, 45(5): 1-5.
[5] 过峰, 赵俊, 陈聚法, 等. 胶州湾贝类养殖区氮、磷污染现状及动态变化[J]. 渔业科学进展, 2012, 33(5): 116-122.
Guo Feng, Zhao Jun, Chen Jufa, et al. Nitrogen and phosphorous pollution in shellfish culture areas of Jiaozhou Bay[J]. Progress in Fishery Science, 2012, 33(5): 116-122.
[6] 黄通谋, 李春强, 于晓玲, 等. 麒麟菜与贝类混养体系净化富营养化海水的研究[J]. 中国农学通报, 2010, 26(18): 419-424.
Huang Tongmou, Li Chunqiang, Yu Xiaoling, et al. Purification of chlorophyll A, nitrogen and phosphate in eutrophic water byPaphiaexarataphilippi and kappaphycus striatum schmitz[j]. Chinese Agricultural Science Bulletin, 2010, 26(18): 419-424.
[7] Prins T C, Escaravage V, Smaal A C, et al. Nutrient cycling and phytoplankton dynamics in relation to mussel grazing in a mesocosm experiment[J]. Ophelia, 1995, 41(1): 289-315.
[8] 方建光, 匡世焕, 孙慧玲, 等. 桑沟湾栉孔扇贝养殖容量的研究[J]. 海洋水产研究, 1996, 17(2): 18-31.
Fang Jianguang, Kuang Shihuan, Sun Huiling, et al. Study on the carrying capacity of Sanggou Bay for the culture of scallopChlamysfarreri[J]. Marine Fisheries Research, 1996, 17(2): 18-31.
[9] 卢静, 李德尚, 董双林. 对虾池混养滤食性动物对浮游生物的影响[J]. 青岛海洋大学学报, 1999, 29(2): 243-248.
Lu Jing, Li Deshang, Dong Shuanglin. The impact of the polycultured filter-feeding animals with peneid shrimp on plankton community[J]. Journal of Ocean University of Qingdao, 1999, 29(2): 243-248.
[10] 潘英, 夏朝林, 谢卫著. 人工诱导香港巨牡蛎雌核发育单倍体的研究[J]. 热带海洋学报, 2008, 27(3): 55-60.
Pan Ying, Xia Chaolin, Xie Weizhu. Studies on artificial induction of haploid gynogenesis in oysterCrassostreahongkongensis[J]. Journal of Tropical Oceanography, 2008, 27(3): 55-60.
[11] 李咏梅, 陈秀荔, 彭敏, 等. 基于线粒体COI基因序列探讨广西钦州湾牡蛎的遗传分化[J]. 西北农林科技大学学报(自然科学版), 2009, 37(3): 60-65.
Li Yongmei, Chen Xiuli, Peng Min, et al. Genetic differentiation of oyster from Qinzhou Bay based on mitochondrial cytochrome oxidase subunit I gene[J]. Journal of Northwest A&F University (Natural Scienice Edition), 2009, 37(3): 60-65.
[12] 孙泽伟, 郑怀平, 杨彦鸿, 等. 近江牡蛎养殖群体数量性状间的相关及通径分析[J]. 中国农学通报, 2010, 26(6): 332-336.
Sun Zewei, Zheng Huaiping, Yang Yanhong, et al. Correlation and path analysis to quantitative traits for a cultured population of JinjiangOysterCrassostreahongkongensis[J]. Chinese Agricultural Science Bulletin, 2010, 26(6): 332-336.
[13] 霍忠明, 王昭萍, 梁健, 等. 香港巨牡蛎与近江牡蛎杂交及回交子代早期生长发育比较[J]. 水产学报, 2013, 37(8): 1155-1161.
Huo Zhongming, Wang Zhaoping, Liang Jian, et al. Comparisons of growth and development of hybrid and backcrossing progenies betweenCrassostreahongkongensisandCrassostreaariakensis[J]. Journal of Fisheries of China, 2013, 37(8): 1155-1161.
[14] 孙军. 海洋浮游植物细胞体积和表面积模型及其转换生物量[D]. 青岛: 中国海洋大学, 2004.
Sun Jun. Geometric models for calculating cell biovolume and surface area for marine phytoplankton and its relative conversion biomass[D]. Qingdao: Ocean University of China Qingdao, 2004.
[15] Riisgård H U. Filtration rate and growth in the blue mussel,Mytilus edulis Linnesus,1785:dependence on algal concentration[J]. Journal of Shellfish Research,1991,10:29-35.
[16] 高露姣, 沈盎绿, 陈亚瞿, 等. 巨牡蛎(Crassostreasp.)的滤水率测定[J]. 海洋环境科学, 2006, 25(4): 62-65.
Gao Lujiao, Shen Anglv, Chen Yaqu, et al. Determination of filtration rate ofCrassostreasp.[J]. Marine Environmental Science, 2006, 25(4): 62-65.
[17] 林丽华, 廖文崇, 谢健文, 等. 盐度对香港巨牡蛎摄食和代谢的影响[J]. 广东农业科学, 2012, 39(11): 10-14.
Lin Lihua, Liao Wenchong, Xie Jianwen, et al. Effect of salinity on the feeding and metabolic physiology ofCrassostreahongkongensis[J]. Guangdong Agricultural Sciences, 2012, 39(11): 10-14.
[18] 廖文崇, 朱长波, 张汉华. 体规格对香港巨牡蛎摄食和代谢的影响[J]. 中国渔业质量和标准, 2011, 1(3): 41-46.
Liao Wenchong, Zhu Changbo, Zhang Hanhua. Effect sizes on the feeding and metabolic physiology ofCrassostreahongkongensis[J]. Chinese Fishery Quality and Standards, 2011, 1(3): 41-46.
[19] 王吉桥, 于晓明, 郝玉冰, 等. 4种滤食性贝类滤水率的测定[J]. 水产科学, 2006, 25(5): 217-221.
Wang Jiqiao, Yu Xiaoming, Hao Yubing, et al. Determination of clearance rate in four filtering mussel species[J]. Fisheries Sicience, 2006, 25(5): 217-221.
[20] 王俊,蒋祖辉,张波,等. 太平洋牡蛎同化率的研究[J]. 应用生态学报,2000,11(3):441-444.
Wang Jun,Jiang Zuhui,Zhang Bo,et al. Assimilation efficiency of pacific oyster Crassortrea gigas[J]. Chinese Journal of Applied Ecology,2000,11(3):441-444.
[21] 王芳, 董双林, 张硕, 等. 海湾扇贝和太平洋牡蛎的食物选择性及滤除率的实验研究[J]. 海洋与湖沼, 2000, 31(2): 139-144.
Wang Fang, Dong Shuanglin, Zhang Shuo, et al. Experimental studies on feeding selectivity and the filter-feeding rate ofArgopectenirradiansandCrassostreagigas[J]. Oceanologia et Limnologia Sinica, 2000, 31(2): 139-144.
[22] 周毅, 毛玉泽, 杨红生, 等. 四十里湾栉孔扇贝清滤率、摄食率和吸收效率的现场研究[J]. 生态学学报, 2002, 22(9): 1455-1462.
Zhou Yi, Mao Yuze, Yang Hongsheng, et al. Clearance rate, ingestion rate and absorption efficiency of the scallopChlamysfarrerimeasured byinsitubiodeposition method[J]. Acta Ecologica Sinica, 2002, 22(9): 1455-1462.
[23] 张继红. 滤食性贝类养殖活动对海域生态系统的影响及生态容量评估[D]. 青岛: 中国科学院海洋研究所, 2008.
Zhang Jihong. Effect on filter feeding shellfish mariculture on the ecosystem and the evaluation of ecology carrying capacity[D]. Qingdao: The Institute of Oceanology, Chinese Academy of Sciences, 2008.
[24] 张继红, 方建光, 孙松, 等. 胶州湾养殖菲律宾蛤仔的清滤率、摄食率、吸收效率的研究[J]. 海洋与湖沼, 2005, 36(6): 548-555.
Zhang Jihong, Fang Jianguang, Sun Song, et al. Clearance rate, ingestion rate and absorption efficiency of cultivated clam ruditapes philippinarum in Jiaozhou Bay, China[J]. Oceanologia et Limnologia Sinica, 2005, 36(6): 548-555.
[25] 王如才, 王昭平, 张建中. 海水贝类养殖学[M]. 青岛: 青岛海洋大学出版社, 1993: 40.
Wang Rucai, Wang Zhaoping, Zhang Jianzhong. Science of Marine Shellfish Culture[M]. Qingdao: Ocean University of China Press, 1993: 40.
[26] 张莉红. 扇贝养殖对浮游植物群落影响的初步研究[D]. 青岛: 中国海洋大学, 2005.
Zhang Lihong. A preliminary study of the impacts of scallop culture on phytoplankton community[D]. Qingdao: Ocean University of China, 2005.
[27] 张爱菊, 朱俊杰, 刘金殿. 3种微藻对池蝶蚌幼蚌的选择滤食与生长的影响[J]. 浙江海洋学院学报(自然科学版), 2012, 31(1): 65-69.
Zhang Aiju, Zhu Junjie, Liu Jindian. Effects of three species of microalgae on ingestion rate and growth of juvenileHyriopsisschlegeli[J]. Journal of Zhejiang Ocean University (Natural Science), 2012, 31(1): 65-69.
[28] Wong W H, Cheung S G. Feeding behaviour of the green mussel,Pernaviridis(L.): responses to variation in seston quantity and quality[J]. Journal of Experimental Marine Biology and Ecology, 1999, 236(2): 191-207.
Research of clearance rate and filtration rate ofCrassostreahongkongensisfeeding on three different phytoplankton
Fu Jiaxiang1, Lan Wenlu2, Li Tianshen2, Li Qiongzhen4, Fan Hangqing3, Huang Lingfeng1
(1.CollegeoftheEnvironmentandEcologicalScience,XiamenUniversity,Xiamen361001,China; 2.MarineEnvironmentalMonitoringCenterofGuangxi,Beihai536000,China; 3.GuangxiMangroveResearchCenter,GuangxiAcademyofScience,GuangxiKeyLaboratoryofMangroveProtection,Beihai536007,China;4.GuangxiAcademyofFisheryScience,Nanning530004,China)
The clearance rate and filtration rate ofCrassostreahongkongensisfeeding onChaetocerosmuelleri,IsochrysisgalbanaandPlatymonassubcordiformiswere examined using the lab experiments under the conditions of same density and biomass of phytoplankton by three different size ofC.hongkongensisto discuss the feeding preference and impact infactors on phytoplankton ofC.hongkongensis.The results showed that at temperature 28℃, the clearance rate per gram and per individual were 0.86-3.17 L/(g·h) and 1.40-8.94 L/(ind·h) respectively. The clearance rate per individual ofC.hongkongensispresented as big-size> medium-size> small-size in the both case of same density and biomass of three phytoplankton, while the clearance rate per garm ofC.hongkongensiswas similar in spite of the size ofC.hongkongensis. The changing pattern of clearance rate per individual presented asPlatymonassubcordiformis>Isochrysisgalbana>Chaetocerosmuelleriunder the condition of same density of three phytoplankton, while presented asIsochrysisgalbana>Platymonassubcordiformis>Chaetocerosmuelleriunder the condition of same biomass of three phytoplankton. The different clearance rate ofC.hongkongensisonto different phytoplanton was mainly due to factors such as the food particle size, density and nutritive value.
Crassostreahongkongensis; phytoplankton; clearance rate; filtration rate; feeding selectivity
10.3969/j.issn.0253-4193.2017.08.006
2017-01-10;
2017-03-20。
国家自然科学基金项目(41466001);广西科学研究与技术开发计划项目(桂科攻1598016-6,桂科AA17129001,桂科AD17129041);广西自然科学基金项目(2015GXNSFBA139194;2015GXNSFAA139244);广西红树林保护与利用重点实验室开放基金项目(GKLMC-200504)。
付家想(1989—),男,河南省信阳市人,主要研究方向为海洋生态学。E-mail:1534759513@qq.com
*通信作者:黄凌风,教授,博士生导师。E-mail:huanglf@xmu.edu.cn
S917.4
A
0253-4193(2017)08-0062-08
付家想,蓝文陆,李天深,等. 香港巨牡蛎对3种浮游植物摄食率和滤清率的研究[J].海洋学报,2017,39(8):62—69,
Fu Jiaxiang, Lan Wenlu, Li Tianshen, et al. Research of clearance rate and filtration rate ofCrassostreahongkongensisfeeding on three different phytoplankton[J]. Haiyang Xuebao,2017,39(8):62—69, doi:10.3969/j.issn.0253-4193.2017.08.006
猜你喜欢
海洋通报(2022年5期)2022-11-30 12:04:10
当代水产(2021年1期)2021-03-19 05:17:14
小资CHIC!ELEGANCE(2020年28期)2020-09-17 13:27:22
理化检验-化学分册(2020年5期)2020-06-15 11:36:14
食品与生活(2019年8期)2019-10-30 12:13:09
当代水产(2019年9期)2019-10-08 08:02:44
甘肃畜牧兽医(2019年2期)2019-03-20 01:49:42
意林(儿童绘本)(2019年2期)2019-03-12 01:01:26
小学生优秀作文(低年级)(2018年6期)2018-05-19 01:54:39
阅读与作文(小学高年级版)(2014年12期)2015-02-04 06:44:59
