
不同光周期条件下金花菜F2代农艺性状的变异特征
2017-07-30 11:29:20周克友李争艳任海龙魏臻武
草地学报 2017年6期
周克友, 李争艳, 陈 祥, 任海龙,2, 江 舟, 魏臻武*
(1. 扬州大学动物科学与技术学院/扬州大学草业科学研究所, 江苏 扬州 225009; 2. 新疆农业科学院海南三亚农作物育种试验中心, 海南 三亚 572014; 3.安徽省各科院畜牧兽医研究所, 安徽, 230031)
金花菜(MedicagopolymorphaL.)是豆科苜蓿属一年生草本植物[1],俗称南苜蓿、秧草、草头等,在长江中下游地区常作为牧草、绿肥和蔬菜等栽培利用[2]。金花菜在我国南方草牧业发展和生态环境建设方面具有重要的应用价值,其种质资源的鉴定、保存和利用正日益受到人们的重视。金花菜具有良好的再生能力,大田种植时能刈割4~6茬[3],并且马伯艳等[4]研究表明不同刈割茬数的金花菜营养成分并没有明显变化。因此,如果能推迟金花菜开花时期,延长其生长期,就能有效增加其刈割次数,提高利用效益。而日照条件是影响植物花期的关键环境因子之一,植物的花期会随植物日照条件的改变而发生变化,同时,植物的性状也会有所变化。
在豆科植物中,大豆(GlycinemaxL.)经常作为研究光周期的模式材料[5]。大豆是典型的短日照植物,光周期是影响大豆产量和品质的重要生态因子[6]。韩天富等[7]通过研究不同生态类型的大豆开花前后的农艺性状及其光周期敏感性反应,确定了大豆生育期结构与产量性状的关系。王英等[6]以开花期相近的181个大豆重组自交系(RIL)为材料,研究了开花后不同光照长度对大豆主要农艺性状的影响,指出开花后光照处理对大豆农艺性状和品质性状有较大影响。豆科苜蓿属植物的光周期研究大多是探讨不同光周期下光敏色素、植物生长激素对苜蓿(MedicagosativaL.)秋眠性的影响[8-9]。研究金花菜农艺性状和光周期关系的报道较少。Del Pozo[10]搜集了9份不同纬度地区金花菜种质材料,利用温室和田间试验研究了光周期对金花菜开花的影响,认为不同来源地的金花菜有着相应的光周期敏感性,湿润地区的金花菜开花较晚,干旱地区的金花菜开花较早。陈祥等[11]通过人工气候室模拟不同光周期条件,研究了光周期与种质特性对金花菜初花期的影响,认为金花菜是长日照植物,确定光周期是影响金花菜初花期的主要环境因素之一。
本研究选取了2个性状差异明显的金花菜种质材料,并利用人工气候室模拟10 h短日照和16 h长日照2种光周期条件,在不同光周期下进行金花菜的开花诱导试验,研究了金花菜F2代株高、叶长、叶宽和分枝数的变异特征,旨在确定金花菜的日照植物类型,并为利用光周期调控金花菜花期、选育优良金花菜品种等方面提供依据。
1 材料与方法
1.1 供试材料
本试验以陈祥等[11]研究中光周期敏感度的量化指标LD-FHR(长日开花促进率)以及性状特征达到显著差异(P< 0.05)的2个金花菜材料江苏海安(花期晚,光照13 h)和浙江温岭(花期早,光照16 h)作为亲本(表1)。试验材料于2013年分别从江苏海安、浙江温岭搜集而来,经过一代自交繁殖,以保证种子纯度。采用人工授粉进行杂交,通过杂交获得金花菜F1代种子,F1代在人工气候室内种植获得F2代。

表1 供试金花菜的来源、形态特征及相关信息Table1 Information of the tested Medicago polymorpha
1.2 试验设计与管理
本试验于2015年10月在扬州大学扬子津校区人工气候室内进行。首先挑出形态饱满、有活力的金花菜P1、P2种子各60粒及F2代种子300粒,用95%的酒精浸泡消毒1 min,然后用蒸馏水清洗3次。再将消毒后的种子划破种皮,均匀铺于含有湿润滤纸的培养皿中,用封口膜密封,放到25℃培养箱中催芽。等到种子长出胚芽后取出,移栽于花盆中,每盆1株。试验用土是肥力中等偏上的沙壤土,在装盆前已搅拌均匀。花盆放在自制的水槽之中,土壤可从花盆底部孔眼吸水至饱和,保证每个花盆中的土壤含水量相同。10 h短日照处理、16 h长日照处理的人工气候室内都移栽了长势一致的金花菜幼苗P120株、P220株以及F2代125株。人工气候室内部温度23℃、湿度50%、光强10 000 lx。
1.3 测定指标与方法
本试验观测株高、叶长、叶宽和分枝数4个主要农艺性状。取金花菜植株中间段单独复叶的中间叶片测量叶长和叶宽,选3个复叶作3次重复。分枝数测定以花盆内土表以上金花菜的分枝数为准。16 h长日照条件下金花菜F2群体的叶长、叶宽和分枝数在生长48 d后测得。10 h短日照条件下金花菜F2群体的叶长、叶宽和分枝数在生长115 d后测得。在金花菜开花初期每日定时观察,长出第一朵小花时测得绝对株高。
1.4 数据分析
利用SPSS 17.0 计算不同光周期条件下金花菜各世代农艺性状的平均值、标准差和变异系数等基本参数并进行单样本的K-S测验以及独立样本的t测验,利用EXCEL绘制各性状的次数分布图。
2 结果与分析
2.1 不同光周期条件下金花菜各世代农艺性状的变异特征
10 h短日照条件下金花菜亲本株高、叶长及叶宽均高于16 h长日照条件,且株高的差异最明显(表2)。在两种不同日照条件下,金花菜F1及F2代的株高、叶长和叶宽与亲本有相似的差异。除母本P2无论在短日照还是长日照条件下都保持1个分枝外,P1、F1和F2代在10 h短日照条件下的分枝数高于16 h长日照条件。
10 h短日照条件下金花菜F2代株高、叶长、叶宽和分枝数的变异系数均大于16 h长日照条件(表2),说明10 h短日照条件下金花菜F2代各性状有更大的变异,表现出更丰富的遗传多样性。同时,无论在何种日照条件下,金花菜F2代分枝数的变异系数都远大于其株高、叶长和叶宽的变异系数(表2),表明金花菜F2代分枝数的变异更为突出。

表2 不同光周期条件下各世代性状数据统计Table 2 The character statistics of all generations under different photoperiods
2.2 不同光周期条件下金花菜F2代性状单样本的K-S测验
单样本K-S测验不仅可以用于测验间断性变量的分布形态,也可以用于测验连续性变量的分布形态。由表3可知,在不同日照条件下,金花菜F2代株高、叶长和叶宽对应的两尾测验概率[Asymp.Sig.(2-tailed)]值均大于0.05,表明在不同日照条件下,金花菜F2代株高、叶长和叶宽都服从正态分布。而金花菜F2代分枝数在不同日照条件下两尾测验概率都小于0.05,表明分枝数不服从正态分布。
2.3 不同光周期条件下金花菜F2代同一性状独立样本的t测验
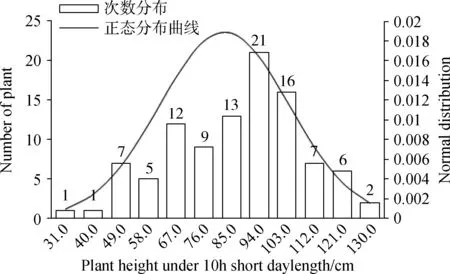
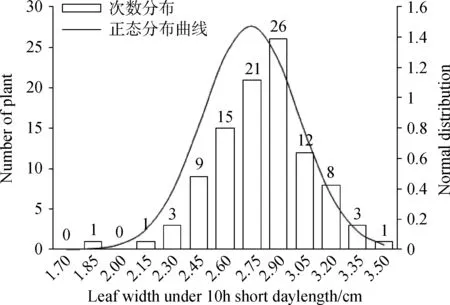
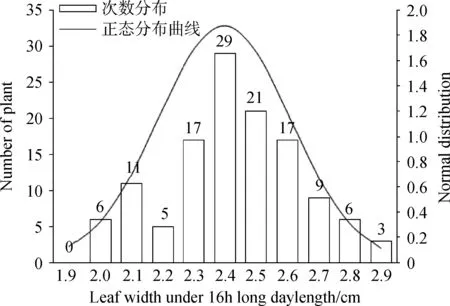
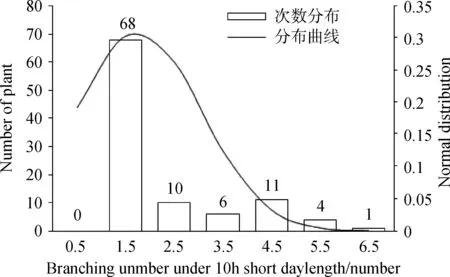
通过对不同光周期条件下金花菜F2代株高进行方差齐性检验,得到F= 80.134,P= 0.000 < 0.01(表4),因此不同光周期条件下金花菜F2代株高的方差差异极显著(P<0.01),说明方差非齐性。由于不同光周期条件下株高方差非齐性,故在进行t测验的时候应该选择“假设方差不相等”一行的结果:t=-16.710,df(自由度) = 125.230,两尾测验的P=0.000<0.01,所以认为不同光周期条件下F2代株高具有极显著差异(P<0.01)。同样,不同光周期条件下金花菜F2代叶长和叶宽的方差非齐性,选择“假设方差不相等”一行后两尾测验的P=0.000<0.01,所以不同光周期条件下金花菜 F2代叶长和叶宽也具有极显著差异(P<0.01)。虽然不同光周期条件下金花菜F2代分枝数的方差也是非齐性,但在选择“假设方差不相等”一行后两尾测验的0.01 表3 不同光周期条件下金花菜F2代性状单样本的K-S测验Table 3 The Kolmogorov-Smirnov test of different characters of F2 generation under different photoperiods 注:a表示检验分布为正态分布 Note:a indicates normal test distribution 表4 不同光周期条件下金花菜F2代同一性状独立样本的t测验Table 4 The Independent-samples t Test of the same character of F2 generation under different photoperiods 2.4.1不同光周期条件下金花菜F2代株高的次数分布 10 h短日照条件下,金花菜F2代株高(82.68cm)高于亲本P1(80.89 cm)而低于亲本P2(97. 37 cm)(表2)。金花菜F2代株高主要集中在67.0~103.0 cm,出现71次,占总数的71%(图1)。10 h短日照条件下,金花菜 F2代株高服从正态分布(表3),偏度为 - 0.36 < 0,峰度为 - 0.50,数据向右靠拢,但偏斜角度较小,表明10 h短日照条件下,金花菜F2代株高较亲本P1有增加的趋势;16 h长日照条件下,金花菜F2代株高(45.15 cm)高于亲本 P1(34.49 cm)而低于亲本P2(45.64 cm)(表2)。金花菜F2代株高集中在40.0~56.0 cm,出现91次,占总数的73.39%(图2)。16 h长日照条件下,金花菜F2代株高服从正态分布,偏度为 - 0.16 < 0,峰度为 - 0.26,其数据向右靠拢,但偏斜角度较小,表明16 h长日照条件下,金花菜F2代株高较亲本P1有增加的趋势,且倾向于亲本P2。 图1 10 h短日照条件下F2 代初花期株高频数分布Fig.1 Frequency distribution of plant height at early-flowering time of F2 generation in 10 h short day 2.4.2不同光周期条件下金花菜F2代叶长的次数分布 10 h短日照条件下,F2代叶长(2.81 cm)高于亲本P1(2.73 cm)而低于亲本P2(3.12 cm)。金花菜F2代叶长集中在2.6~3.4 cm,出现88次,占总数的88 %。10 h短日照条件下,金花菜F2代叶长服从正态分布(表3),偏度 - 1.23 < 0,峰度2.34,其数据向右靠拢,有较为明显的尖峰,数据集中(图3),表明10 h短日照条件下,金花菜F2代叶长较亲本P1有增加的趋势;16 h长日照条件下,金花菜F2代叶长(2.62 cm)高于亲本P1(2.16 cm)而小于亲本P2(2.83 cm)。金花菜F2代叶长集中在2.5~2.9 cm,出现87次,占总数的70.16 %。16 h长日照条件下,金花菜F2代叶长服从正态分布,偏度 - 0.32 < 0,峰度 - 0.58,数据向右靠拢,但偏斜程度小(图4),表明16 h长日照条件下,金花菜F2代叶长较亲本P1有增加的趋势,且倾向于亲本P2。 图2 16 h长日照条件下F2 代初花期株高频数分布Fig.2 Frequency distribution of plant height at early-flowering time of F2 generation in 16 h long day注:图中纵轴线P表示亲本性状平均值; 纵轴线F2表示F2代性状平均值,下同。Note: P indicates the average value of character of the parents; F2 indicates the average value of character of F2 generation, The same as below. 图3 10 h短日照条件下F2代叶长频数分布Fig.3 Frequency distribution of leaf length of F2 generation in 10 h short day 2.4.3不同光周期条件下金花菜F2代叶宽的次数分布 10 h短日照条件下,金花菜 F2代叶宽( 2.74 cm)高于亲本P1(2.50 cm)而低于亲本P2(2.91 cm)。F2代叶宽集中在2.45 ~ 3.05 cm,出现83次,占总数的83%。10 h短日照条件下,金花菜F2代叶宽服从正态分布(表3),偏度 - 0.47 < 0,峰度1.31,其数据向右靠拢,但偏斜程度小,有较为明显的尖峰,数据集中(图5),表明10 h短日照条件下,金花菜F2代叶长较亲本P1有增加的趋势。16 h长日照条件下,F2代叶宽(2.40 cm)高于亲本P1(1.99 cm)而低于亲本P2(2.50 cm)。F2代叶宽集中在2.3~2.7 cm,出现93次,占总数的75%。16 h长日照条件下,金花菜F2代叶宽服从正态分布,偏度 - 0.11 < 0,峰度 - 0.24,其数据向右靠拢,但偏斜程度小(图6),表明16 h长日照条件下,金花菜F2代叶长较亲本P1有增加的趋势,且倾向亲本P2。 图4 16 h长日照条件下F2代叶长频数分布Fig.4 Frequency distribution of leaf length of F2 generation in 16 h long day 图5 10 h短日照条件下F2代叶宽频数分布Fig.5 Frequency distribution of leaf width of F2 generation in 10 h short day 2.4.4不同光周期条件下金花菜F2代分枝数的次数分布 10 h短日照条件下,金花菜F2代分枝数(1.76个)低于亲本P1(4.5个)而高于亲本P2(1.0个)。F2代分枝数集中在1个,出现68次,占总数的68 %。10 h短日照条件下,F2代分枝数不服从正态分布(表3),偏度1.53 > 0,峰度1.11,其数据向左集中靠拢(图7);16 h长日照条件下,金花菜F2代分枝数(1.5个),低于亲本P1(4.1个)而高于亲本P2(1.0个)。F2代分枝数集中在1个,出现89次,占总数的71.77%。16 h长日照条件下,F2代分枝数不服从正态分布,偏度1.95 > 0,峰度3.50 > 3,其数据向左集中靠拢,有明显尖峰(图8)。结果表明即使在不同光周期条件下,金花菜F2代分枝数的变异都倾向于分枝数为1的母本P2。 图6 16 h长日照条件下F2代叶宽频数分布Fig.6 Frequency distribution of leaf width of F2 generation in 16 h long day 图7 10 h短日照条件下F2代分枝数频数分布Fig.7 Frequency distribution of branches of F2 generation in 10 h short day 图8 16 h长日照条件下F2代分枝数频数分布Fig.8 Frequency distribution of branches of F2 generation in 16 h long day 种质资源的鉴定、分类和评价以及育种材料的选择通常都是依据植物农艺性状来进行的,在揭示植物进化的变异规律和建立植物演化模式等方面具有十分重要的作用[12]。由于地理分布和生境的差异,不同种质材料为了适应生态环境,相应地表现出了丰富的形态多样性。同时这也为育种者提供了种间差异大且遗传多样性丰富的材料,有利于优势育种材料的选择。本研究根据陈祥[11]的试验结论选择了其中形态及生理特性具有显著差异(P<0.05)的2个金花菜材料进行不同光周期条件下的开花诱导试验,并以此分析了不同光周期条件下金花菜F2代主要农艺性状变异特征。不同光周期条件下,金花菜F2代的株高、叶长和叶宽相对于亲本P1都有增加的趋势,金花菜F2代与来自江苏海安的金花菜种质材料相比有很明显的改良效果。金花菜F2代的分枝数和株高在不同光周期下变异系数都很大,且以分枝数为最,并且分枝数的变异都倾向于分枝数为1的母本P2,表明日照条件并不是影响分枝数变异的主要因素,种质特性才是影响金花菜F2代分枝数变异的关键所在。本试验中还观察到,在同一光周期条件下,若F2代分枝数较少,则植株较高,叶片较大,株型趋于母本P2;若其分枝数较多,则植株较矮,叶片较小,株型趋于父本P1。因此,分枝数是判断金花菜F2代在性状变异方面更偏向于母本还是父本的主要性状,而株高、叶长和叶宽则可作为判断的辅助性状。 自Garner和Allard[13]通过人工调控光周期证明大豆是短日照植物以来,利用光周期对水稻(OryzasativaL.)、玉米(ZeamaysL.)以及其他作物的种质资源的研究报道有很多。宋远丽等[14]的研究指出不同的植物对光周期的要求是不同的,例如,水稻是短日照植物而拟南芥(ArabidopsisthalialaL.)则是长日照植物。江院等[15]通过对光周期敏感的玉米材料进行了不同光周期处理,研究了光周期对玉米农艺性状和光合特性指标的影响,表明玉米属于短日照植物,长光周期可以促进玉米营养生长阶段的生长发育。本研究中,短日照条件下金花菜花期长,长日照条件下金花菜花期短,而且金花菜各世代在短日照条件下营养生长的性状指标要明显高于其在长日照条件下营养生长的性状指标,长光周期促进金花菜从营养生长阶段向生殖生长阶段的转变,使金花菜成花加快,生育期变短,表明金花菜属于长日照植物,与陈祥等[11]的研究结论一致。光周期的改变使得金花菜花期、性状和生育期产生了变化,却也遵循着一定的生长规律。因此,在栽培生产中,可以利用短日照条件延长金花菜生育期,提高性状指标,有效增加刈割茬数,获得高产。同时,也可以利用短日照条件筛选优良金花菜种质,培育优质高产品种。 不同光周期条件下,金花菜F2代株高、叶长和叶宽都服从正态分布,而分枝数不服从正态分布。不同光周期条件下,金花菜F2代株高、叶长和叶宽都具有极显著差异(P<0.01),而分枝数具有显著差异(P< 0.05)。无论在何种日照条件下,金花菜F2代分枝数的变异系数都远大于其株高、叶长和叶宽的变异系数,表明金花菜F2代分枝数的变异更为突出;即使在不同光周期条件下,金花菜F2代分枝数的变异都倾向于分枝数为1的母本P2。 本试验得出金花菜属于长日照植物。在栽培生产中,利用短日照诱导延长金花菜生育期,改善目标性状,增加刈割茬数,获得高产。在金花菜的品种选育中,利用此特性筛选优良金花菜种质,培育优质高产品种。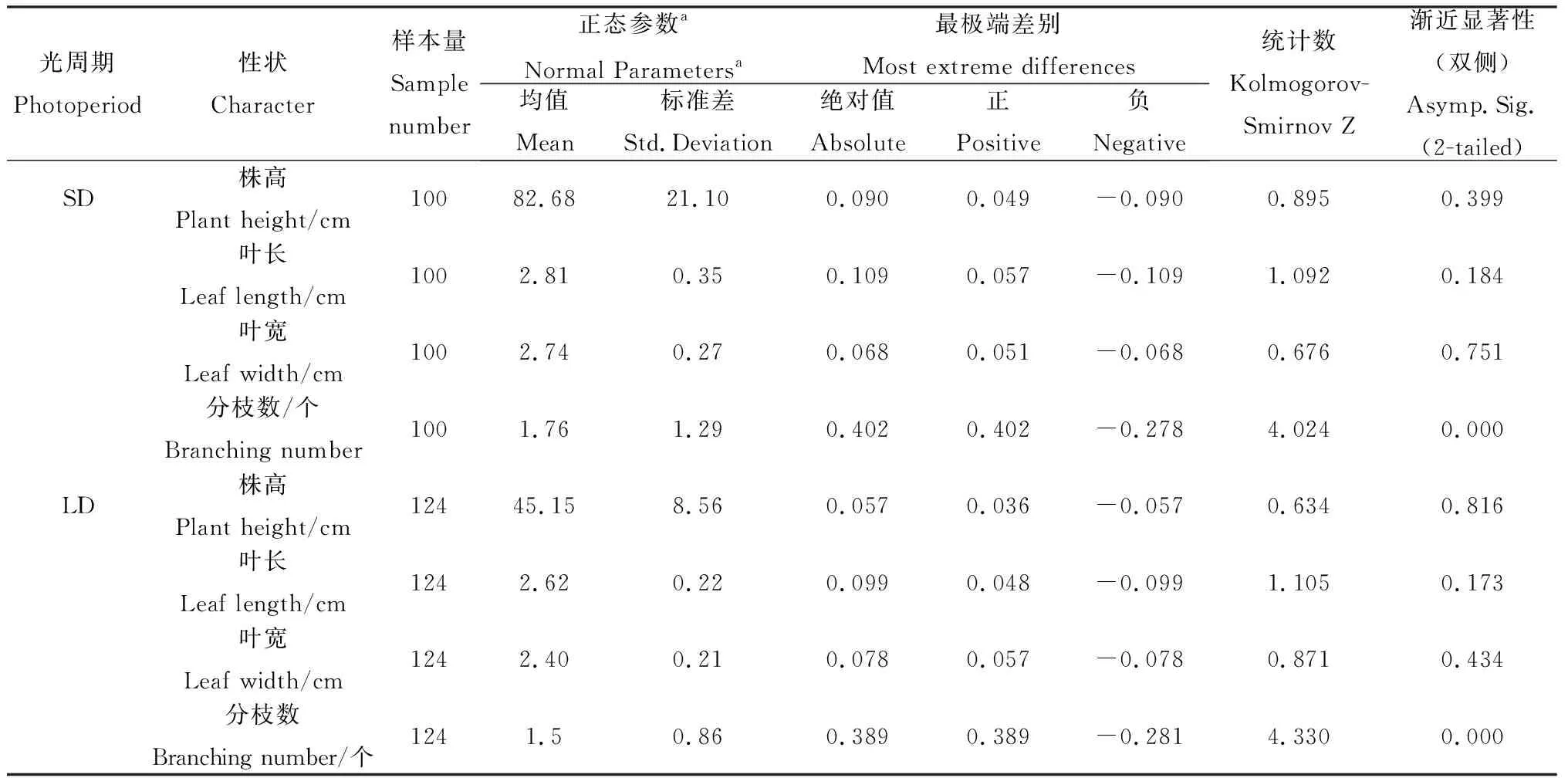

2.4 不同光周期条件下金花菜F2代农艺性状的次数分布




3 讨论
4 结论
猜你喜欢
植物保护(2024年4期)2024-01-01 00:00:00
农业科技通讯(2023年1期)2023-02-12 07:08:50
新疆农业科学(2020年1期)2020-02-14 03:22:46
亚热带植物科学(2019年2期)2019-07-22 13:16:12
江苏农业学报(2018年5期)2018-09-10 22:36:44
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
江苏农业科学(2016年10期)2017-02-05 14:45:09
家家乐·上半月(2013年6期)2013-04-29 00:44:03
